
Bioactive Compounds Isolated from a Marine Sponge Selectively Inhibit Neisseria gonorrhoeae
 ,
, 
Abstract

1. Introduction
2. Results
2.1. Determining Whether Methanol Sponge Extracts Display Antibacterial Activity
2.2. Antibiotic Profiling of High-Level Resistant Gonococcal Strain
2.3. Determining Whether EtOAc Sponge Extracts Inhibit Ng Strains
2.4. Determining Whether EtOAc Sponge Subfractions Inhibit Ng Strains
2.5. Evaluating the Synergistic Interaction of BH5 and Ceftriaxone Against Ng Strains
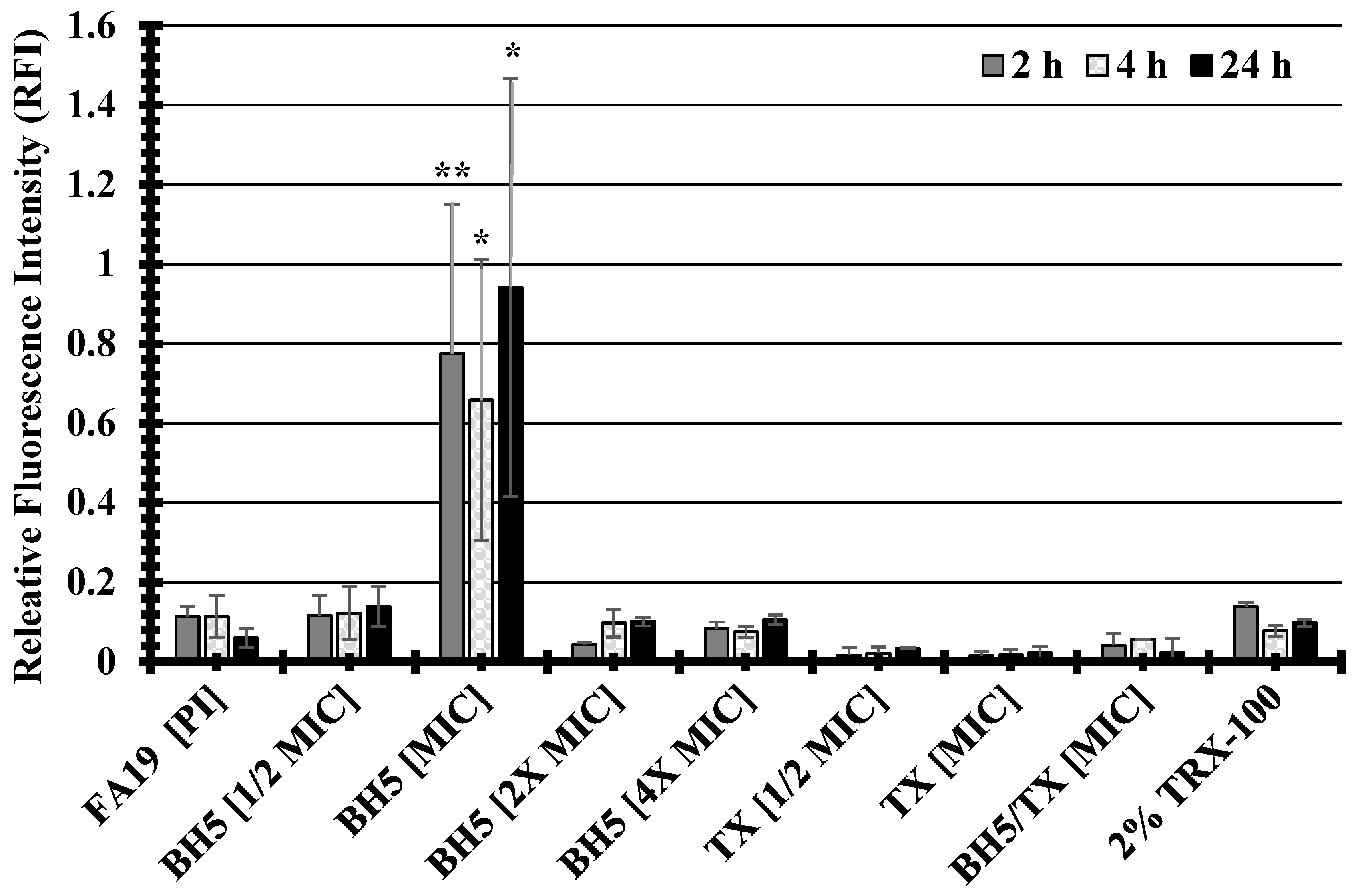
2.6. Examination of the Impact of BH5 on Bacterial Membrane Permeability
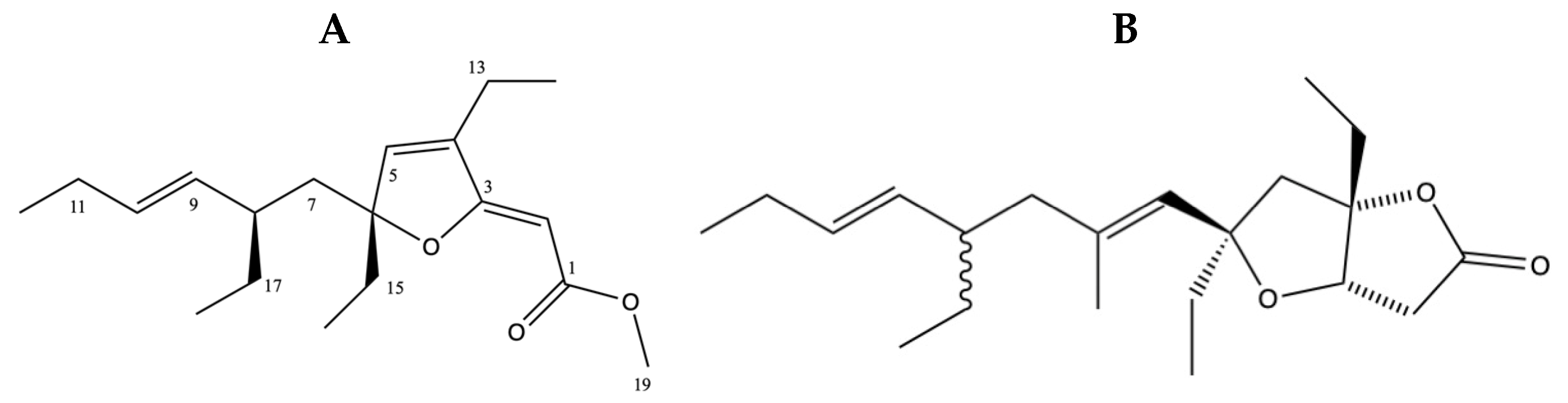
2.7. Structure Elucidation of 6-Desmethyl-6-Ethylspongosoritin A (1) and Plakortone B (2)
