
Validation of Recombinase Polymerase Amplification with In-House Lateral Flow Assay for mcr-1 Gene Detection of Colistin Resistant Escherichia coli Isolates

 , and
, and
Abstract
1. Introduction
2. Results
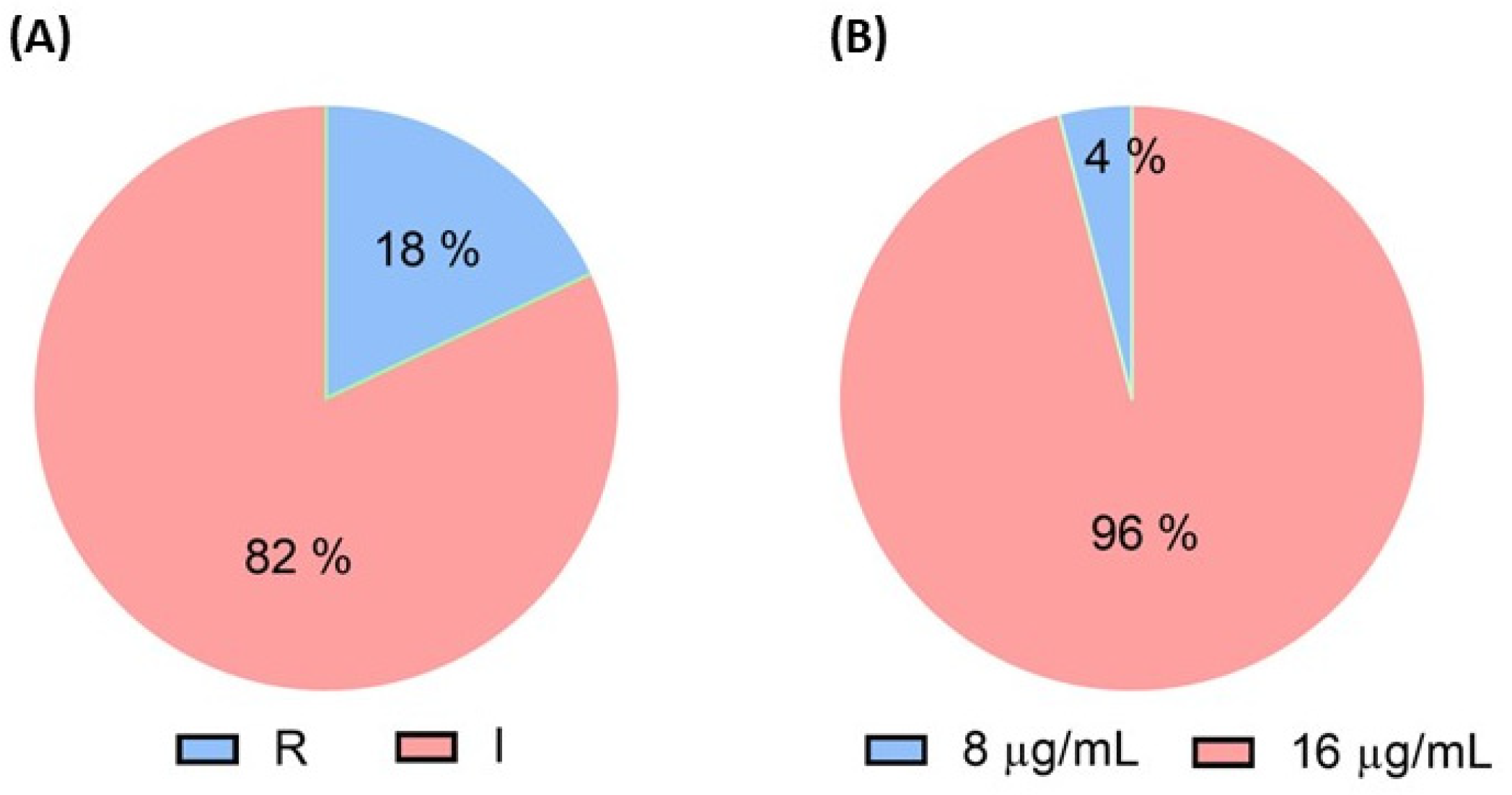
2.1. Phenotypic Detection of Colistin-Resistant E. coli Isolated from Antibiotics Using Pig Farms
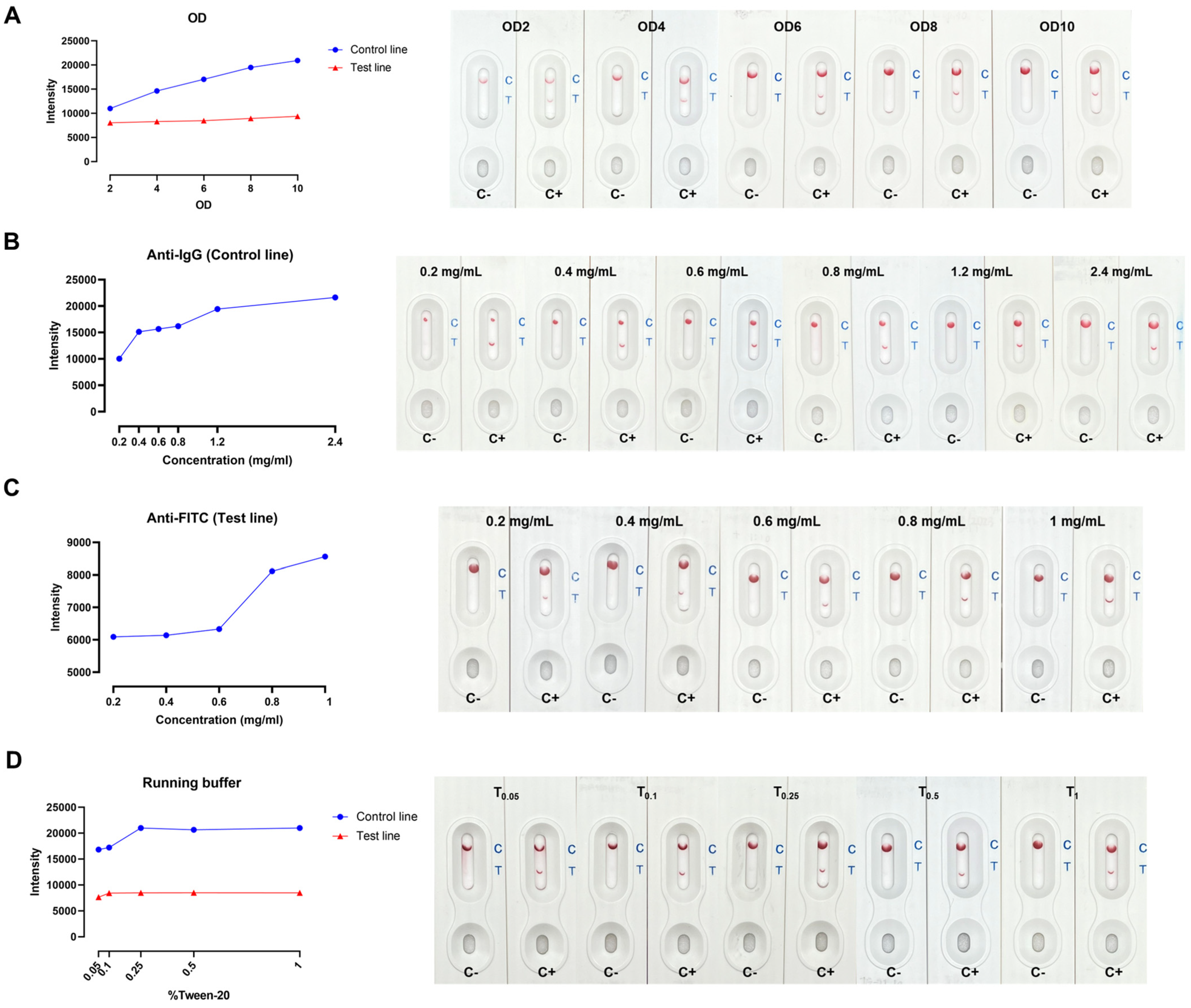
2.2. Optimization of Key Parameters in In-House LFA Development
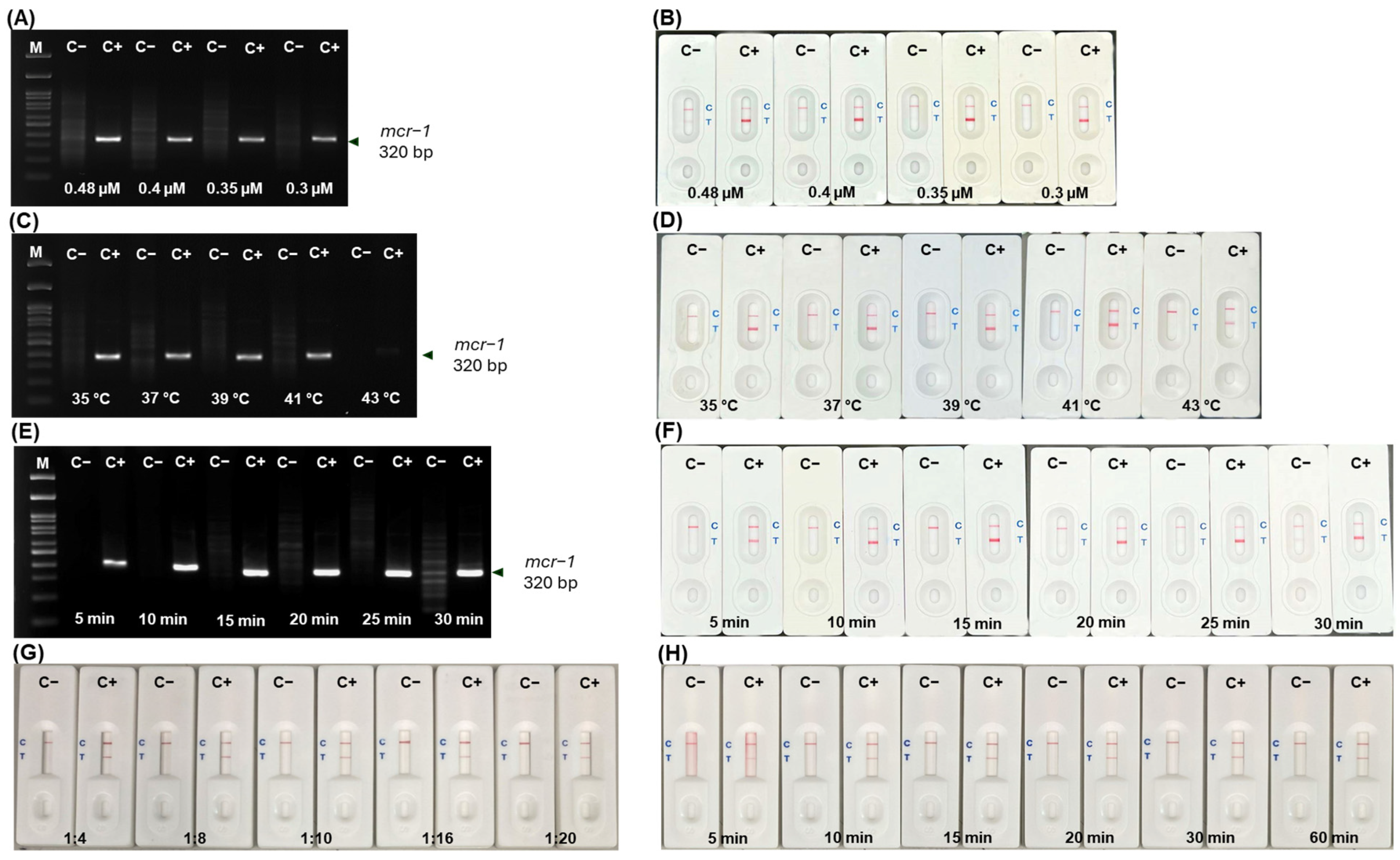
2.3. Optimization of mcr-1 Primer Concentrations and Conditions in RPA-in-House LFA
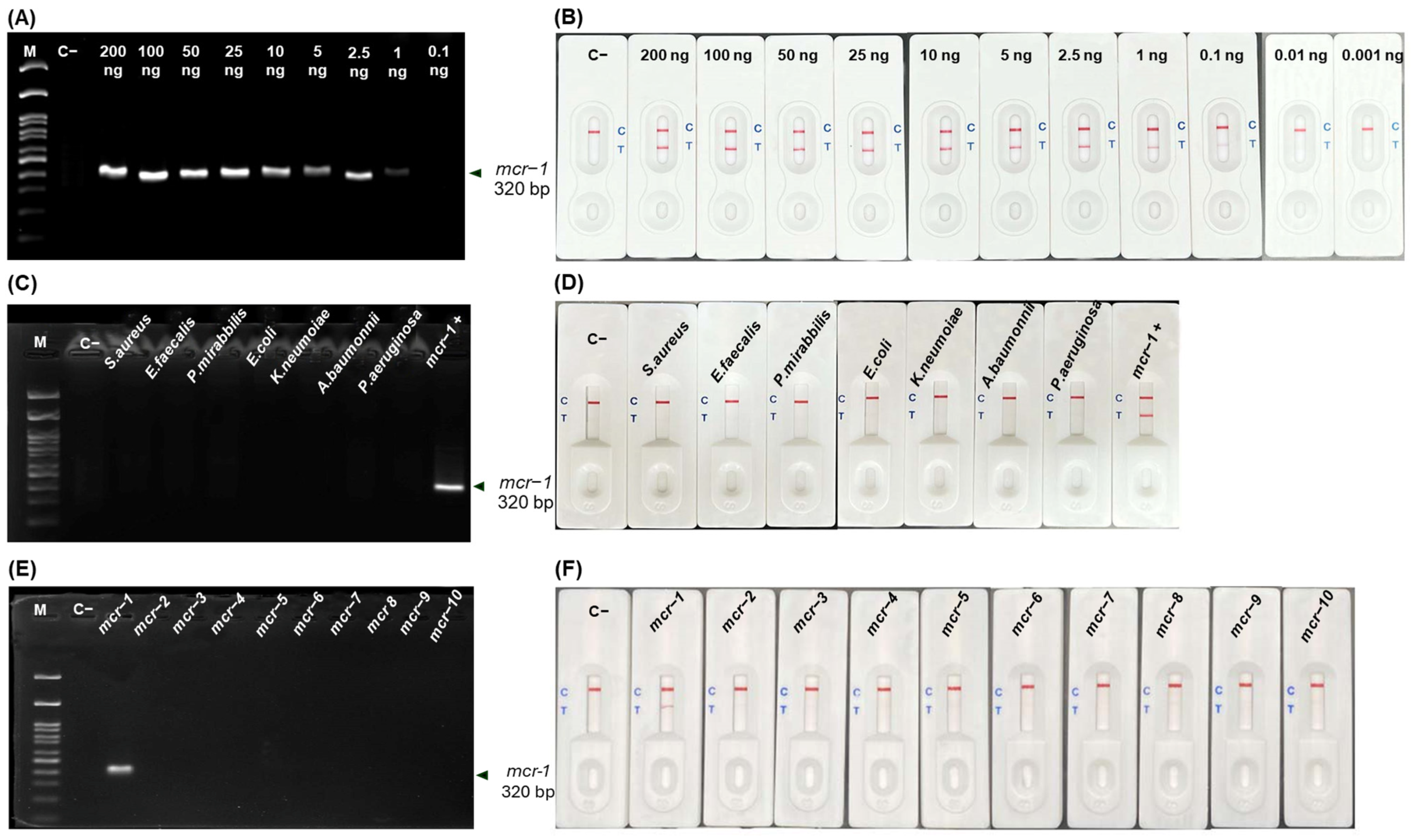
2.4. Limit of Detection (LOD) and Cross-Reactivity of RPA-LFA
2.5. Detection of mcr-1 Gene from Pig and Clinical Samples by RPA-LFA
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains for Detection of Colistin Resistance by Phenotyping, PCR and RPA-LFA
4.2. Phenotypic Colistin Resistance Screening
4.3. DNA Extraction
4.4. PCR for MCR-1 Gene Detection
4.5. In-House LFA Optimization
4.6. RPA-LFA Optimization
4.7. Limit of Detection (LOD) and Specificity of RPA-LFA
4.8. Validation of the RPA-LFA
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Sayed Ahmed, M.A.E.-G.; Zhong, L.-L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.-B. Colistin and its role in the era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef]
- Mondal, A.H.; Khare, K.; Saxena, P.; Debnath, P.; Mukhopadhyay, K.; Yadav, D. A Review on colistin resistance: An antibiotic of last resort. Microorganisms 2024, 12, 772. [Google Scholar] [CrossRef]
