
First Report of Colistin-Resistant Escherichia coli Carrying mcr-1 IncI2(delta) and IncX4 Plasmids from Camels (Camelus dromedarius) in the Gulf Region
 , , , ,
, , , ,  , and
, and
Abstract
1. Introduction
2. Results
2.1. Detection of mcr Genes, Plasmid Profiling and Antibiotic Susceptibility Profile
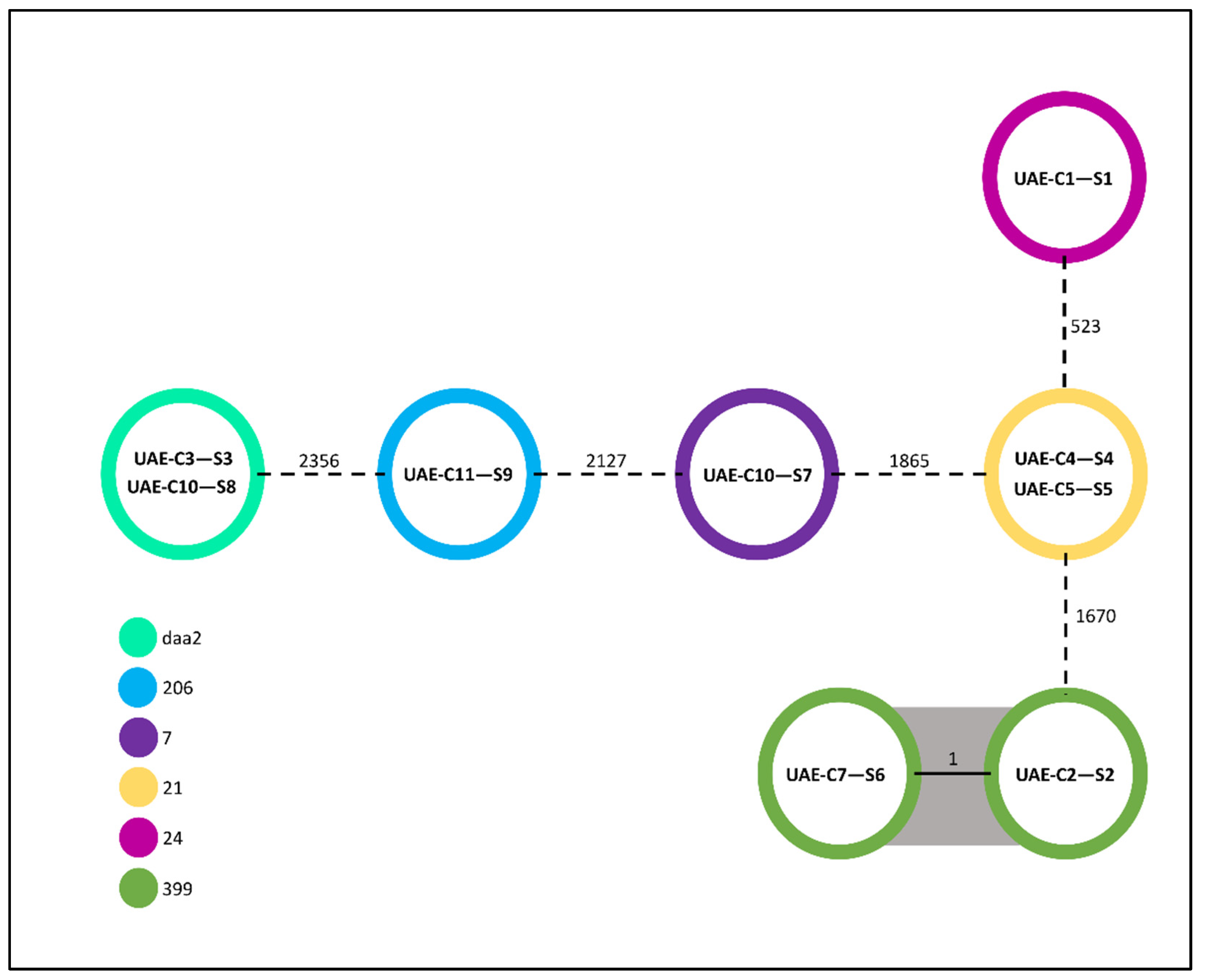
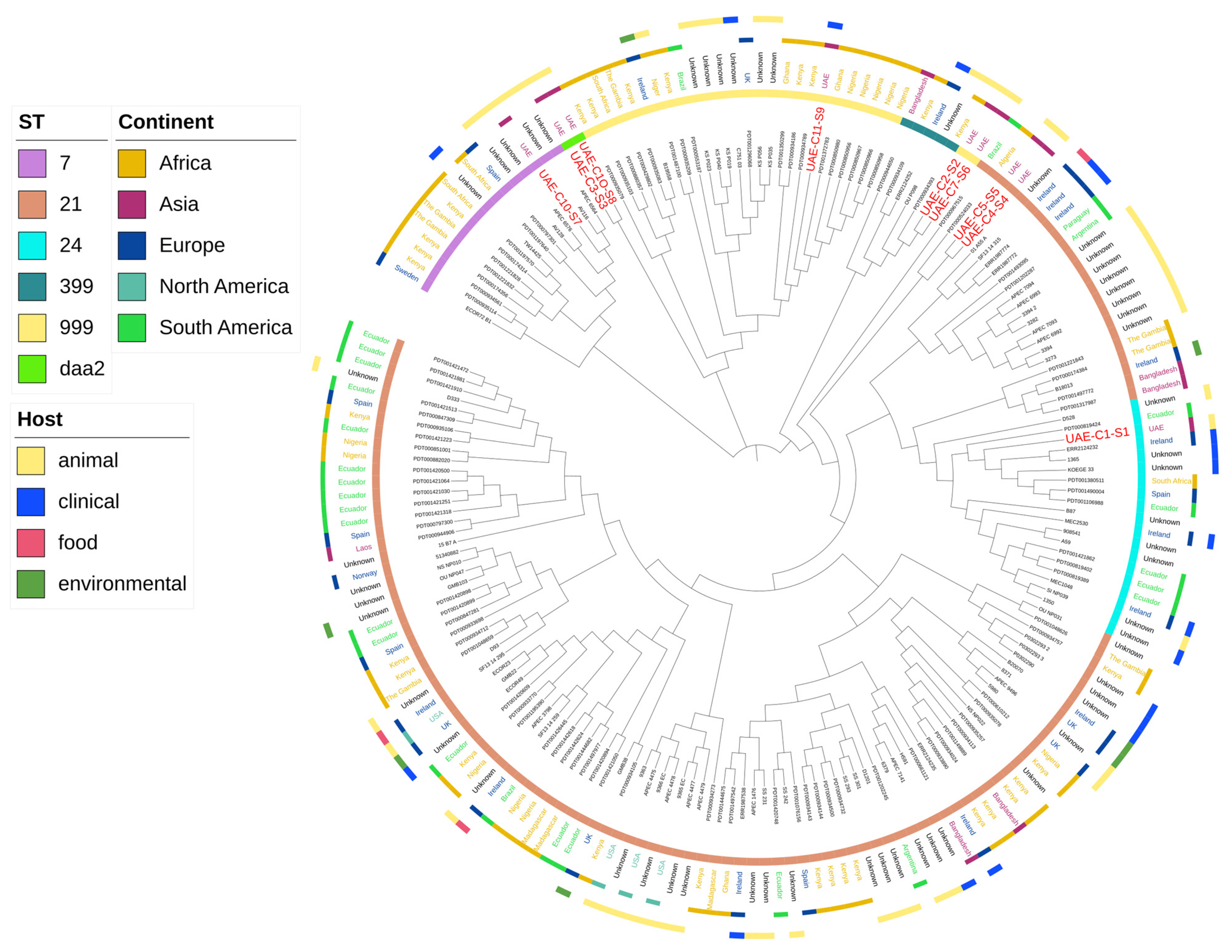
2.2. Clonality of the Isolates
2.3. Antimicrobial Resistance Genes
2.4. Serotype of E. coli Isolates
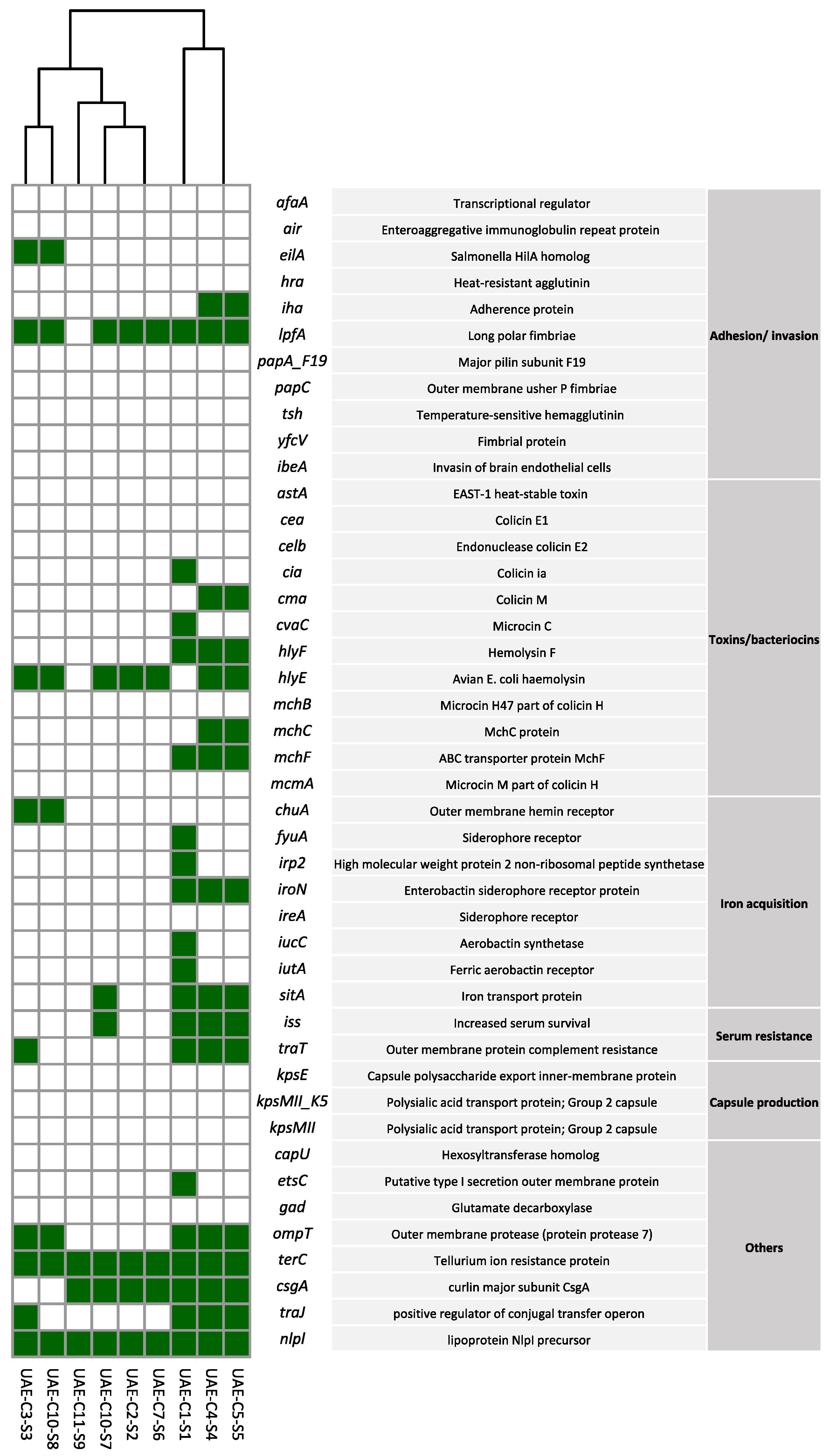
2.5. Virulence Profiles of mcr-1 Producing E. coli Isolates
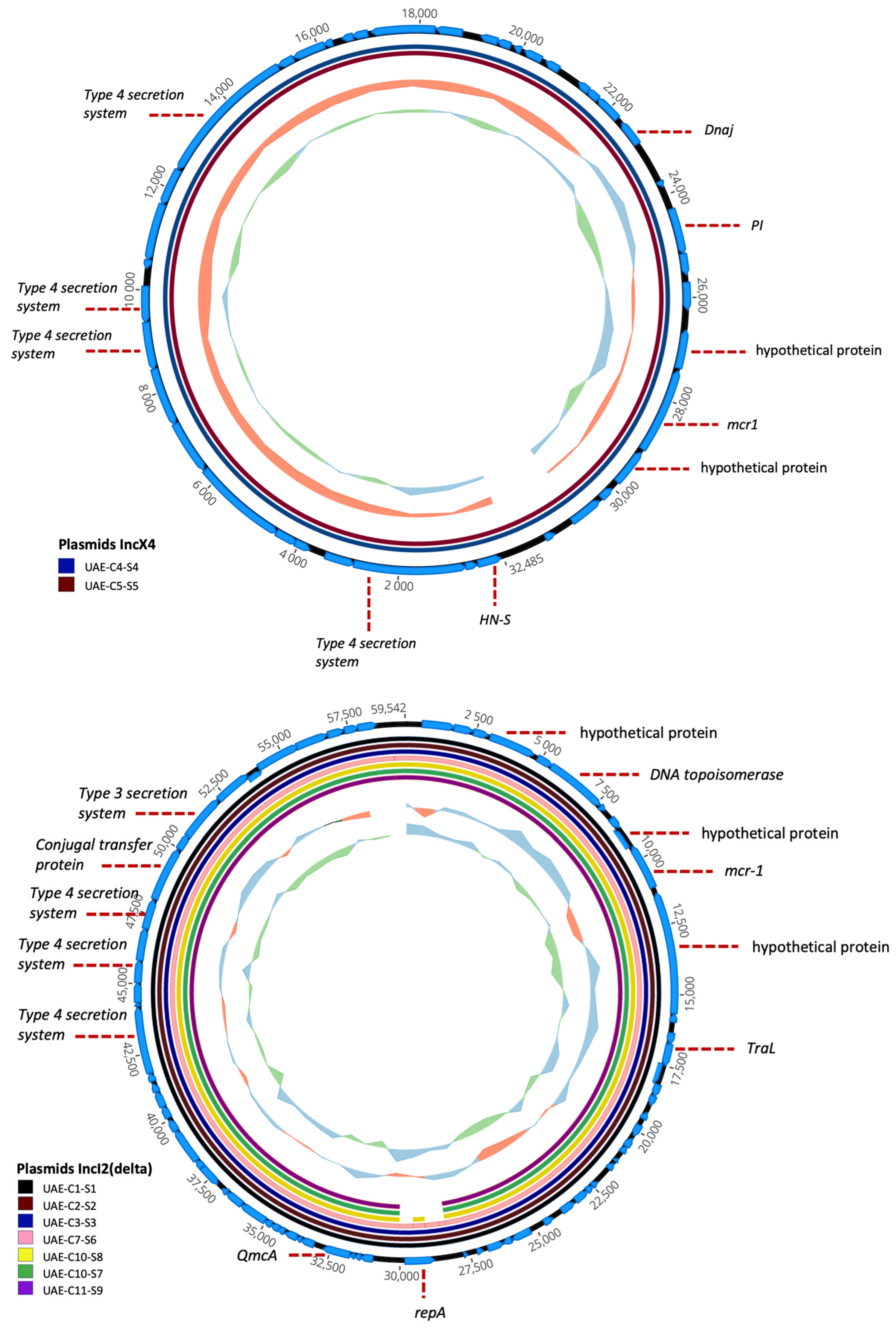
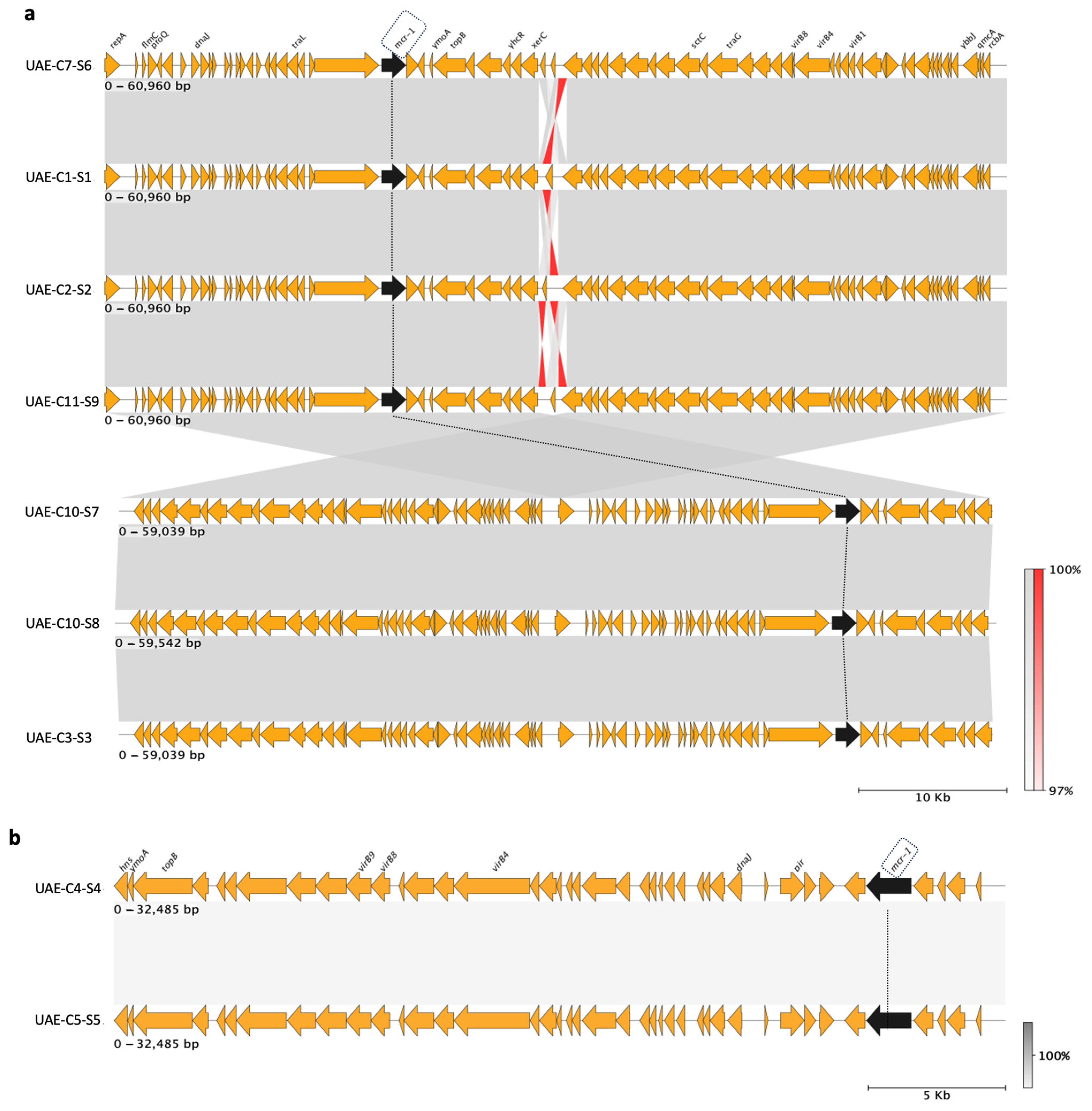
2.6. Characterization of mcr-1 Plasmids
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Detection of mcr Genes and Plasmid Profiling
4.3. Antimicrobial Susceptibility Test
4.4. Whole-Genome Sequencing
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lim, L.M.; Ly, N.; Anderson, D.; Yang, J.C.; Macander, L.; Jarkowski, A., III; Forrest, A.; Bulitta, J.B.; Tsuji, B.T. Resurgence of colistin: A review of resistance, toxicity, pharmacodynamics, and dosing. Pharmacotherapy 2010, 30, 1279–1291. [Google Scholar] [CrossRef]
- Bialvaei, A.Z.; Samadi Kafil, H. Colistin, mechanisms and prevalence of resistance. Curr. Med. Res. Opin. 2015, 31, 707–721. [Google Scholar] [CrossRef]
- Dhariwal, A.K.; Tullu, M.S. Colistin: Re-emergence of the ‘forgotten’ antimicrobial agent. J. Postgrad. Med. 2013, 59, 208–215. [Google Scholar] [CrossRef]
- Capone, A.; Giannella, M.; Fortini, D.; Giordano, A.; Meledandri, M.; Ballardini, M.; Venditti, M.; Bordi, E.; Capozzi, D.; Balice, M.P.; et al. High rate of colistin resistance among patients with carbapenem-resistant Klebsiella pneumoniae infection accounts for an excess of mortality. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2013, 19, E23–E30. [Google Scholar] [CrossRef]
