1. Introduction
Anti-microbial peptides (AMPs) are any polyamide, or even their bio-co-polymer with esters, thioesters, or otherwise modified backbone, that can be made on a contemporary chemical peptide synthesizer [
1]. This provides an option of taking advantage of using the remarkable toolkit of quantitative structure–activity relation (QSAR) [
2,
3] in designing novel anti-microbial-active sister molecules of any newly discovered natural drug-candidate AMP molecule. As for their biosynthesis, peptides of anti-microbial potential can be either ribosomal encoded (RP) single gene products [
4] or enzymatically biosynthesized non-ribosomal templated peptides (NRP) [
5], (NR-AMPs) [
6]. Considering that there are already >100 peptide-based drugs that have been of clinical use (see [
7]), an lookout at that field may be advisable. As for history, we refer to the review of Zasloff [
8], who outlined those discoveries that can be considered as milestones, like cecropins [
9], defensins [
10,
11], maganins [
12], and add the proline-rich AMP (PrAMP family) [
13,
14] to the still incomplete list. Many derivatives of discovered natural AMPs have been chemically synthesized and or modified, and there are several excellent reviews [
15,
16,
17,
18,
19] beyond the scope of this paper.
The two selected targets here are
Leishmania [
20], a mammalian pathogen, transmittable by insects like cave-dwelling sand flies [
21], and
Histomonas meleagridis, the etiological agent of the life-threatening histomonosis (syn. blackhead disease) of poultry species such as turkeys, chickens, and pheasants, first described in 1893 [
22,
23], and have been studied in detail until the mid of last century [
24].
In the actual study we aimed to investigate if any of those biosynthetic NR-AMPs, like fabclavine [
25,
26], phenazine [
27], and others, produced by entomopathogenic–nematode symbiont bacteria (EPB) we have been studying [
28,
29] were suitable as anti-protozoal drug candidates against the intracellular parasite
Leishmania or the extracellular parasite
H. meleagridis. The arguments for choosing in vitro EPB liquid cultures as appropriate sources of efficient AMPs for this purpose are that obligate bacterial symbionts of entomopathogenic–nematode species synthesize and release several non-ribosomal hybrid peptides [
30] with large target specificities. These mainly serve to provide well-balanced pathobiome conditions for this symbiosis in polyxenic soil and cadaver environments [
31]. These peptides are also considered potential sources of potent natural anti-microbial compounds [
30]. The motivation for choosing in vitro EPB liquid cultures as a source of efficient AMPs for this purpose is the proven activity on Gram-positive [
32,
33] and Gram-negative [
34] pathogenic bacteria, oomycetes, and different plant pathogens [
35,
36] in our previous studies. Since we have reasons to believe that some of the autoclaved
Xenorhabdus cultures may not be harmful or toxic when added as a food supplement, while some anti-microbial ingredients retain active [
6], we consider the potential for probiotic applications, at least on an experimental level.
The eukaryotic pathogens studied here include the
Leishmania donovanii species, kinetoplastid protozoa [
37], and the causative agent of human and canine leishmaniasis [
38,
39,
40].
Leishmania are intracellular parasites that target professional phagocytes (macrophages and dendritic cells) [
41] and human infection leads to diverse forms of disease from singular or diffuse cutaneous lesions (cutaneous leishmaniasis), invasive destruction of mucous membranes (mucocutaneous leishmaniasis), or dissemination to the liver, spleen, and bone marrow (visceral leishmaniasis) [
42,
43]. While mainly relegated to endemic foci in poor rural areas in the third world, the infections have a global impact due to human migration, climate change, and anthropogenic disturbance, causing significant worldwide health and economic burdens [
44]. There are currently no approved vaccines available [
45], leaving control of leishmaniasis reliant on chemotherapy [
46]. The mainstays of therapy are antimonial derivatives [
47] of amphotericin B (AmB), a polyene macrolide antibiotic derived from actinomycetes [
48], which can be renally toxic and are not widely available in poor rural areas. Increasingly, there are issues with leishmanial infections that are resistant or refractory to antimony and amphotericin therapy. Miltefosine is the first oral agent approved for the treatment of leishmaniasis; however, there have been descriptions of miltefosine resistance [
46]. There is an obvious need for the development of newer, less toxic agents to combat these infections.