- Rhouma, M.; Madec, J.-Y.; Laxminarayan, R. Colistin: From the shadows to a one health approach for addressing antimicrobial resistance. Int. J. Antimicrob. Agents 2023, 61, 106713. [Google Scholar] [CrossRef]
- Velkov, T.; Roberts, K.D.; Nation, R.L.; Thompson, P.E.; Li, J. Pharmacology of polymyxins: New insights into an ‘old’class of antibiotics. Future Microbiol. 2013, 8, 711–724. [Google Scholar] [CrossRef]
- Anyanwu, M.U.; Jaja, I.F.; Okpala, C.O.R.; Njoga, E.O.; Okafor, N.A.; Oguttu, J.W. Mobile colistin resistance (mcr) gene-containing organisms in poultry sector in low-and middle-income countries: Epidemiology, characteristics, and one health control strategies. Antibiotics 2023, 12, 1117. [Google Scholar] [CrossRef]
- Hussein, N.H.; Al-Kadmy, I.M.S.; Taha, B.M.; Hussein, J.D. Mobilized colistin resistance (mcr) genes from 1 to 10: A comprehensive review. Mol. Biol. Rep. 2021, 48, 2897–2907. [Google Scholar] [CrossRef]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase polymerase amplification: Basics, applications and recent advances. Trac Trends Anal. Chem. 2018, 98, 19–35. [Google Scholar] [CrossRef]
- M100-S31; Performance Standards for Antimicrobial Susceptibility Testing. Thirty-One Informational Supplement. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021.
- Tan, T.Y.; Ng, S.Y. Comparison of Etest, Vitek and agar dilution for susceptibility testing of colistin. Clin. Microbiol. Infect. 2007, 13, 541–544. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. Recommendations for MIC Determination of Colistin (Polymyxin E) as Recommended by the Joint CLSI-EUCAST Polymyxin Breakpoints Working Group; EUCAST: Växjö, Sweden, 2016. [Google Scholar]