- Nang, S.C.; Li, J.; Velkov, T. The rise and spread of mcr plasmid-mediated polymyxin resistance. Crit. Rev. Microbiol. 2019, 45, 131–161. [Google Scholar] [CrossRef]
- Caniaux, I.; van Belkum, A.; Zambardi, G.; Poirel, L.; Gros, M.F. MCR: Modern colistin resistance. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2017, 36, 415–420. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, H.; Liu, Y.H.; Feng, Y. Towards Understanding MCR-like Colistin Resistance. Trends Microbiol. 2018, 26, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Caselli, E.; D’Accolti, M.; Soffritti, I.; Piffanelli, M.; Mazzacane, S. Spread of mcr-1-Driven Colistin Resistance on Hospital Surfaces, Italy. Emerg. Infect. Dis. 2018, 24, 1752–1753. [Google Scholar] [CrossRef] [PubMed]
- Sia, C.M.; Greig, D.R.; Day, M.; Hartman, H.; Painset, A.; Doumith, M.; Meunier, D.; Jenkins, C.; Chattaway, M.A.; Hopkins, K.L.; et al. The characterization of mobile colistin resistance (mcr) genes among 33000 Salmonella enterica genomes from routine public health surveillance in England. Microb. Genom. 2020, 6, e000331. [Google Scholar] [CrossRef] [PubMed]
- García, V.; García-Meniño, I.; Mora, A.; Flament-Simon, S.C.; Díaz-Jiménez, D.; Blanco, J.E.; Alonso, M.P.; Blanco, J. Co-occurrence of mcr-1, mcr-4 and mcr-5 genes in multidrug-resistant ST10 Enterotoxigenic and Shiga toxin-producing Escherichia coli in Spain (2006-2017). Int. J. Antimicrob. Agents 2018, 52, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Hadjadj, L.; Baron, S.A.; Olaitan, A.O.; Morand, S.; Rolain, J.M. Co-occurrence of Variants of mcr-3 and mcr-8 Genes in a Klebsiella pneumoniae Isolate From Laos. Front. Microbiol. 2019, 10, 2720. [Google Scholar] [CrossRef]