The other chosen pathogen,
H. meleagridis, belongs to the Dientamoebidae, order
Tritrichomanidida. Infections in turkeys may cause nearly 100% mortality whereas outbreaks in chickens, peafowl, quail, and pheasants are more often marked by morbidity and subsequent recovery.
H. meleagridgridis is carried by the eggs of the cecal worm
Heterakis gallinarum, enabling them to survive for long periods in the soil as a source of infection [
22,
23,
24]. In the EU and USA, there are currently no drugs available for the treatment of blackhead disease [
22,
24]. (As for molecular taxonomy and identification, the 5.8S, ITS-1, and ITS-2 rRNA regions were first sequenced [
49]. Since then, the complete, annotated
Histomonas reference genome sequence became available [
50]).
Since the introduction of effective drugs into the market in the middle of the previous century, there was no practical need for further research. Following the recent ban of available drugs, research programs with new profiles were set up in various places focusing on different features of the parasite and the disease.
Consequently, the poultry industry works without approved prophylactics, therapeutics, or vaccines to combat histomonosis [
51].
Recently, numerous chemical and botanical compounds were also tested for their efficacy against
H. meleagridis, with varying outcomes. One of the explanations for these half-failures/half-successes may be the complicated in vitro
Histomonas culturing technique. In this context, it is of high importance that
H. meleagridis relies on live bacteria and
Escherichia coli strongly supports the growth of
H. meleagridis in a monoxenic culture without influencing its pathogenicity [
52]. In the previously mentioned study, a clonal cultures of
H. meleagridis was further optimized to obtain a monoxenic culture in a liquid medium [
52]. For this, the fecal flora was exchanged for defined bacterial strains by selective destruction of the initial bacteria with various antibiotics, keeping the flagellate alive.
E. coli was found to strongly support the growth of the parasite, whereas
Salmonella enterica serovar Typhimurium and
Pseudomonas aeruginosa were less efficient. Whether the special feature of the parasite’s intricate interplay with bacteria in vitro and in vivo can be considered mutualistic or a predator-prey one is an open question [
50,
51,
52,
53,
54,
55,
56,
57,
58,
59,
60]. (We are shared about it). The option of the monoxenic culturing technique may open a door to preliminary attempts to protect birds with cultured attenuated histomonads, forecasting the possibility of vaccination [
56].
After banning all previously used prophylactic and therapeutic drugs against
H. meleagridis in the USA and the European Union, benzimidazoles were tested but found inactive [
61]. Similar half-successes were reported about various herbal substances, some of which showed good efficacy in vitro but failed in vivo [
62]. Reduced sensitivity of
H. meleagridis to nitarsone in vitro and in vivo was also shown [
63].
A publication talked about that, identical monoxenic settings for cultures of the same
H. meleagridis clonal strain, in its virulent low-passage and attenuated high-passage form, enabled a comparative analysis of parasite characteristics [
64], but the conclusions severely contradict a previously cited, unambiguously reliable publication [
59]. This debate is out of scope of the present paper.
Herein, we compared the spectrum of in vitro anti-microbial/-prozozoal activity of cell-free conditioned culture media (CFCM) from three EPB species, together with testing against
Leishmania donovanii, (together with
Trypanosoma cruzi, as an expectedly negative control),
Histomonas meleagridis, and also against a panel of clinical bacterial and fungal isolates, as positive controls. Considering the roles of the bacterial symbiont in the EPN/EPB symbiosis, we suppose that ani-protozoal compounds must coexist with other anti-microbial compounds in the CFCM of EPBs. Therefore, for reliable bioassays, we should use appropriate controls. Two main aspects should be taken into consideration: (A) To be able to distinguish between the specified antiprotist activity and general cytotoxicity of the future drug-candidate anti-microbial compounds in the CFCM. (B) To be able to identify the special antiprotist compounds. In this paper, we describe the very first steps toward these goals. For (A), one has to consider that the lack of uniformity regarding the choice of cell types for cytotoxicity assays may lead to incomparable and inconclusive data. In vitro assays relying solely on non-phagocytic cell models may not represent a realistic result, as the effect of an anti-leishmanial agent should ideally be presented based on its cytotoxicity profile against reticuloendothelial system cells [
65]. In the
Leishmania studies, we used the macrophage cell line J774A.1 [
66,
67,
68], as a cytopathogenicity control test organism (see
Section 2 Materials and Methods). In the
Histomonas studies, we used the permanent chicken liver cells (LMH leghorn male hepatoma) cell line [
6,
69,
70] as a cytopathogenicity control test organism (see
Section 2 Materials and Methods). For (B), one has to take into consideration the following points: each NR-AMP-producing EPB cell is in the so-called primary (1
0) phase, or Phase 1 [
71]. In other terms, the phenotypic precondition for antibiotic production (as well as the capability of symbiosis) of the EPB species is the primary phase [
30,
72,
73]. For NR-AMP production, it means that in a liquid culture of the primary (1
0) cells (of each of the studied EPB species, strain, and isolate), more than one biosynthetic AMP is present simultaneously. Each expresses anti-microbial (maybe antiprotist) activity, so the anti-microbial (antiprotist) activity of a CFCM sample represents a cumulative activity. However, in an optimal test system, one would prefer to determine the anti-microbial activity of each of the single anti-microbial compounds one by one.