- Daniels, J.B.; Campbell, D.; Boyd, S.; Ansari, U.; Lutgring, J.; Rasheed, J.K.; Halpin, A.L.; Sjölund-Karlsson, M. Development and validation of a clinical laboratory improvement amendments-compliant multiplex real-time PCR assay for detection of mcr genes. Microb. Drug Resist. 2019, 25, 991–996. [Google Scholar] [CrossRef]
- Zou, D.; Huang, S.; Lei, H.; Yang, Z.; Su, Y.; He, X.; Zhao, Q.; Wang, Y.; Liu, W.; Huang, L. Sensitive and rapid detection of the plasmid-encoded colistin-resistance gene mcr-1 in Enterobacteriaceae isolates by loop-mediated isothermal amplification. Front. Microbiol. 2017, 8, 2356. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, H.; Pan, Q.; Wang, J.; Jiao, X.; Zhang, Y. Development and evaluation of rapid and accurate one-tube RPA-CRISPR-Cas12b-based detection of mcr-1 and tet (x4). Appl. Microbiol. Biotechnol. 2024, 108, 345. [Google Scholar] [CrossRef]
- Bahadır, E.B.; Sezgintürk, M.K. Lateral flow assays: Principles, designs and labels. TrAC Trends Anal. Chem. 2016, 82, 286–306. [Google Scholar] [CrossRef]
- Koczula, K.M.; Gallotta, A. Lateral flow assays. Essays Biochem. 2016, 60, 111–120. [Google Scholar] [CrossRef]