- Callens, B.; Persoons, D.; Maes, D.; Laanen, M.; Postma, M.; Boyen, F.; Haesebrouck, F.; Butaye, P.; Catry, B.; Dewulf, J. Prophylactic and metaphylactic antimicrobial use in Belgian fattening pig herds. Prev. Vet. Med. 2012, 106, 53–62. [Google Scholar] [CrossRef]
- Zowawi, H.M.; Sartor, A.L.; Balkhy, H.H.; Walsh, T.R.; Al Johani, S.M.; AlJindan, R.Y.; Alfaresi, M.; Ibrahim, E.; Al-Jardani, A.; Al-Abri, S.; et al. Molecular characterization of carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in the countries of the Gulf cooperation council: Dominance of OXA-48 and NDM producers. Antimicrob. Agents Chemother. 2014, 58, 3085–3090. [Google Scholar] [CrossRef]
- Sonnevend, Á.; Ghazawi, A.A.; Hashmey, R.; Jamal, W.; Rotimi, V.O.; Shibl, A.M.; Al-Jardani, A.; Al-Abri, S.S.; Tariq, W.U.; Weber, S.; et al. Characterization of Carbapenem-Resistant Enterobacteriaceae with High Rate of Autochthonous Transmission in the Arabian Peninsula. PLoS ONE 2015, 10, e0131372. [Google Scholar] [CrossRef]
- Moubareck, C.A.; Mouftah, S.F.; Pál, T.; Ghazawi, A.; Halat, D.H.; Nabi, A.; AlSharhan, M.A.; AlDeesi, Z.O.; Peters, C.C.; Celiloglu, H.; et al. Clonal emergence of Klebsiella pneumoniae ST14 co-producing OXA-48-type and NDM carbapenemases with high rate of colistin resistance in Dubai, United Arab Emirates. Int. J. Antimicrob. Agents 2018, 52, 90–95. [Google Scholar] [CrossRef]