Since the discovery that the activity (the “switch-on”/“switch-off” states) of those group genes (operons, biosynthetic gene complexes, and BGCs), which are responsible for the primary-secondary phenotypic phase shift in each known EPB strain, not only work under the control of the versatile regulator hexA [
74,
75], but are also coordinately regulated at a higher level (via
hfq-gene controlled sRNS/HexA-mRNA base pairing). More precisely, the Hfq-dependent sRNA, ArcZ, directly base-pairs with the HexA-encoding mRNA [
76,
77,
78]; it is possible to construct double-mutant strains from each EPB species (or isolate) following the genuine strategy called easyPACId (easy promoter-activated compound identification) approach [
79]. Each biosynthesizes and releases only one single NR-AMP molecule into the culture media. This discovery may be a starting point of the renaissance of
Xenorhabdus anti-microbial peptide research [
76,
77,
78,
79,
80,
81,
82,
83]. We managed to reconstruct the “easyPACId”
hfq deletion mutant strains from EMA and EMC (Boros et al., in preparation) and publish here some information, including data about their anti-microbial potential. In this paper, we give an account of the details of constructing our
hfq-del mutants from our EMA and EMC [
29] strains and provide their phenotypic descriptions. Our “easyPACId” strains will be available for cooperation with fellow scientists worldwide.
4. Discussion
When planning experiments aimed at applying AMPs against eukaryotic (for instance, anti-protozoal) pathogens, we have to take into consideration that the strategy of designing antiprotozoals to control eukaryotic parasites (pathogens) must be different from that of designing antimicrobials to control prokaryotic pathogens [
97,
98,