- Zheng, T.; Li, X.; Si, Y.; Wang, M.; Zhou, Y.; Yang, Y.; Liang, N.; Ying, B.; Wu, P. Specific lateral flow detection of isothermal nucleic acid amplicons for accurate point-of-care testing. Biosens. Bioelectron. 2023, 222, 114989. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Z.; Feng, Y.; Hu, H.; Yu, Y.; Qiu, L.; Liu, H.; Guo, Z.; Huang, J.; Du, C. Molecular detection of the mcr genes by multiplex PCR. Infect. Drug Resist. 2020, 2020, 3463–3468. [Google Scholar] [CrossRef]
- Lu, C.; Wang, J.; Pan, L.; Gu, X.; Lu, W.; Chen, D.; Zhang, C.; Ye, Q.; Xiao, C.; Liu, P. Rapid detection of multiple resistance genes to last-resort antibiotics in Enterobacteriaceae pathogens by recombinase polymerase amplification combined with lateral flow dipstick. Front. Microbiol. 2023, 13, 1062577. [Google Scholar] [CrossRef]
- Kanokudom, S.; Assawakongkarat, T.; Akeda, Y.; Ratthawongjirakul, P.; Chuanchuen, R.; Chaichanawongsaroj, N. Rapid detection of extended spectrum β-lactamase producing Escherichia coli isolated from fresh pork meat and pig cecum samples using multiplex recombinase polymerase amplification and lateral flow strip analysis. PLoS ONE 2021, 16, e0248536. [Google Scholar] [CrossRef]
- Khine, N.O.; Lugsomya, K.; Niyomtham, W.; Pongpan, T.; Hampson, D.J.; Prapasarakul, N. Longitudinal monitoring reveals persistence of colistin-resistant Escherichia coli on a pig farm following cessation of colistin use. Front. Vet. Sci. 2022, 9, 845746. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Zhang, R.; Chen, Y.; Shen, Y.; Hu, F.; Liu, D.; Lu, J.; Guo, Y.; Xia, X. Changes in colistin resistance and mcr-1 abundance in Escherichia coli of animal and human origins following the ban of colistin-positive additives in china: An epidemiological comparative study. Lancet Infect. Dis. 2020, 20, 1161–1171. [Google Scholar] [CrossRef]