2.8. Evaluating the Synergistic Interaction of Plakortone B (2) and Ceftriaxone Against Ng Strains
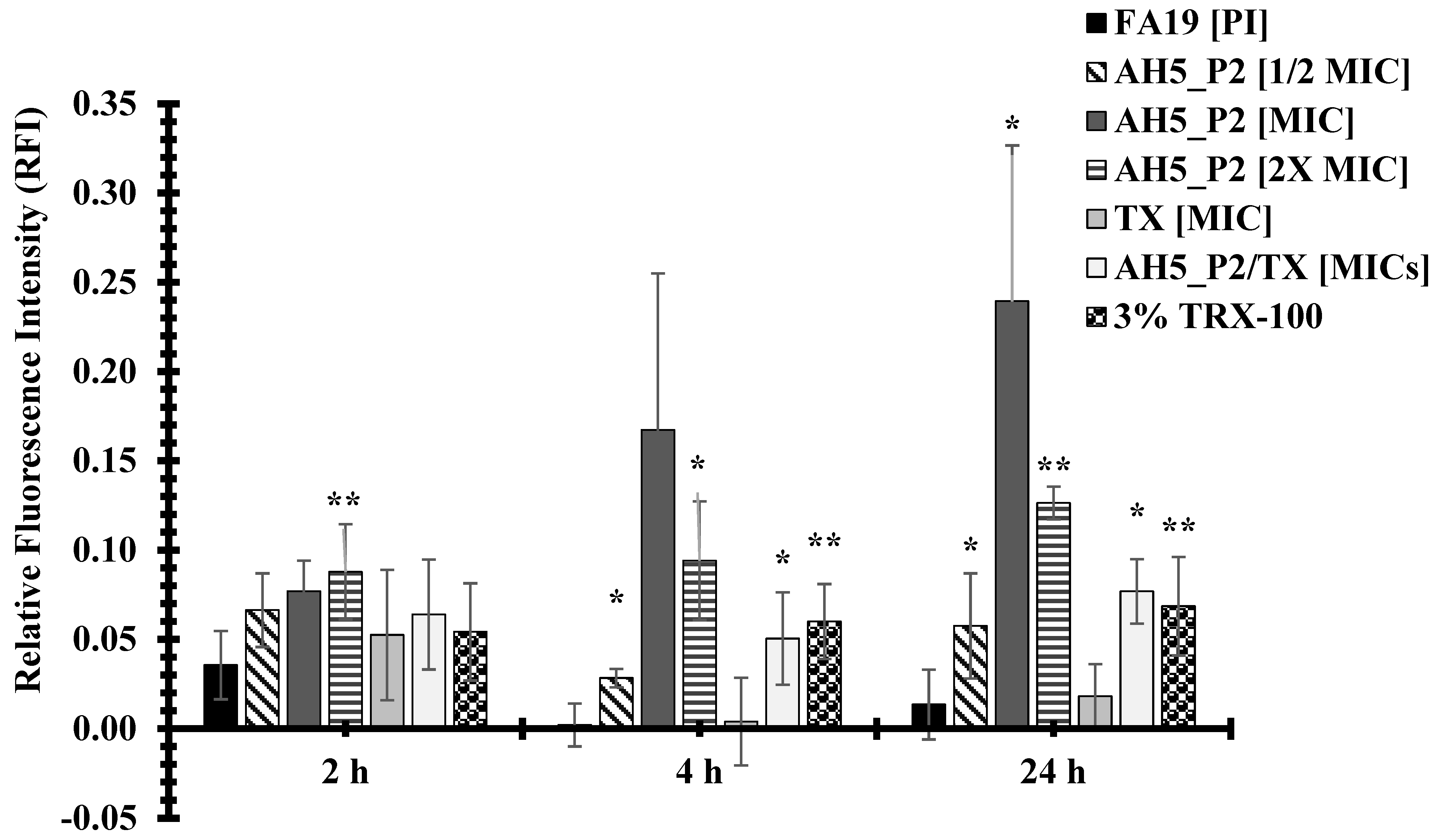
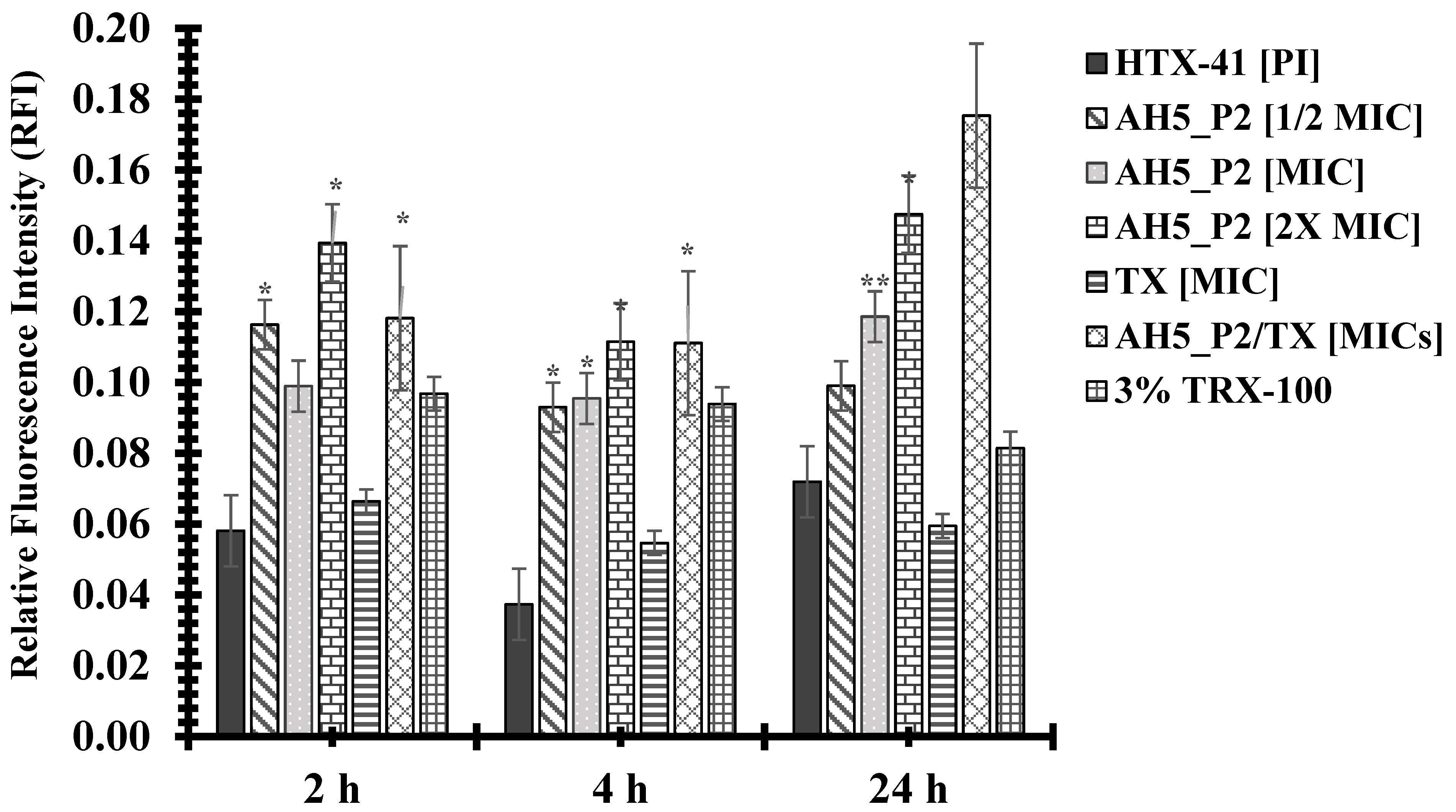
2.9. The Impact of AH5_P2 on Bacterial Membrane Permeability
3. Discussion
4. Materials and Methods
4.1. Collection and Isolation
4.2. NMR Analysis of Sponge Components
4.3. Bacterial Strains and Growth Conditions
4.4. Antibacterial Susceptibility Assay
4.5. Membrane Permeability Assays
4.6. Checkerboard Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cohen, M.S.; Hoffman, I.F.; Royce, R.A.; Kazembe, P.; Dyer, J.R.; Daly, C.C.; Zimba, D.; Vernazza, P.L.; Maida, M.; Fiscus, S.A.; et al. Reduction of concentration of HIV-1 in semen after treatment of urethritis: Implications for prevention of sexual transmission of HIV-1. AIDSCAP Malawi Research Group. Lancet 1997, 349, 1868–1873. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.; Weiss, K.; Mermin, J.; Dietz, P.; Rosenberg, E.S.; Gift, T.L.; Chesson, H.; Sullivan, P.S.; Lyles, C.; Bernstein, K.T.; et al. Proportion of Incident Human Immunodeficiency Virus Cases Among Men Who Have Sex With Men Attributable to Gonorrhea and Chlamydia: A Modeling Analysis. Sex. Transm. Dis. 2019, 46, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Drug Resistant Neisseria gonorrhoeae. 2019. Available online: https://www.cdc.gov/antimicrobial-resistance/media/pdfs/gonorrhea-508.pdf (accessed on 27 June 2024).
