
Lack of Association between Antimicrobial Consumption and Antimicrobial Resistance in a HIV Preexposure Prophylaxis Population: A Cross-Sectional Study

 , and
, and
Abstract
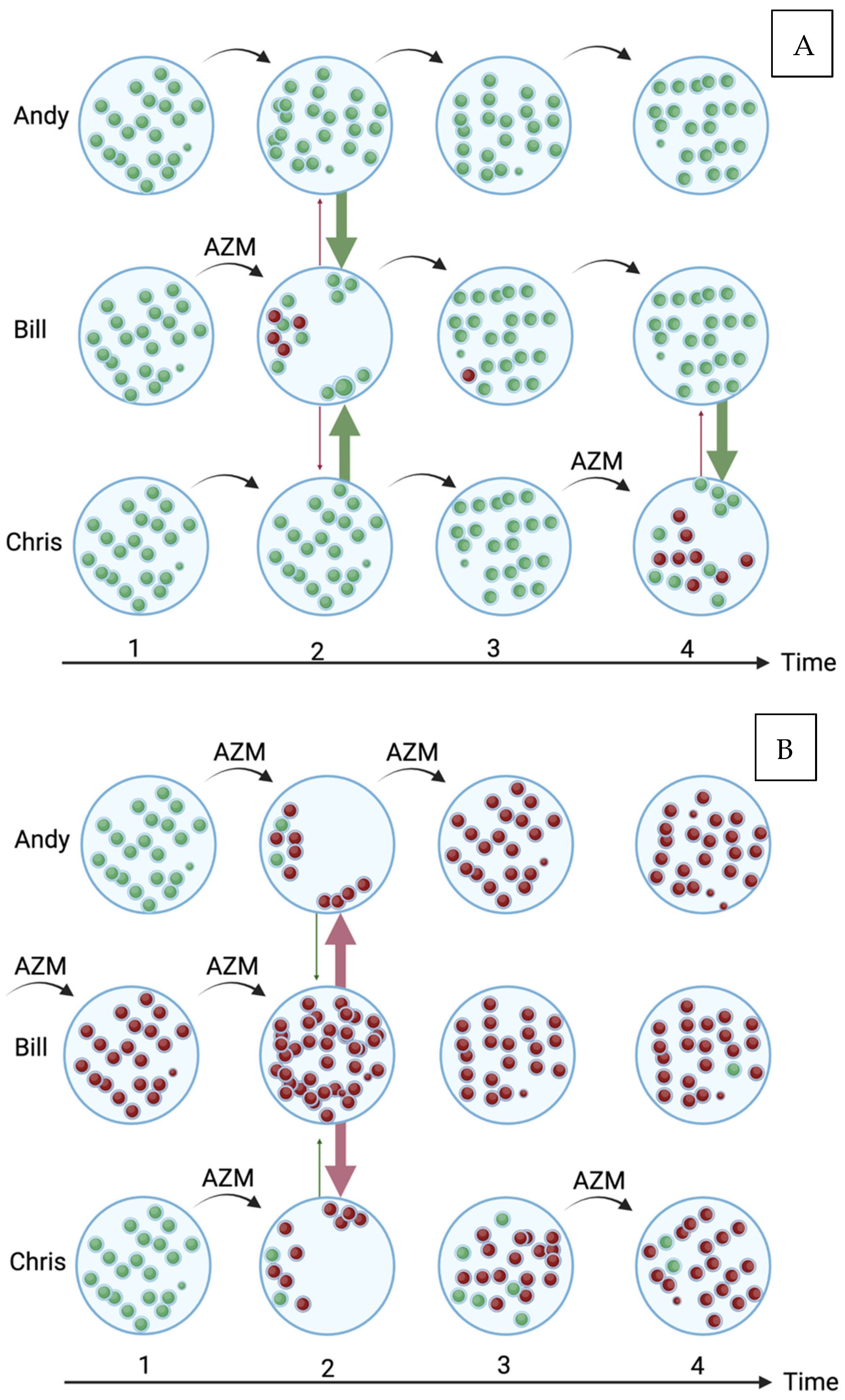
1. Introduction
2. Methods
2.1. Study Population
2.2. Data and Samples
2.3. DNA Extraction, Shotgun Metagenomic Sequencing and Resistome Characterisation
2.4. Taxonomic and Resistome Characterisation
2.5. Culture and Antimicrobial Susceptibility Determination of Neisseria Species
2.6. Ethics
3. Data Analysis
3.1. Antimicrobial Resistance Gene (ARG) Abundance
3.2. Antimicrobial Susceptibility of N. subflava
4. Results
4.1. Antimicrobial Susceptibility of N. subflava
4.2. Abundance of Resistance Associated Genes
4.3. Association AMU/Susceptibility in N. subflava
4.4. Association AMU/Gene Abundance
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bahn, J.M.; Ackerman, H.; Carpenter, C.M. Development in vitro of Penicillin-Resistant Strains of the Gonococcus. Proc. Soc. Exp. Biol. Med. 1945, 58, 21–24. [Google Scholar] [CrossRef]
- Carpenter, C.M.; Bahn, J.M.; Bahn, J.M.; Stokinger, H.E. Adaptability of gonococcus to four bacteriostatic agents, sodium sulfathiazole, rivanol lactate, promin, and penicillin. Proc. Soc. Exp. Biol. Med. 1945, 60, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Bell, B.G.; Schellevis, F.; Stobberingh, E.; Goossens, H.; Pringle, M. A systematic review and meta-analysis of the effects of antibiotic consumption on antibiotic resistance. BMC Infect. Dis. 2014, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.C.; Nogueira, P.J.; Paiva, J.-A. Determinants of antimicrobial resistance among the different European countries: More than human and animal antimicrobial consumption. Antibiotics 2021, 10, 834. [Google Scholar] [CrossRef]
- Bungau, S.; Tit, D.M.; Behl, T.; Aleya, L.; Zaha, D.C. Aspects of excessive antibiotic consumption and environmental influences correlated with the occurrence of resistance to antimicrobial agents. Curr. Opin. Environ. Sci. Health 2021, 19, 100224. [Google Scholar] [CrossRef]
- Cižman, M.; Srovin, T.P. Antibiotic consumption and resistance of gram-negative pathogens (collateral damage). GMS Infect. Dis. 2018, 6, Doc05. [Google Scholar]
- Baditoiu, L.; Axente, C.; Lungeanu, D.; Muntean, D.; Horhat, F.; Moldovan, R.; Hogea, E.; Bedreag, O.; Sandesc, D.; Licker, M. Intensive care antibiotic consumption and resistance patterns: A cross-correlation analysis. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 71. [Google Scholar] [CrossRef]