- Peng, Z.; Zhang, X.; Li, X.; Hu, Z.; Li, Z.; Jia, C.; Dai, M.; Tan, C.; Chen, H.; Wang, X. Characteristics of colistin-resistant Escherichia coli from pig farms in central China. Anim. Dis. 2021, 1, 1–11. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, R.; Schwarz, S.; Wu, C.; Shen, J.; Walsh, T.R.; Wang, Y. Farm animals and aquaculture: Significant reservoirs of mobile colistin resistance genes. Environ. Microbiol. 2020, 22, 2469–2484. [Google Scholar] [CrossRef]
- Bastidas-Caldes, C.; de Waard, J.H.; Salgado, M.S.; Villacís, M.J.; Coral-Almeida, M.; Yamamoto, Y.; Calvopiña, M. Worldwide prevalence of mcr-mediated colistin-resistance Escherichia coli in isolates of clinical samples, healthy humans, and livestock—A systematic review and meta-analysis. Pathogens 2022, 11, 659. [Google Scholar] [CrossRef]
- Zhong, L.-L.; Zhou, Q.; Tan, C.-Y.; Roberts, A.P.; El-Sayed Ahmed, M.A.E.-G.; Chen, G.; Dai, M.; Yang, F.; Xia, Y.; Liao, K. Multiplex loop-mediated isothermal amplification (multi-lamp) assay for rapid detection of mcr-1 to mcr-5 in colistin-resistant bacteria. Infect. Drug Resist. 2019, 2019, 1877–1887. [Google Scholar] [CrossRef]
- Xu, J.; Wang, X.; Yang, L.; Kan, B.; Lu, X. Rapid detection of mcr-1 by recombinase polymerase amplification. J. Med. Microbiol. 2018, 67, 1682–1688. [Google Scholar] [CrossRef]