99]. The targetable intracellular structures of a eukaryotic parasite (pathogen) biochemically might be similar to some intracellular structures of vital functions in the eukaryote host to be protected.
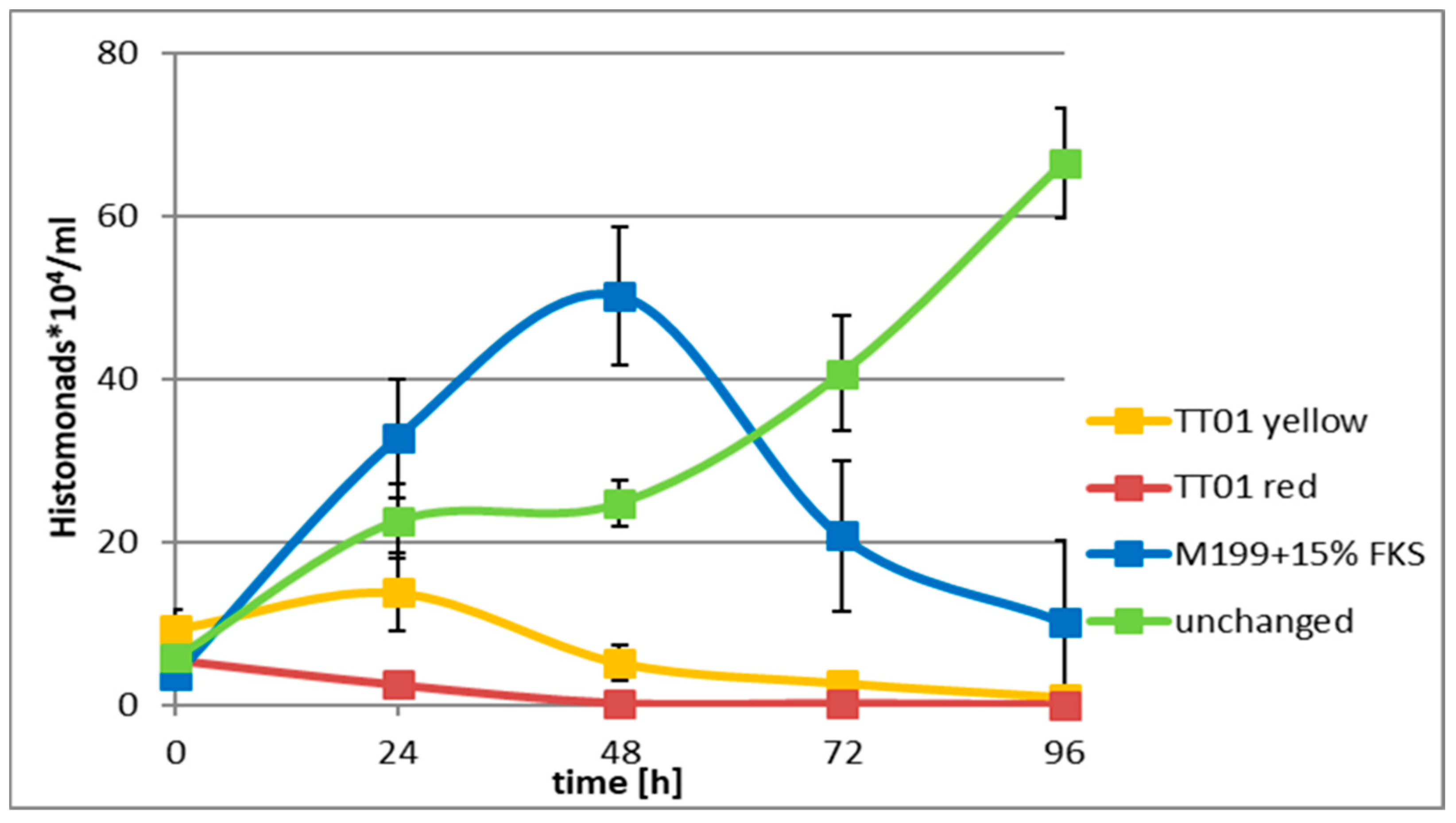
As for our H. meleagrididis exeriments, we worked in a monoxenic system where prokaryotic and eukaryotics cells coexisted. We found that both the prokaryotic and the eukaryotic cells died when EPB CFCM were added in th right concentration.
The question is, however, that what was the primary reason for the death of the H. meleagridis cells. They might die (1) because their procaryote mutualistic partner died; (2) they might be killed by a direct antiprotist activity of any (or more) AMP molecules of the EPB CFCMs; or (3) the two killing effects cumulated. Unfortunately, these published experiments cannot give an unambiguous answer to that question. We hope for the answer from our future experiments when we intend to use CFCMs from double (easyPACId) mutants (with hfq-deletion and exchanged (re-activated) promoter of only one AMP-responsible operon. We will search for some which is harmless to prokaryotic partner but kill H. meleagridis cells selectively). It is true, that the peptides killed the mutualistic (E. coli DH5@) bacteria, which itself could be considered as an explanation of the death of the H. meleagridis cells. It is also true, that from our experiments no proof supports the idea that histomonads were killed directly. But we also have evidence that CFCM contains active anti-protist active components as well, (see the Leishmania donovanii data from this study or the published results of the Bode group and others), so the only question is what of these compounds is capable of (in lucky case, selectively) killing H. meleagridis. This is what we intend to learn from our future easyPACId mutant experiments.