- Lai, C.-C.; Wang, C.-Y.; Chu, C.-C.; Tan, C.-K.; Lu, C.-L.; Lee, Y.-C.; Huang, Y.-T.; Lee, P.-I.; Hsueh, P.-R. Correlation between antibiotic consumption and resistance of Gram-negative bacteria causing healthcare-associated infections at a university hospital in Taiwan from 2000 to 2009. J. Antimicrob. Chemother. 2011, 66, 1374–1382. [Google Scholar] [CrossRef]
- Kenyon, C.; Manoharan-Basil, S.S.; Van Dijck, C. Is there a resistance threshold for macrolide consumption? Positive evidence from an ecological analysis of resistance data from streptococcus pneumoniae, treponema pallidum, and mycoplasma genitalium. Microb. Drug Resist. 2021, 27, 1079–1086. [Google Scholar] [CrossRef]
- Baquero, F.; Baquero-Artigao, G.; Cantón, R.; García-Rey, C. Antibiotic consumption and resistance selection in Streptococcus pneumoniae. J. Antimicrob. Chemother. 2002, 50, 27–38. [Google Scholar] [CrossRef]
- Bruyndonckx, R.; Hens, N.; Aerts, M.; Goossens, H.; Abrahantes, J.C.; Coenen, S. Exploring the association between resistance and outpatient antibiotic use expressed as DDDs or packages. J. Antimicrob. Chemother. 2015, 70, 1241–1244. [Google Scholar] [CrossRef]
- Goossens, H.; Ferech, M.; Stichele, R.V.; Elseviers, M. Outpatient antibiotic use in Europe and association with resistance: A cross-national database study. Lancet 2005, 365, 579–587. [Google Scholar] [CrossRef]
- Machalek, D.A.; Tao, Y.; Shilling, H.; Jensen, J.S.; Unemo, M.; Murray, G.; Chow, E.P.F.; Low, N.; Garland, S.M.; Vodstrcil, L.A.; et al. Prevalence of mutations associated with resistance to macrolides and fluoroquinolones in Mycoplasma genitalium: A systematic review and meta-analysis. Lancet Infect. Dis. 2020, 20, 1302–1314. [Google Scholar] [CrossRef]
- Seppala, H. The effect of changes in the consumption of macrolide antibiotics on erythromycin resistance in group A streptococci in Finland: Finnish Study Group for Antimicrobial Resistance. N. Engl. J. Med. 1997, 337, 441–446. [Google Scholar] [CrossRef]
- Bergman, M.; Huikko, S.; Huovinen, P.; Paakkari, P.; Seppala, H.; Finnish Study Group for Antimicrobial Resistance. Macrolide and azithromycin use are linked to increased macrolide resistance in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2006, 50, 3646–3650. [Google Scholar] [CrossRef] [PubMed]
- Čižman, M.; Pokorn, M.; Seme, K.; Oražem, A.; Paragi, M. The relationship between trends in macrolide use and resistance to macrolides of common respiratory pathogens. J. Antimicrob. Chemother. 2001, 47, 475–477. [Google Scholar] [CrossRef]
- Molina, J.-M.; Charreau, I.; Chidiac, C.; Pialoux, G.; Cua, E.; Delaugerre, C.; Capitant, C.; Rojas-Castro, D.; Fonsart, J.; Bercot, B. Post-exposure prophylaxis with doxycycline to prevent sexually transmitted infections in men who have sex with men: An open-label randomised substudy of the ANRS IPERGAY trial. Lancet Infect. Dis. 2018, 18, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Luetkemeyer, A.F.; Donnell, D.; Dombrowski, J.C.; Cohen, S.; Grabow, C.; Brown, C.E.; Malinski, C.; Perkins, R.; Nasser, M.; Lopez, C. Postexposure doxycycline to prevent bacterial sexually transmitted infections. N. Engl. J. Med. 2023, 388, 1296–1306. [Google Scholar] [CrossRef] [PubMed]