- Li, J.; Macdonald, J.; von Stetten, F. Correction: Review: A comprehensive summary of a decade development of the recombinase polymerase amplification. Analyst 2020, 145, 1950–1960. [Google Scholar] [CrossRef]
- Gong, L.; Jin, Z.; Liu, E.; Tang, F.; Yuan, F.; Liang, J.; Wang, Y.; Liu, X.; Wang, Y. Highly sensitive and specific detection of mobilized colistin resistance gene mcr-1 by CRISPR-based platform. Microbiol. Spectr. 2022, 10, e01884-22. [Google Scholar] [CrossRef]
- Tao, J.; Liu, D.; Xiong, J.; Dou, L.; Zhai, W.; Zhang, R.; Wang, Y.; Shen, J.; Wen, K. Rapid on-site detection of extensively drug-resistant genes in enterobacteriaceae via enhanced recombinase polymerase amplification and lateral flow biosensor. Microbiol. Spectr. 2022, 10, e03344-22. [Google Scholar] [CrossRef]
- Buderer, N.M.F. Statistical Methodology: I. Incorporating the prevalence of disease into the sample size calculation for sensitivity and specificity. Acad. Emerg. Med. 1996, 3, 895–900. [Google Scholar] [CrossRef]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Eurosurveillance 2018, 23, 17-00672. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotypic BMD | PCR | |||
|---|---|---|---|---|
| RPA-LFA | Colistin resistance | Colistin susceptible | MCR1+ | MCR1− |
| MCR1+ | 65 | 0 | 65 | 0 |
| MCR1− | 25 | 33 | 0 | 58 |
| Total | 90 | 33 | 65 | 58 |
| Sensitivity 72.2% CI 61.78% to 81.15% | Sensitivity 100.00% CI 94.48% to 100.00% | |||
| Specificity 100.00% CI 89.42% to 100.00% | Specificity 100.00% CI 93.84% to 100.00% | |||
| PPV 100.00% CI 94.48% to 100.00% | PPV 100.00% CI 94.48% to 100.00% | |||