- Costa-Lourenco, A.; Barros Dos Santos, K.T.; Moreira, B.M.; Fracalanzza, S.E.L.; Bonelli, R.R. Antimicrobial resistance in Neisseria gonorrhoeae: History, molecular mechanisms and epidemiological aspects of an emerging global threat. Braz. J. Microbiol. 2017, 48, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.L.; Chaudhry, U.; Sachdev, D.; Sachdeva, P.N.; Bala, M.; Saluja, D. An insight into the drug resistance profile & mechanism of drug resistance in Neisseria gonorrhoeae. Indian J. Med. Res. 2011, 134, 419–431. [Google Scholar]
- Whittington, W.L.; Knapp, J.S. Trends in resistance of Neisseria gonorrhoeae to antimicrobial agents in the United States. Sex. Transm. Dis. 1988, 15, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Starnino, S.; Group, G.-L.W.; Galarza, P.; Carvallo, M.E.; Benzaken, A.S.; Ballesteros, A.M.; Cruz, O.M.; Hernandez, A.L.; Carbajal, J.L.; Borthagaray, G.; et al. Retrospective analysis of antimicrobial susceptibility trends (2000–2009) in Neisseria gonorrhoeae isolates from countries in Latin America and the Caribbean shows evolving resistance to ciprofloxacin, azithromycin and decreased susceptibility to ceftriaxone. Sex. Transm. Dis. 2012, 39, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Allen, V.G.; Mitterni, L.; Seah, C.; Rebbapragada, A.; Martin, I.E.; Lee, C.; Siebert, H.; Towns, L.; Melano, R.G.; Low, D.E. Neisseria gonorrhoeae treatment failure and susceptibility to cefixime in Toronto, Canada. JAMA 2013, 309, 163–170. [Google Scholar] [CrossRef]
- Tomberg, J.; Unemo, M.; Ohnishi, M.; Davies, C.; Nicholas, R.A. Identification of amino acids conferring high-level resistance to expanded-spectrum cephalosporins in the penA gene from Neisseria gonorrhoeae strain H041. Antimicrob. Agents Chemother. 2013, 57, 3029–3036. [Google Scholar] [CrossRef] [PubMed]
- Unemo, M.; Golparian, D.; Nicholas, R.; Ohnishi, M.; Gallay, A.; Sednaoui, P. High-level cefixime- and ceftriaxone-resistant Neisseria gonorrhoeae in France: Novel penA mosaic allele in a successful international clone causes treatment failure. Antimicrob. Agents Chemother. 2012, 56, 1273–1280. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Update to CDC’s Sexually transmitted diseases treatment guidelines, 2010: Oral cephalosporins no longer a recommended treatment for gonococcal infections. MMWR Morb. Mortal. Wkly. Rep. 2012, 61, 590–594. [Google Scholar]
- Yang, K.J.; Kojima, N.; Bristow, C.C.; Klausner, J.D. Effectiveness of Cefixime for the Treatment of Neisseria gonorrhoeae Infection at 3 Anatomic Sites: A Systematic Review and Meta-Analysis. Sex. Transm. Dis. 2023, 50, 131–137. [Google Scholar] [CrossRef]
- St Cyr, S.; Barbee, L.; Workowski, K.A.; Bachmann, L.H.; Pham, C.; Schlanger, K.; Torrone, E.; Weinstock, H.; Kersh, E.N.; Thorpe, P. Update to CDC’s Treatment Guidelines for Gonococcal Infection, 2020. MMWR. Morb. Mortal. Wkly. Rep. 2020, 69, 1911–1916. [Google Scholar] [CrossRef] [PubMed]
- Deguchi, T.; Yasuda, M.; Hatazaki, K.; Kameyama, K.; Horie, K.; Kato, T.; Mizutani, K.; Seike, K.; Tsuchiya, T.; Yokoi, S.; et al. New Clinical Strain of Neisseria gonorrhoeae with Decreased Susceptibility to Ceftriaxone, Japan. Emerg. Infect. Dis. 2016, 22, 142–144. [Google Scholar] [CrossRef]
- Vincent, L.R.; Kerr, S.R.; Tan, Y.; Tomberg, J.; Raterman, E.L.; Dunning Hotopp, J.C.; Unemo, M.; Nicholas, R.A.; Jerse, A.E. In Vivo-Selected Compensatory Mutations Restore the Fitness Cost of Mosaic penA Alleles That Confer Ceftriaxone Resistance in Neisseria gonorrhoeae. mBio 2018, 9, 01905-17. [Google Scholar] [CrossRef]