This paper is about the anti-protist potential of anti-microbial compounds produced by entomopathogenic bacterium species (EMA, EMC, and EMK
Photorhabdus luminescens TT01) [
6,
28,
29,
93], the obligate symbionts of certain entomopathogenic–nematodes (EPN). In laboratory in vitro conditions they release their products into their conditioned culture media (CFCM) in vitro. The chemical nature of these antimicrobials is oligo-peptide. However they are non-ribosomal anti-microbial peptides (NR-AMPs), which means that they are enzymatically synthesized through more than one step in their respective biosynthetic pathways. Those genes encoding for the enzymes forming the respective biosynthetic pathway of given NR-AMP are clustered in a single operon [
100], also called biosynthetic gene complex (BGC) [
101]. One of the natural roles of the prokaryote partner in the EPN/EPB symbiosis is to protect the given monoxenic symbiotic association in a given polyxenic milieu, producing a chemical toolkit (e.g., a particular set of NR-AMPs) providing competitiveness against the prokaryotic and eukaryotic competitors, including protists. The biosynthesis of the given sets of these effective antimicrobials is synchronized by a unique, genuine hierarchical genetic regulation mechanism called primary-secondary phase shift [
71] with the
hfq gene on the top [
74,
75,
76,
77,
78]. In the laboratory liquid culture conditions (as well as in the active symbiotic stage), these compounds are produced abundantly, providing an inexhaustible “gold mine” for the anti-microbial searching scientist. The bioassays of the CFCMs are the tools to find the best sources and optimize the culture conditions. We found that those strains (EMA and EMC) which were discovered and further developed not only by our international team of a “laboratory without walls” in those famous laboratories of Maxime Gualtieri in France and Helge Bode in Germany, proving that these two strains are probably best found so far. In addition, other laboratories are probably really excellent sources. The data presented here on anti-leishmanial and anti-histomonal potentials seem to confirm this conclusion. The next step is to find a way to obtain mutants, not “genetically manipulated”, but mutant strains which produce only single NR-AMP molecules instead of a bunch of them. The method to do that was recently discovered in the Bode laboratory [
79,
80,
81,
82,
83]. We demonstrate here that we have started this project as well by constructing and characterizing
hfq-del mutants from both EPB species EMA and EMC.
Our larger scope is to benefit from the existence of NR-AMP amongst the metabolites of our EPB bacteria. In other terms, our goals are to beat target pathogens, including prokaryotic pathogenic and zoonic bacteria, and eukaryotic parasites and pathogens of different taxa, which are important from the aspects of importance for animal and human health.
We are keenly interested in exploring whether the anti-microbial activity of NR-AMP of EMA and EMC can be developed as chemotherapeutic agents to treat protozoa such as the intracellular parasite
Leishmania [
37] and the extracellular avian pathogen
H. gallinarum [
97]. Both of these flagellated protozoa have been demonstrated to have unique and efficient resistance mechanisms to other anti-parasitic agents and require the development of additional effective agents.
Leishmania spp. is particularly problematic in light of its sophisticated resistance mechanisms, but it is also an excellent model for analyzing the mechanisms of drug resistance in eukaryotes [
98,
99]. Our research conception is that we think that some
Xenorhabdus CFCM molecules (i.e., phenazine, fabclavine, or other molecules) may be selectively capable of dysregulating eukaryotic cell functions and stimulating apoptosis in protozoa without affecting the host [
100]. From an application point of view, the development of natural compounds as anti-protozoal requires several aspects to be taken into consideration: (A) the anti-protozoal potential; (B) the durability, thermotolerance, and bioavailability; and (C) cytotoxicity, and unwanted side effects, especially if target organisms are eukaryotic pathogens. We have demonstrated that
Xenorhabdus CFCM has broad-spectrum anti-protozoal activity against several human microbial pathogens. The key issue is identifying the specific active molecules in a complex mixture of potential anti-microbial molecules. The CFCMs of all three
Xenorhabdus species were all active against
Leishmania promastigotes in our experiments, but interestingly, none were active against
Trypanosoma cruzi epimastigotes. Other researchers [
83] found that EMA and EMK CFCMs were equally efficient against both
Leishmania and
Trypanosoma. In that study, they found that
X. budapestensis,
X. cabanillasii,
X. hominickii,
X. indica,
X. innexii, and
X. stockiae supernatants caused 100% mortality at the highest tested concentration (10%) against the promastigote form of
L. tropica. No differences occurred between this treatment group and the positive control (
p > 0.05) using N-methyl meglumine. There are several possible explanations for the differences between our findings and those of the Turkish team. For instance, the two studies used different strains of
T. cruzi. In addition, there are differences in the experimental methods used between the two studies. In the case of the differential susceptibility of
L. amazoennsis and
T. cruzi (Brazil strain), it is likely due to differences in the surface compositions between the two parasites that lead to differential disruption of the cell membrane and/or intracellular penetration to act on specific subcellular targets. We focus on EMA and EMC species. We compared the anti-microbial potential of each of the used
hfq-del mutants before using them as the source of double mutants producing single NR-AMPs. Our data in
Table 3,
Table 4 and
Table 5 show that the anti-microbial activities of the
hfq-del mutants can have enhanced anti-microbial activity not present in the wild-type stains. This may not be quite surprising considering that approximately 7.5% of the total genes in
Xenorhabdus bacteria are dedicated to secondary metabolite biosynthesis, and probably most of these products are NR-AMPs. With an average 2.4 Mb-genome size of
Xenorhabdus and a 1 kb locus size with about 10 ORFs per operon, there are 2400 genes and 240 respective operons (BGS) per genome. It is comparable with that of
Staphylococcus, which has about 225 operons. According to the data accumulated, only about a dozen of biosynthetic anti-microbials have been discovered. In our lab, we intend to work with about 12 operons, responsible for the biosynthesis of 12 NR-AMPs in EMC, and operons which are responsible for 3 NR_AMPs in EMA under
hfq regulations.