- Vanbaelen, T.; Van Dijck, C.; De Baetselier, I.; Florence, E.; Reyniers, T.; Vuylsteke, B.; Jacobs, B.K.; Kenyon, C. Screening for STIs is one of the main drivers of macrolide consumption in PrEP users. Int. J. STD AIDS 2021, 32, 1183–1184. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C. Dual azithromycin/ceftriaxone therapy for gonorrhoea in PrEP cohorts results in levels of macrolide consumption that exceed resistance thresholds by up to 7-fold. J. Infect. Dis. 2021, 224, 623–1624. [Google Scholar] [CrossRef] [PubMed]
- Laumen, J.G.E.; Manoharan-Basil, S.S.; Abdellati, S.; De Baetselier, I.; Van Dijck, C.; Martiny, D.; Serrano, G.; Bottieau, E.; Kenyon, C. Antimicrobial susceptibility of commensal Neisseria in the general population and men who have sex with men in Belgium. Sci. Rep. 2022, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Van Dijck, C.; Laumen, J.G.E.; de Block, T.; Abdellati, S.; De Baetselier, I.; Tsoumanis, A.; Malhotra-Kumar, S.; Manoharan-Basil, S.S.; Kenyon, C.; Xavier, B.B. The oropharynx of men using HIV pre-exposure prophylaxis is enriched with antibiotic resistance genes: A cross-sectional observational metagenomic study. J. Infect. 2023, 86, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Goytia, M.; Wadsworth, C.B. Canary in the Coal Mine: How Resistance Surveillance in Commensals Could Help Curb the Spread of AMR in Pathogenic Neisseria. Mbio 2022, 13, e01991-22. [Google Scholar] [CrossRef] [PubMed]
- Vanbaelen, T.; Van Dijck, C.; Laumen, J.; Gonzalez, N.; De Baetselier, I.; Manoharan-Basil, S.S.; De Block, T.; Kenyon, C. Global epidemiology of antimicrobial resistance in commensal Neisseria species: A systematic review. Int. J. Med. Microbiol. 2022, 312, 151551. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C.; Laumen, J.; Manoharan-Basil, S. Choosing new therapies for gonorrhoea: We need to consider the impact on the Pan-Neisseria Genome. A viewpoint. Antibiotics 2021, 10, 515. [Google Scholar] [CrossRef] [PubMed]
- Yahara, K.; Ma, K.C.; Mortimer, T.D.; Shimuta, K.; Nakayama, S.