- Ohnishi, M.; Golparian, D.; Shimuta, K.; Saika, T.; Hoshina, S.; Iwasaku, K.; Nakayama, S.; Kitawaki, J.; Unemo, M. Is Neisseria gonorrhoeae initiating a future era of untreatable gonorrhea?: Detailed characterization of the first strain with high-level resistance to ceftriaxone. Antimicrob. Agents Chemother. 2011, 55, 3538–3545. [Google Scholar] [CrossRef]
- Melnyk, A.H.; Wong, A.; Kassen, R. The fitness costs of antibiotic resistance mutations. Evol. Appl. 2015, 8, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Abdelmohsen, U.R.; Cheng, C.; Reimer, A.; Kozjak-Pavlovic, V.; Ibrahim, A.K.; Rudel, T.; Hentschel, U.; Edrada-Ebel, R.; Ahmed, S.A. Antichlamydial sterol from the Red Sea sponge Callyspongia aff. implexa. Planta Med. 2015, 81, 382–387. [Google Scholar] [CrossRef]
- Barzkar, N.; Sukhikh, S.; Babich, O. A comprehensive review of marine sponge metabolites, with emphasis on Neopetrosia sp. Int. J. Biol. Macromol. 2024, 280, 135823. [Google Scholar] [CrossRef] [PubMed]
- Beesoo, R.; Bhagooli, R.; Neergheen-Bhujun, V.S.; Li, W.W.; Kagansky, A.; Bahorun, T. Antibacterial and antibiotic potentiating activities of tropical marine sponge extracts. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2017, 196, 81–90. [Google Scholar] [CrossRef]
- Gogineni, V.; Hamann, M.T. Marine natural product peptides with therapeutic potential: Chemistry, biosynthesis, and pharmacology. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 81–196. [Google Scholar] [CrossRef]
- Mehbub, M.F.; Lei, J.; Franco, C.; Zhang, W. Marine sponge derived natural products between 2001 and 2010: Trends and opportunities for discovery of bioactives. Mar. Drugs 2014, 12, 4539–4577. [Google Scholar] [CrossRef]
- Nalini, S.; Sandy Richard, D.; Mohammed Riyaz, S.U.; Kavitha, G.; Inbakandan, D. Antibacterial macro molecules from marine organisms. Int. J. Biol. Macromol. 2018, 115, 696–710. [Google Scholar] [CrossRef] [PubMed]
- Pech-Puch, D.; Perez-Povedano, M.; Gomez, P.; Martinez-Guitian, M.; Lasarte-Monterrubio, C.; Vazquez-Ucha, J.C.; Novoa-Olmedo, M.L.; Guillen-Hernandez, S.; Villegas-Hernandez, H.; Bou, G.; et al. Marine Organisms from the Yucatan Peninsula (Mexico) as a Potential Natural Source of Antibacterial Compounds. Mar. Drugs 2020, 18, 369. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.D.; Cobar, O.M.; Padilla, O.L. The calyxolanes: New 1,3-diphenylbutanoid metabolites isolated from the Caribbean marine sponge Calyx podatypa. J. Nat. Prod. 1997, 60, 915–917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dong, M.; Chen, J.; Wang, H.; Tenney, K.; Crews, P. Bioactive Secondary Metabolites from the Marine Sponge Genus Agelas. Mar. Drugs 2017, 15, 351. [Google Scholar] [CrossRef]
- Sato, K.; Horibe, K.; Amano, K.; Mitusi-Saito, M.; Hori, M.; Matsunaga, S.; Fusetani, N.; Ozaki, H.; Karaki, H. Membrane permeabilization induced by discodermin A, a novel marine bioactive peptide. Toxicon 2001, 39, 259–264. [Google Scholar] [CrossRef]
- Mehbub, M.F.; Perkins, M.V.; Zhang, W.; Franco, C.M.M. New marine natural products from sponges (Porifera) of the order Dictyoceratida (2001 to 2012); a promising source for drug discovery, exploration and future prospects. Biotechnol. Adv. 2016, 34, 473–491. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.D.d.; Scheuer, P.J. Manoalide, an antibiotic sesterterpenoid from the marine sponge Luffariella variabilis (polejaeff). Tetrahedron Lett. 1980, 21, 1611–1614. [Google Scholar] [CrossRef]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep. 1996, 13, 75–125. [Google Scholar] [CrossRef]