However, we cannot rule out that there are several other operons encoding for the synthesis of additional anti-microbial compounds, which are not under hfq control. In the future, we intend to establish and analyze RNA-seq from our wild-type stains and their
hfq mutants. We hypothesize that the situation is somewhat simpler since the hfq likely regulates a well-defined group of operons providing the genetic machinery for the so-called “primary-secondary phase shift” [
72] (or “phenotypic phase variation”, the term of Professor S. A. Forst), which describes that only a relatively small set of natural products (NP) are anti-microbial synthetic genes. Toxicity studies using J774 macrophages [
66,
67] have indicated that only the CFCM of EMA was able to cause significant mortality of these cells. In contrast, the CFCMs of EMC or EMK had no effect. This experiment confirmed our previous unpublished observations that EMA exerts stronger cytotoxicity on a range of eukaryotic cells, which may cause limitations concerning future medical applications. However, this selective cytotoxicity may provide an option to select for and find specific CFCMs against different eukaryotic parasites of different animal and plant hosts. Interestingly, our studies with J774 cells and the CFCMs of EMA, EMC, and EMK did not show toxicity. These differences may be related to different experimental methods or differences in the bacterial and cell types used.
Overall, in light of the availability of few agents for the treatment of leishmaniasis and the increasing incidence of development resistance, there is a demand for new agents [
100,
101,
102,
103]. Moving forward, drug discovery will be based on complete genome sequencing of multiple strains, utilization of CRISPR/Cas9 technology for gene inactivation, in vivo, and bioluminescence-based imaging together with high-throughput analysis of compounds [
44]. As for the results in the control (bacterium, fungi, and other strains), previous studies have identified several anti-microbial molecules in the CFCM of
Xenorhabdus, including fabclavine [
25,
26]. This strange metabolite polypeptide/polyketide hybrid molecule is synthesized via non-ribosomal peptide synthesis. Many different-sized derivatives of fabclavines can be produced by
X. szentirmaii [
80] and can vary between entomopathogenic–nematode–symbiont (
Xenorhabdus,
Photorhabdus) bacterium species [
80] (Wenski et al., 2019). These have a wide-ranging anti-microbial activity against various microbes [
29]. Several other anti-microbial molecules are known to be produced by different EPB species and released into their CFCMs. Xenofuranone A and B [
104], cabanillasin [
105], PAX peptides [
106], odilorhabdins [
107], cyclic depsipeptides (xenematides, F and G), anti-oomycete peptides [
108], novel anti-microbial peptides [
109], xenortide [
110], xenortides, rhabdopeptided/xenortide-like peptides [
111], other rhabdopeptides [
112], and szentiamid [
113,
114] have been discovered in the last ten years in different
Xenorhabdus species.The proper approach is the easyPACId method, based on the latest information on the highest organization level of gene regulation in bacteria [
115,
116,
117,
118,
119,
120]. Whether any of them can be considered a potential anti-histomonal drug candidate will be learned if we had a chance to screen both the single mutant
hfq-del stains of EMA and EMC.



 ,
, 




{kind=link}
{kind=link}