- Sonnevend, Á.; Ghazawi, A.; Darwish, D.; Barathan, G.; Hashmey, R.; Ashraf, T.; Rizvi, T.A.; Pál, T. In vitro efficacy of ceftazidime-avibactam, aztreonam-avibactam and other rescue antibiotics against carbapenem-resistant Enterobacterales from the Arabian Peninsula. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2020, 99, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Bessalah, S.; Salhi, I.; Thériault, W.; Fairbrother, J.M.; Fravalo, P. Screening for fecal presence of colistin-resistant Escherichia coli and mcr-1 and mcr-2 genes in camel-calves in southern Tunisia. Acta Vet. Scand. 2018, 60, 35. [Google Scholar] [CrossRef] [PubMed]
- Saidani, M.; Messadi, L.; Mefteh, J.; Chaouechi, A.; Soudani, A.; Selmi, R.; Dâaloul-Jedidi, M.; Ben Chehida, F.; Mamlouk, A.; Jemli, M.H.; et al. Various Inc-type plasmids and lineages of Escherichia coli and Klebsiella pneumoniae spreading blaCTX-M-15, blaCTX-M-1 and mcr-1 genes in camels in Tunisia. J. Glob. Antimicrob. Resist. 2019, 19, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Nüesch-Inderbinen, M.; Kindle, P.; Baschera, M.; Liljander, A.; Jores, J.; Corman, V.M.; Stephan, R. Antimicrobial resistant and extended-spectrum ß-lactamase (ESBL) producing Escherichia coli isolated from fecal samples of African dromedary camels. Sci. Afr. 2020, 7, e00274. [Google Scholar] [CrossRef]
- Ngbede, E.O.; Poudel, A.; Kalalah, A.; Yang, Y.; Adekanmbi, F.; Adikwu, A.A.; Adamu, A.M.; Mamfe, L.M.; Daniel, S.T.; Useh, N.M.; et al. Identification of mobile colistin resistance genes (mcr-1.1, mcr-5 and mcr-8.1) in Enterobacteriaceae and Alcaligenes faecalis of human and animal origin, Nigeria. Int. J. Antimicrob. Agents 2020, 56, 106108. [Google Scholar] [CrossRef] [PubMed]
- Alhababi, D.A.; Eltai, N.O.; Nasrallah, G.K.; Farg, E.A.; Al Thani, A.A.; Yassine, H.M. Antimicrobial Resistance of Commensal Escherichia coli Isolated from Food Animals in Qatar. Microb. Drug Resist. 2020, 26, 420–427. [Google Scholar] [CrossRef]