- Costantino, V.; Della Sala, G.; Saurav, K.; Teta, R.; Bar-Shalom, R.; Mangoni, A.; Steindler, L. Plakofuranolactone as a Quorum Quenching Agent from the Indonesian Sponge Plakortis cf. lita. Mar. Drugs 2017, 15, 59. [Google Scholar] [CrossRef]
- Jimenez-Romero, C.; Rode, J.E.; Perez, Y.M.; Franzblau, S.G.; Rodriguez, A.D. Exploring the Sponge Consortium Plakortis symbiotica-Xestospongia deweerdtae as a Potential Source of Antimicrobial Compounds and Probing the Pharmacophore for Antituberculosis Activity of Smenothiazole A by Diverted Total Synthesis. J. Nat. Prod. 2017, 80, 2295–2303. [Google Scholar] [CrossRef]
- Manzo, E.; Ciavatta, M.L.; Melck, D.; Schupp, P.; de Voogd, N.; Gavagnin, M. Aromatic cyclic peroxides and related keto-compounds from the Plakortis sp. component of a sponge consortium. J. Nat. Prod. 2009, 72, 1547–1551. [Google Scholar] [CrossRef] [PubMed]
- Cafieri, F.; Fattorusso, E.; Taglialatela-Scafati, O.; Di Rosa, M.; Ianaro, A. Metabolites from the sponge plakortis simplex. II.1: Isolation of four bioactive lactone compounds and of a novel related amino acid. Tetrahedron 1999, 55, 13831–13840. [Google Scholar] [CrossRef]
- Patil, A.D.; Freyer, A.J.; Bean, M.F.; Carte, B.K.; Westley, J.W.; Johnson, R.K. The Plakortones, Novel Bicyclic Lactones from the Sponge Plakortis halichondrioides: Activators of Cardiac SR-Ca2+ -Pumping ATPase. Tetrahedron 1996, 52, 377–394. [Google Scholar] [CrossRef]
- Ohnishi, M.; Saika, T.; Hoshina, S.; Iwasaku, K.; Nakayama, S.; Watanabe, H.; Kitawaki, J. Ceftriaxone-resistant Neisseria gonorrhoeae, Japan. Emerg. Infect. Dis. 2011, 17, 148–149. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.A.; Quintela, A.L.; Ferreira, E.G.; Sousa, T.S.; Pinto, F.; Hajdu, E.; Carvalho, M.S.; Salani, S.; Rocha, D.D.; Wilke, D.V.; et al. Cytotoxic Plakortides from the Brazilian Marine Sponge Plakortis angulospiculatus. J. Nat. Prod. 2015, 78, 996–1004. [Google Scholar] [CrossRef]
- Stierle, D.B.; Faulkner, D.J. Metabolites of three marine sponges of the genus Plakortis. J. Org. Chem. 1980, 45, 3396–3401. [Google Scholar] [CrossRef]
- Le Van, A.; Rahman, N.; Sandy, R.; Dozier, N.; Smith, H.J.; Martin, M.J.; Bartlett, K.V.; Harncharoenkul, K.; Nanava, A.; Akhvlediani, T.; et al. Common Patterns and Unique Threats in Antimicrobial Resistance as Demonstrated by Global Gonococcal Surveillance. Emerg. Infect. Dis. 2024, 30, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Melendez, J.H.; Edwards, V.L.; Muniz Tirado, A.; Hardick, J.; Mehta, A.; Aluvathingal, J.; D’Mello, A.; Gaydos, C.A.; Manabe, Y.C.; Tettelin, H. Local emergence and global evolution of Neisseria gonorrhoeae with high-level resistance to azithromycin. Antimicrob. Agents Chemother. 2024, e0092724. [Google Scholar] [CrossRef]
- Wi, T.; Lahra, M.M.; Ndowa, F.; Bala, M.; Dillon, J.R.; Ramon-Pardo, P.; Eremin, S.R.; Bolan, G.; Unemo, M. Antimicrobial resistance in Neisseria gonorrhoeae: Global surveillance and a call for international collaborative action. PLoS Med. 2017, 14, e1002344. [Google Scholar] [CrossRef]
- Lin, E.Y.; Adamson, P.C.; Klausner, J.D. Epidemiology, Treatments, and Vaccine Development for Antimicrobial-Resistant Neisseria gonorrhoeae: Current Strategies and Future Directions. Drugs 2021, 81, 1153–1169. [Google Scholar] [CrossRef] [PubMed]