| NPV 56.9% CI 48.61% to 64.81% | NPV 100.00% CI 93.84% to 100.00% | |||
| Accuracy 76.67% CI 71.48% to 86.39% | Accuracy 100.00% CI 97.0522% to 100.00% | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, N.; Suchanta, N.; Pimpitak, U.; Santanirand, P.; Am-In, N.; Chaichanawongsaroj, N. Validation of Recombinase Polymerase Amplification with In-House Lateral Flow Assay for mcr-1 Gene Detection of Colistin Resistant Escherichia coli Isolates. Antibiotics 2024, 13, 984. https://doi.org/10.3390/antibiotics13100984
Ullah N, Suchanta N, Pimpitak U, Santanirand P, Am-In N, Chaichanawongsaroj N. Validation of Recombinase Polymerase Amplification with In-House Lateral Flow Assay for mcr-1 Gene Detection of Colistin Resistant Escherichia coli Isolates. Antibiotics. 2024; 13(10):984. https://doi.org/10.3390/antibiotics13100984
Chicago/Turabian StyleUllah, Naeem, Nutchaba Suchanta, Umaporn Pimpitak, Pitak Santanirand, Nutthee Am-In, and Nuntaree Chaichanawongsaroj. 2024. "Validation of Recombinase Polymerase Amplification with In-House Lateral Flow Assay for mcr-1 Gene Detection of Colistin Resistant Escherichia coli Isolates" Antibiotics 13, no. 10: 984. https://doi.org/10.3390/antibiotics13100984
APA StyleUllah, N., Suchanta, N., Pimpitak, U., Santanirand, P., Am-In, N., & Chaichanawongsaroj, N. (2024). Validation of Recombinase Polymerase Amplification with In-House Lateral Flow Assay for mcr-1 Gene Detection of Colistin Resistant Escherichia coli Isolates. Antibiotics, 13(10), 984. https://doi.org/10.3390/antibiotics13100984