- Wani, N.A. In vitro embryo production (IVEP) in camelids: Present status and future perspectives. Reprod. Biol. 2021, 21, 100471. [Google Scholar] [CrossRef]
- Olaitan, A.O.; Morand, S.; Rolain, J.M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Biswas, U.; Das, S.; Barik, M.; Mallick, A. Situation Report on mcr-Carrying Colistin-Resistant Clones of Enterobacterales: A Global Update Through Human-Animal-Environment Interfaces. Curr. Microbiol. 2023, 81, 12. [Google Scholar] [CrossRef]
- Kempf, I.; Jouy, E.; Chauvin, C. Colistin use and colistin resistance in bacteria from animals. Int. J. Antimicrob. Agents 2016, 48, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Sonnevend, Á.; Alali, W.Q.; Mahmoud, S.A.; Ghazawi, A.; Bharathan, G.; Melegh, S.; Rizvi, T.A.; Pál, T. Molecular Characterization of MCR-1 Producing Enterobacterales Isolated in Poultry Farms in the United Arab Emirates. Antibiotics 2022, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Habib, I.; Elbediwi, M.; Ghazawi, A.; Mohamed, M.I.; Lakshmi, G.B.; Khan, M. First report from supermarket chicken meat and genomic characterization of colistin resistance mediated by mcr-1.1 in ESBL-producing, multidrug-resistant Salmonella Minnesota. Int. J. Food Microbiol. 2022, 379, 109835. [Google Scholar] [CrossRef] [PubMed]
- Habib, I.; Elbediwi, M.; Mohamed, M.I.; Ghazawi, A.; Abdalla, A.; Khalifa, H.O.; Khan, M. Enumeration, antimicrobial resistance and genomic characterization of extended-spectrum β-lactamases producing Escherichia coli from supermarket chicken meat in the United Arab Emirates. Int. J. Food Microbiol. 2023, 398, 110224. [Google Scholar] [CrossRef] [PubMed]