- Hooshiar, M.H.; Sholeh, M.; Beig, M.; Azizian, K.; Kouhsari, E. Corrigendum: Global trends of antimicrobial resistance rates in Neisseria gonorrhoeae: A systematic review and meta-analysis. Front. Pharmacol. 2024, 15, 1465628. [Google Scholar] [CrossRef]
- Varijakzhan, D.; Loh, J.Y.; Yap, W.S.; Yusoff, K.; Seboussi, R.; Lim, S.E.; Lai, K.S.; Chong, C.M. Bioactive Compounds from Marine Sponges: Fundamentals and Applications. Mar. Drugs 2021, 19, 246. [Google Scholar] [CrossRef]
- Keita, K.; Darkoh, C.; Okafor, F. Secondary plant metabolites as potent drug candidates against antimicrobial-resistant pathogens. SN Appl. Sci. 2022, 4, 209. [Google Scholar] [CrossRef]
- Amin, M.U.; Khurram, M.; Khattak, B.; Khan, J. Antibiotic additive and synergistic action of rutin, morin and quercetin against methicillin resistant Staphylococcus aureus. BMC Complement. Altern. Med. 2015, 15, 59. [Google Scholar] [CrossRef]
- Williams, D.; Perry, D.; Carraway, J.; Simpson, S.; Uwamariya, P.; Christian, O.E. Antigonococcal Activity of (+)-Medicarpin. ACS Omega 2021, 6, 15274–15278. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, X.; Li, J.; Li, P.; de Voogd, N.J.; Ni, X.; Jin, X.; Yao, X.; Li, P.; Li, G. Cytotoxic polyketide derivatives from the South China Sea sponge Plakortis simplex. J. Nat. Prod. 2013, 76, 600–606. [Google Scholar] [CrossRef]
- Bhattacharya, A.K.; Chand, H.R.; John, J.; Deshpande, M.V. Clerodane type diterpene as a novel antifungal agent from Polyalthia longifolia var. pendula. Eur. J. Med. Chem. 2015, 94, 1–7. [Google Scholar] [CrossRef]
- Mazur, M.; Maslowiec, D. Antimicrobial Activity of Lactones. Antibiotics 2022, 11, 1327. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Description | Abbreviation |
|---|---|---|
| N. gonorrhoeae FA19 | wildtype | WT |
| FA19 penA41 | High level ceftriaxone-resistant strain [9] | HTX-41 |
| Escherichia coli | Gram-negative bacteria | E. coli |
| Staphylococcus aureus | Gram-positive bacteria | S. aureus |
| Fractions, MF | Minimum Inhibitory Concentration (mg/mL) | |||||
|---|---|---|---|---|---|---|
| N. gonorrhoeae | E. coli | S. aureus | ||||
| 4 h | 24 h | 4 h | 24 h | 4 h | 24 h | |
| MF-1 | 2.93 (±0.7) | 1.56 (±0.3) | NI | NI | NI | NI |
| MF-2 | 6.25 (±0.0) | 5.86 (±0.4) | NI | NI | NI | NI |
| MF-3 | 2.95 (±0.1) | 2.04 (±0.2) | NI | NI | NI | NI |
| MF-4 | 4.04 (±0.5) | 2.6 (±0.2) | NI | NI | NI | NI |
| MF-5 | 7.55 (±1.1) | 4.69 (±0.5) | NI | NI | NI | NI |
| MF-6 | 1.17 (±0.1) | 1.04 (±0.1) | NI | NI | NI | NI |
| MF-7 | 1.56 (±0.0) | 0.98 (±0.1) | NI | NI | NI | NI |
| Time (h) | Minimum Inhibitory Concentration (μg/mL) | |
|---|---|---|
| FA19WT | HTX-41 | |
| 4 | 0.08 | 6 |
| 24 | 0.08 | 2 |
| Time (h) | Minimum Inhibitory Concentration (mg/mL) | |
|---|---|---|
| FA19WT | HTX-41 | |
| 4 | 0.122 | 0.052 |
| 24 | 0.178 | 0.084 |
| Fractions | Minimum Inhibitory Concentration (mg/mL) | |||
|---|---|---|---|---|
| FA19WT | HTX-41 | |||
| 4 h | 24 h | 4 h | 24 h | |
| A | 0.059 | 0.051 | <0.019 | <0.019 |
| B | 0.34 | 0.287 | 0.110 | 0.086 |
| C | 0.115 | 0.074 | 0.085 | 0.077 |
| D | 0.277 | 0.235 | 0.230 | 0.245 |
| E | 0.434 | 0.244 | 0.2 | 0.175 |
| F | 0.246 | 0.292 | 0.230 | 0.170 |
| Fractions | Minimum Inhibitory Concentrations (mg/mL) | |||
|---|---|---|---|---|
| 4 h | 24 h | |||
| FA19WT | HTX-41 | FA19WT | HTX-41 | |
| AH1 | NI | 15 | NI | 15 |
| AH2 | NI | 35 | NI | 35 |
| AH3 | NI | 1.2 | NI | 35 |
| AH4 | 0.1 | 0.078 | 0.082 | 0.078 |
| AH5 | 0.136 | 0.196 | 0.136 | 0.152 |
| AH6 | 0.078 | 0.1 | 0.078 | 0.095 |
| AH7 | 0.781 | 0.416 | 0.729 | 0.494 |
| Fractions | Minimum Inhibitory Concentrations (mg/mL) | |||