-i.; Hirabayashi, A.; Suzuki, M.; Jinnai, M.; Ohya, H.; Kuroki, T. Emergence and evolution of antimicrobial resistance genes and mutations in Neisseria gonorrhoeae. Genome Med. 2021, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lipsitch, M.; Samore, M.H. Antimicrobial use and antimicrobial resistance: A population perspective. Emerg. Infect. Dis. 2002, 8, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Van Dijck, C.; Tsoumanis, A.; Rotsaert, A.; Vuylsteke, B.; Van den Bossche, D.; Paeleman, E.; De Baetselier, I.; Brosius, I.; Laumen, J.; Buyze, J. Antibacterial mouthwash to prevent sexually transmitted infections in men who have sex with men taking HIV pre-exposure prophylaxis (PReGo): A randomised, placebo-controlled, crossover trial. Lancet Infect. Dis. 2021, 21, 657–667. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Doster, E.; Lakin, S.M.; Dean, C.J.; Wolfe, C.; Young, J.G.; Boucher, C.; Belk, K.E.; Noyes, N.R.; Morley, P.S. MEGARes 2.0: A database for classification of antimicrobial drug, biocide and metal resistance determinants in metagenomic sequence data. Nucleic Acids Res. 2020, 48, D561–D569. [Google Scholar] [CrossRef]
- Lu, J.; Breitwieser, F.P.; Thielen, P.; Salzberg, S.L. Bracken: Estimating species abundance in metagenomics data. PeerJ Comput. Sci. 2017, 3, e104. [Google Scholar] [CrossRef]
- Slizovskiy, I.B.; Mukherjee, K.; Dean, C.J.; Boucher, C.; Noyes, N.R. Mobilization of antibiotic resistance: Are current approaches for colocalizing resistomes and mobilomes useful? Front. Microbiol. 2020, 11, 1376. [Google Scholar] [CrossRef]
- Bennett, J.S.; Jolley, K.A.; Earle, S.G.; Corton, C.; Bentley, S.D.; Parkhill, J.; Maiden, M.C. A genomic approach to bacterial taxonomy: An examination and proposed reclassification of species within the genus Neisseria. Microbiology 2012, 158, 1570. [Google Scholar] [CrossRef] [PubMed]
- Malhotra-Kumar, S.; Lammens, C.; Coenen, S.; Van Herck, K.; Goossens, H. Effect of azithromycin and clarithromycin therapy on pharyngeal carriage of macrolide-resistant streptococci in healthy volunteers: A randomised, double-blind, placebo-controlled study. Lancet 2007, 369, 482–490. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Jernberg, C.; Andersson, A.F.; Sjolund-Karlsson, M.; Jansson, J.K.; Engstrand, L. Short-term antibiotic treatment has differing long-term impacts on the human throat and gut microbiome. PLoS ONE 2010, 5, e9836. [Google Scholar] [CrossRef] [PubMed]
- Choo, J.M.; Abell, G.C.; Thomson, R.; Morgan, L.; Waterer, G.; Gordon, D.L.; Taylor, S.L.; Leong, L.E.; Wesselingh, S.L.; Burr, L.D. Impact of long-term erythromycin therapy on the oropharyngeal microbiome and resistance gene reservoir in non-cystic fibrosis bronchiectasis. Msphere 2018, 3, e00103–e00118. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Stobberingh, E.E.; Savelkoul, P.H.; Wolffs, P.F. The human microbiome as a reservoir of antimicrobial resistance. Front. Microbiol. 2013, 4, 87. [Google Scholar] [CrossRef]