- Hussein, N.H.; Al-Kadmy, I.M.S.; Taha, B.M.; Hussein, J.D. Mobilized colistin resistance (mcr) genes from 1 to 10: A comprehensive review. Mol. Biol. Rep. 2021, 48, 2897–2907. [Google Scholar] [CrossRef]
- Madec, J.Y.; Haenni, M.; Nordmann, P.; Poirel, L. Extended-spectrum β-lactamase/AmpC- and carbapenemase-producing Enterobacteriaceae in animals: A threat for humans? Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2017, 23, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Yi, L.X.; Yu, L.F.; Wang, J.; Liu, Y.; Chen, X.; Lv, L.; Yang, J.; Liu, J.H. Fitness Advantage of mcr-1-Bearing IncI2 and IncX4 Plasmids in Vitro. Front. Microbiol. 2018, 9, 331. [Google Scholar] [CrossRef]
- Shen, C.; Zhong, L.L.; Yang, Y.; Doi, Y.; Paterson, D.L.; Stoesser, N.; Ma, F.; El-Sayed Ahmed, M.A.E.; Feng, S.; Huang, S.; et al. Dynamics of mcr-1 prevalence and mcr-1-positive Escherichia coli after the cessation of colistin use as a feed additive for animals in China: A prospective cross-sectional and whole genome sequencing-based molecular epidemiological study. Lancet. Microbe 2020, 1, e34–e43. [Google Scholar] [CrossRef] [PubMed]
- Paveenkittiporn, W.; Kamjumphol, W.; Ungcharoen, R.; Kerdsin, A. Whole-Genome Sequencing of Clinically Isolated Carbapenem-Resistant Enterobacterales Harboring mcr Genes in Thailand, 2016–2019. Front. Microbiol. 2021, 11, 586368. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wu, H.; Zhuang, Y.; Luo, J.; Chen, Y.; Wu, Y.; Fei, J.; Shen, Q.; Yuan, Z.; Chen, M. Stability and genetic insights of the co-existence of blaCTX-M-65, blaOXA-1, and mcr-1.1 harboring conjugative IncI2 plasmid isolated from a clinical extensively-drug resistant Escherichia coli ST744 in Shanghai. Front. Public Health 2023, 11, 1216704. [Google Scholar] [CrossRef] [PubMed]