|---|---|---|---|---|
| 4 h | 24 h | |||
| FA19WT | HTX-41 | FA19WT | HTX-41 | |
| BH1 | NI | NI | NI | NI |
| BH2 | NI | NI | NI | NI |
| BH3 | 0.038 | 0.052 | 0.074 | 0.069 |
| BH4 | 0.037 | 0.043 | 0.052 | 0.046 |
| BH5 | 0.046 | 0.060 | 0.156 | 0.049 |
| BH6 | 0.208 | 0.156 | 0.174 | 0.156 |
| Time (h) | Strains | Agent | MIC (mg/mL) | FIC | FICI | Interpretation | |
|---|---|---|---|---|---|---|---|
| Alone | Combined | ||||||
| 4 | FA19WT | BH5 | 0.125 | 0.0521 | 0.417 | 0.469 | Synergy |
| TX | 0.000038 | 0.000002 | 0.053 | ||||
| HTX-41 | BH5 | 0.072 | 0.019 | 0.26 | 0.416 | Synergy | |
| TX | 0.025 | 0.004 | 0.155 | ||||
| 24 | FA19WT | BH5 | 0.125 | 0.0313 | 0.250 | 0.376 | Synergy |
| TX | 0.000015 | 0.000002 | 0.125 | ||||
| HTX-41 | BH5 | 0.109 | 0.000063 | 1 | 1.011 | Additive | |
| TX | 0.00283 | 0.000031 | 0.011 | ||||
| Experimental | Theoretical | |
|---|---|---|
| 1 | 167.0 | 166.0 |
| 2 | 83.3 | 84.4 |
| 3 | 172.2 | 171.5 |
| 4 | 140.3 | 139.9 |
| 5 | 139.3 | 139.7 |
| 6 | 97.8 | 97.2 |
| 7 | 43.5 | 43.6 |
| 8 | 39.7 | 39.9 |
| 9 | 134.0 | 133.5 |
| 10 | 132.1 | 131.9 |
| 11 | 25.6 | 25.9 |
| 12 | 14.0 | 14.1 |
| 13 | 18.5 | 18.7 |
| 14 | 11.8 | 11.9 |
| 15 | 32.3 | 32.4 |
| 16 | 8.0 | 7.9 |
| 17 | 29.4 | 29.6 |
| 18 | 11.4 | 11.5 |
| 19 | 50.5 | 50.1 |
| Fractions | Minimum Inhibitory Concentrations (mg/mL) | |||
|---|---|---|---|---|
| 4 h | 24 h | |||
| FA19WT | HTX-41 | FA19WT | HTX-41 | |
| AH5_P1 | NI | 1.75 | NI | 1.75 |
| AH5_P2 | 0.175 | 0.066 | 0.131 | 0.131 |
| BH5_P1 | NI | NI | NI | NI |
| BH5_P2 | 0.5 | 0.333 | 0.5 | 0.5 |
| Time (h) | Strain | Agent (mg/mL) | MIC (mg/mL) | FIC | FICI | Interpretation | |
|---|---|---|---|---|---|---|---|
| Alone | Combined | ||||||
| 4 | FA19WT | BH5_P2 | 0.5 | 0.25 | 0.5 | 0.567 | Synergy |
| TX | 0.0009 | 0.0000625 | 0.067 | ||||
| BH5_P2 | 0.5 | 0.375 | 0.75 | 0.783 | Additive | ||
| TX | 0.0009 | 0.0000313 | 0.033 | ||||
| HTX | BH5_P2 | 0.5 | 0.5 | 1 | 1.016 | Additive | |
| TX | 0.004 | 0.000063 | 0.016 | ||||
| BH5_P2 | 0.5 | 0.5 | 1 | 1.008 | Additive | ||
| TX | 0.004 | 0.000031 | 0.008 | ||||
| 24 | FA19WT | BH5_P2 | 0.5 | 0.25 | 0.5 | 0.5 | Synergy |
| TX | 0.01 | 0.0000047 | 0.0003 | ||||
| BH5_P2 | 0.5 | 0.5 | 1 | 1 | Additive | ||
| TX | 0.01 | 0.0000023 | 0.00015 | ||||
| HTX | BH5_P2 | 0.375 | 0.5 | 1.33 | 1.3 | Additive | |
| TX | 0.004 | 0.000065 | 0.016 | ||||
| BH5_P2 | 0.375 | 0.5 | 1.33 | 1.3 | Additive | ||
| TX | 0.004 | 0.0000313 | 0.008 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christian, O.E.; Perry, D.A.; Telchy, A.I.; Walton, P.N.; Williams, D. Bioactive Compounds Isolated from a Marine Sponge Selectively Inhibit Neisseria gonorrhoeae. Antibiotics 2024, 13, 1229. https://doi.org/10.3390/antibiotics13121229
Christian OE, Perry DA, Telchy AI, Walton PN, Williams D. Bioactive Compounds Isolated from a Marine Sponge Selectively Inhibit Neisseria gonorrhoeae. Antibiotics. 2024; 13(12):1229. https://doi.org/10.3390/antibiotics13121229
Chicago/Turabian StyleChristian, Omar E., Dreyona A. Perry, Alaa I. Telchy, Preston N. Walton, and Daniel Williams. 2024. "Bioactive Compounds Isolated from a Marine Sponge Selectively Inhibit Neisseria gonorrhoeae" Antibiotics 13, no. 12: 1229. https://doi.org/10.3390/antibiotics13121229
APA StyleChristian, O. E., Perry, D. A., Telchy, A. I., Walton, P. N., & Williams, D. (2024). Bioactive Compounds Isolated from a Marine Sponge Selectively Inhibit Neisseria gonorrhoeae. Antibiotics, 13(12), 1229. https://doi.org/10.3390/antibiotics13121229