- Vanbaelen, T.; Florence, E.; Van Dijck, C.; Tsoumanis, A.; Laumen, J.G.E.; Manoharan-Basil, S.S.; Abdellati, S.; de Block, T.; De Baetselier, I.; Van den Bossche, D.; et al. Effect on the resistome of dual- vs monotherapy for the treatment of Neisseria gonorrhoeae: Results from a randomized controlled trial (ResistAZM Trial). Open Forum Infect. Dis. 2023, 10, ofad462. [Google Scholar] [CrossRef]
- Aversa, Z.; Atkinson, E.J.; Schafer, M.J.; Theiler, R.N.; Rocca, W.A.; Blaser, M.J.; LeBrasseur, N.K. Association of Infant Antibiotic Exposure With Childhood Health Outcomes. Mayo Clin. Proc. 2021, 96, 66–77. [Google Scholar] [CrossRef]
- Baquero, F.; Tedim, A.P.; Coque, T.M. Antibiotic resistance shaping multi-level population biology of bacteria. Front. Microbiol. 2013, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Kwong, J.C.; Chow, E.P.; Stevens, K.; Stinear, T.P.; Seemann, T.; Fairley, C.K.; Chen, M.Y.; Howden, B.P. Whole-genome sequencing reveals transmission of gonococcal antibiotic resistance among men who have sex with men: An observational study. Sex. Transm. Infect. 2018, 94, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rey, C.; Aguilar, L.; Baquero, F.; Casal, J.; Dal-Re, R. Importance of local variations in antibiotic consumption and geographical differences of erythromycin and penicillin resistance in Streptococcus pneumoniae. J. Clin. Microbiol. 2002, 40, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Mårdh, O.; Plachouras, D. Using doxycycline for prophylaxis of bacterial sexually transmitted infections: Considerations for the European Union and European Economic Area. Eurosurveillance 2023, 28, 2300621. [Google Scholar] [CrossRef] [PubMed]
- Harrison, W.O.; Hooper, R.R.; Wiesner, P.J.; Campbell, A.F.; Karney, W.W.; Reynolds, G.H.; Jones, O.G.; Holmes, K.K. A trial of minocycline given after exposure to prevent gonorrhea. N. Engl. J. Med. 1979, 300, 1074–1078. [Google Scholar] [CrossRef]
- Fiore, M.A.; Raisman, J.C.; Wong, N.R.H.; Hudson, A.O.; Wadsworth, C.B. Exploration of the Neisseria Resistome Reveals Resistance Mechanisms in Commensals That May Be Acquired by N. gonorrhoeae through Horizontal Gene Transfer. Antibiotics 2020, 9, 656. [Google Scholar] [CrossRef]
- Wadsworth, C.B.; Arnold, B.J.; Sater, M.R.A.; Grad, Y.H. Azithromycin Resistance through Interspecific Acquisition of an Epistasis-Dependent Efflux Pump Component and Transcriptional Regulator in Neisseria gonorrhoeae. Mbio 2018, 9, e01419-18. [Google Scholar] [CrossRef]

{kind=link}
| MSM ABs (Median (IQR)) | MSM No Abs [N (%)] | General Population [N (%)] | |
|---|---|---|---|
| Men | 32 (100) | 32 (100) | 10 (31.3) |
| Age categories (N persons per category (%) | |||
| 20–29 | 5 (15.6%) | 4 (12.5%) | 5 (15.6%) |
| 30–39 | 16 (50.0%) | 10 (31.3%) | 9 (28.1%) |