- Carhuaricra, D.; Duran Gonzales, C.G.; Rodríguez Cueva, C.L.; Ignacion León, Y.; Silvestre Espejo, T.; Marcelo Monge, G.; Rosadio Alcántara, R.H.; Lincopan, N.; Espinoza, L.L.; Maturrano Hernández, L. Occurrence and Genomic Characterization of mcr-1-Harboring Escherichia coli Isolates from Chicken and Pig Farms in Lima, Peru. Antibiotics 2022, 11, 1781. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Du, P.; Zhang, P.; Li, Y.; Yang, X.; Wang, Z.; Wang, J.; Bai, L. Comprehensive Genomic Investigation of Coevolution of mcr genes in Escherichia coli Strains via Nanopore Sequencing. Glob. Chall. 2021, 5, 2000014. [Google Scholar] [CrossRef]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef]
- Li, W.; Liu, Z.; Yin, W.; Yang, L.; Qiao, L.; Song, S.; Ling, Z.; Zheng, R.; Wu, C.; Wang, Y.; et al. MCR Expression Conferring Varied Fitness Costs on Host Bacteria and Affecting Bacteria Virulence. Antibiotics 2021, 10, 872. [Google Scholar] [CrossRef]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. 2018, 23, 17–00672. [Google Scholar] [CrossRef]
- Borowiak, M.; Baumann, B.; Fischer, J.; Thomas, K.; Deneke, C.; Hammerl, J.A.; Szabo, I.; Malorny, B. Development of a Novel mcr-6 to mcr-9 Multiplex PCR and Assessment of mcr-1 to mcr-9 Occurrence in Colistin-Resistant Salmonella enterica Isolates From Environment, Feed, Animals and Food (2011–2018) in Germany. Front. Microbiol. 2020, 11, 80. [Google Scholar] [CrossRef]
- Kado, C.I.; Liu, S.T. Rapid procedure for detection and isolation of large and small plasmids. J. Bacteriol. 1981, 145, 1365–1373. [Google Scholar] [CrossRef]
- CLSI Performance Standards for Antimicrobial Susceptibility Testing, 33rd ed.; CLSI Guideline M100Performance Standards for Antimicrobial Susceptibility Testing; CLSI: Wayne, PA, USA, 2022.
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Mikheenko, A.; Prjibelski, A.; Saveliev, V.; Antipov, D.; Gurevich, A. Versatile genome assembly evaluation with QUAST-LG. Bioinformatics 2018, 34, i142–i150. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Néron, B.; Littner, E.; Haudiquet, M.; Perrin, A.; Cury, J.; Rocha, E.P.C. IntegronFinder 2.0: Identification and Analysis of Integrons across Bacteria, with a Focus on Antibiotic Resistance in Klebsiella. Microorganisms 2022, 10, 700. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Maiden, M.C. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinform. 2010, 11, 595. [Google Scholar] [CrossRef]
- Gardner, S.N.; Slezak, T.; Hall, B.G. kSNP3.0: SNP detection and phylogenetic analysis of genomes without genome alignment or reference genome. Bioinformatics 2015, 31, 2877–2878. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Camel ID | Colistin MIC (mg/L) | Resistance Detected by Disc Diffusion | |
|---|---|---|---|---|
| Resistant | Intermediate | |||
| UAE-C1-S1 | 1-inside | 4 | AUG, TET, SXT, AP, DXT | CAZ, CPD, NA |
| UAE-C2-S2 | 2-inside | 4 | NA | CIP, AP |
| UAE-C3-S3 | 3-inside | 4 | - | CIP, NA |
| UAE-C4-S4 | 4-outside | 4 | GM, TET, CIP, SXT, CHL, NA, AP, DXT, TOB | AUG |
| UAE-C5-S5 | 5-outside | 4 | GM, TET, CIP, SXT, CHL, NA, AP, DXT, TOB | AUG |
| UAE-C7-S6 | 7-outside | 4 | NA | CIP, AP |
| UAE-C10-S7 | 10-outside | 4 | - | NA |
| UAE-C10-S8 | 10-outside | 4 | - | NA |
| UAE-C11-S9 | 11-outside | 4 | NA | CIP, AP |
| Strains | Sequence Type | Phylogroup | Serotype | FimH Type | CH Type: 4-31 |
|---|---|---|---|---|---|
| UAE-C1-S1 | 24 | B1 | H25, O9 | fimH27 | fumC4-fimH24: 4-27 |
| UAE-C2-S2 | 399 | B1 | H14, O81 | fimH24 | fumC6-fimH24: 6-24 |
| UAE-C3-S3 | daa2 | D | H48, O56 | fimH577 | fumC26-fimH577: 26-577 |
| UAE-C4-S4 | 21 | B1 | H9, O49 | fimH32 | fumC4-fimH32: 4-32 |
| UAE-C5-S5 | 21 | B1 | H9, O49 | fimH32 | fumC4-fimH32: 4-32 |
| UAE-C7-S6 | 399 | B1 | H14, O81 | fimH24 | fumC6-fimH24: 6-24 |
| UAE-C10-S7 | 7 | C | H9, O21 | fimH35 | fumC4-fimH35: 4-35 |
| UAE-C10-S8 | daa2 | D | H48, O56 | fimH577 | fumC26-fimH577: 26-577 |
| UAE-C11-S9 | 999 | A | H5 | fimH41 | fumC7-fimH41: 7-41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghazawi, A.; Strepis, N.; Anes, F.; Yaaqeib, D.; Ahmed, A.; AlHosani, A.; AlShehhi, M.; Manzoor, A.; Habib, I.; Wani, N.A.; et al. First Report of Colistin-Resistant Escherichia coli Carrying mcr-1 IncI2(delta) and IncX4 Plasmids from Camels (Camelus dromedarius) in the Gulf Region. Antibiotics 2024, 13, 227. https://doi.org/10.3390/antibiotics13030227
Ghazawi A, Strepis N, Anes F, Yaaqeib D, Ahmed A, AlHosani A, AlShehhi M, Manzoor A, Habib I, Wani NA, et al. First Report of Colistin-Resistant Escherichia coli Carrying mcr-1 IncI2(delta) and IncX4 Plasmids from Camels (Camelus dromedarius) in the Gulf Region. Antibiotics. 2024; 13(3):227. https://doi.org/10.3390/antibiotics13030227
Chicago/Turabian StyleGhazawi, Akela, Nikolaos Strepis, Febin Anes, Dana Yaaqeib, Amal Ahmed, Aysha AlHosani, Mirah AlShehhi, Ashrat Manzoor, Ihab Habib, Nisar A. Wani, and et al. 2024. "First Report of Colistin-Resistant Escherichia coli Carrying mcr-1 IncI2(delta) and IncX4 Plasmids from Camels (Camelus dromedarius) in the Gulf Region" Antibiotics 13, no. 3: 227. https://doi.org/10.3390/antibiotics13030227
APA StyleGhazawi, A., Strepis, N., Anes, F., Yaaqeib, D., Ahmed, A., AlHosani, A., AlShehhi, M., Manzoor, A., Habib, I., Wani, N. A., Hays, J. P., & Khan, M. (2024). First Report of Colistin-Resistant Escherichia coli Carrying mcr-1 IncI2(delta) and IncX4 Plasmids from Camels (Camelus dromedarius) in the Gulf Region. Antibiotics, 13(3), 227. https://doi.org/10.3390/antibiotics13030227