| 40–49 | 6 (18.8%) | 8 (25.0%) | 9 (28.1%) |
| 50–59 | 4 (12.5%) | 7 (21.9%) | 8 (25.0%) |
| 60–69 | 1 (3.1%) | 2 (6.3%) | 1 (3.1%) |
| 70–79 | 0 (0%) | 1 (3.1%) | 0 (0%) |
| Antibiotic exposure in the previous 6 months, n (%) | |||
| B-lactams | 25 (78.1%) | ||
| Macrolides | 19 (59.4%) | ||
| Fluoroquinolones | 2 (6.3%) | ||
| Others | 8 (25.0%) | ||
| N. subflava median MIC (ug/mL) # | |||
| Ceftriaxone | 0.035 (0.023–0.047) | 0.032 (0.023–0.06) | 0.036 (0.027–0.056) |
| Azithromycin | 3 (1.75–4) | 1.75 (0.38–6) | 3 (2–4) |
| Ciprofloxacin | 0.03 (0.01–0.38) | 0.02 (0.01–0.192) | 0.04 (0.02–0.19) |
| Antimicrobial read abundance (normalised read count) | |||
| Macrolides | 99 (0–539) | 0 (0–351) | 52 (0–167) |
| Betalactams | 0 (0–48) | 0 (0–65) | 0 (0–66) |
| Fluoroquinolones | 0 (0–83) | 0 (0–101) | 0 (0–63) |
| Tetracyclines | 0 (0–527) | 0 (0–415) | 0 (0–134) |
| Coef. (95% CI) | p-Value | |
|---|---|---|
| Azithromycin/macrolides | 0.19 (−1.06–1.45) | 0.759 |
| Ceftriaxone/betalactams | −0.03 (−0.66–0.37) | 0.926 |
| Antimicrobial Consumption/Gene Abundance | Coef. (95% CI) | p-Value |
|---|---|---|
| Azithromycin/macrolides | 0.23 (−0.30–0.77) | 0.393 |
| Ceftriaxone/betalactams | −0.03 (−0.66–0.37) | 0.926 |
| Fluoroquinolone/fluoroquinolone | −0.35 (−1.67–0.96) | 0.598 |
| Tetracycline/tetracycline | 0.03 (−0.60–0.67) | 0.916 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanbaelen, T.; Laumen, J.; Van Dijck, C.; De Block, T.; Manoharan-Basil, S.S.; Kenyon, C. Lack of Association between Antimicrobial Consumption and Antimicrobial Resistance in a HIV Preexposure Prophylaxis Population: A Cross-Sectional Study. Antibiotics 2024, 13, 188. https://doi.org/10.3390/antibiotics13020188
Vanbaelen T, Laumen J, Van Dijck C, De Block T, Manoharan-Basil SS, Kenyon C. Lack of Association between Antimicrobial Consumption and Antimicrobial Resistance in a HIV Preexposure Prophylaxis Population: A Cross-Sectional Study. Antibiotics. 2024; 13(2):188. https://doi.org/10.3390/antibiotics13020188
Chicago/Turabian StyleVanbaelen, Thibaut, Jolein Laumen, Christophe Van Dijck, Tessa De Block, Sheeba Santhini Manoharan-Basil, and Chris Kenyon. 2024. "Lack of Association between Antimicrobial Consumption and Antimicrobial Resistance in a HIV Preexposure Prophylaxis Population: A Cross-Sectional Study" Antibiotics 13, no. 2: 188. https://doi.org/10.3390/antibiotics13020188
APA StyleVanbaelen, T., Laumen, J., Van Dijck, C., De Block, T., Manoharan-Basil, S. S., & Kenyon, C. (2024). Lack of Association between Antimicrobial Consumption and Antimicrobial Resistance in a HIV Preexposure Prophylaxis Population: A Cross-Sectional Study. Antibiotics, 13(2), 188. https://doi.org/10.3390/antibiotics13020188