

Drivers of Antibiotic Resistance Gene Abundance in an Urban River


Abstract
1. Introduction
2. Results and Discussion
3. Conclusions
4. Materials and Methods
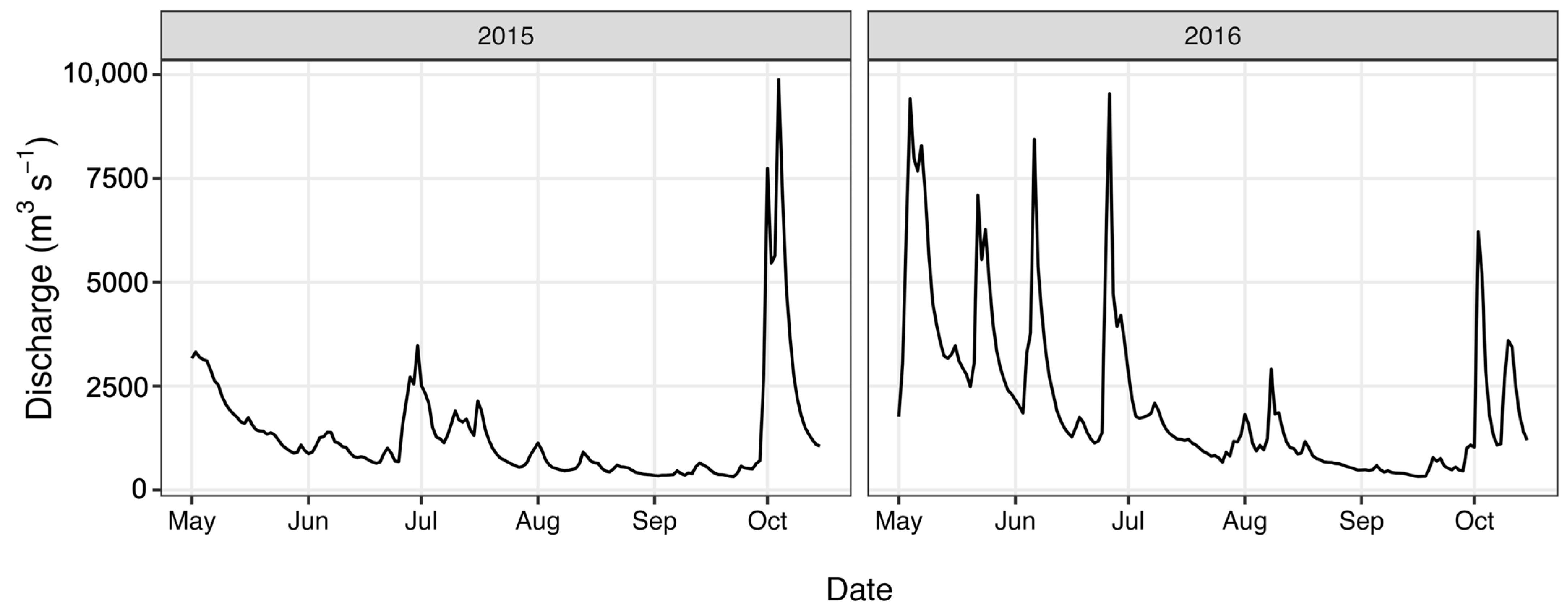
4.1. Site Description
4.2. Water Sampling
4.3. Environmental Data
4.4. DNA Extractions
4.5. Quantitative Polymerase Chain Reaction (qPCR)
4.6. Data Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Z.; Zhang, Q.; Wang, T.; Xu, N.; Lu, T.; Hong, W.; Penuelas, J.; Gillings, M.; Wang, M.; Gao, W.; et al. Assessment of Global Health Risk of Antibiotic Resistance Genes. Nat. Commun. 2022, 13, 1553. [Google Scholar] [CrossRef]
- Koch, N.; Islam, N.F.; Sonowal, S.; Prasad, R.; Sarma, H. Environmental Antibiotics and Resistance Genes as Emerging Contaminants: Methods of Detection and Bioremediation. Curr. Res. Microb. Sci. 2021, 2, 100027. [Google Scholar] [CrossRef]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Proia, L.; Anzil, A.; Subirats, J.; Borrego, C.; Farrè, M.; Llorca, M.; Balcázar, J.L.; Servais, P. Antibiotic Resistance along an Urban River Impacted by Treated Wastewaters. Sci. Total Environ. 2018, 628–629, 453–466. [Google Scholar] [CrossRef]
- Thakali, O.; Malla, B.; Tandukar, S.; Sthapit, N.; Raya, S.; Furukawa, T.; Sei, K.; Sherchand, J.B.; Haramoto, E. Release of Antibiotic-Resistance Genes from Hospitals and a Wastewater Treatment Plant in the Kathmandu Valley, Nepal. Water 2021, 13, 2733. [Google Scholar] [CrossRef]
- Thai, P.K.; Ky, L.X.; Binh, V.N.; Nhung, P.H.; Nhan, P.T.; Hieu, N.Q.; Dang, N.T.T.; Tam, N.K.B.; Anh, N.T.K. Occurrence of Antibiotic Residues and Antibiotic-Resistant Bacteria in Effluents of Pharmaceutical Manufacturers and Other Sources around Hanoi, Vietnam. Sci. Total Environ. 2018, 645, 393–400. [Google Scholar] [CrossRef]
- Kotwani, A.; Joshi, J.; Kaloni, D. Pharmaceutical Effluent: A Critical Link in the Interconnected Ecosystem Promoting Antimicrobial Resistance. Environ. Sci. Pollut. Res. 2021, 28, 32111–32124. [Google Scholar] [CrossRef]
- He, Y.; Yuan, Q.; Mathieu, J.; Stadler, L.; Senehi, N.; Sun, R.; Alvarez, P.J.J. Antibiotic Resistance Genes from Livestock Waste: Occurrence, Dissemination, and Treatment. Npj Clean Water 2020, 3, 4. [Google Scholar] [CrossRef]
- Reddy, S.; Kaur, K.; Barathe, P.; Shriram, V.; Govarthanan, M.; Kumar, V. Antimicrobial Resistance in Urban River Ecosystems. Microbiol. Res. 2022, 263, 127135. [Google Scholar] [CrossRef]
- Sowah, R.A.; Molina, M.; Georgacopoulos, O.; Snyder, B.; Cyterski, M. Sources and Drivers of ARGs in Urban Streams in Atlanta, Georgia, USA. Microorganisms 2022, 10, 1804. [Google Scholar] [CrossRef]
- Marti, E.; Variatza, E.; Balcazar, J.L. The Role of Aquatic Ecosystems as Reservoirs of Antibiotic Resistance. Trends Microbiol. 2014, 22, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Balasa, G.; Levengood, E.S.; Battistelli, J.M.; Franklin, R.B. Diversity of Multidrug-Resistant Bacteria in an Urbanized River: A Case Study of the Potential Risks from Combined Sewage Overflows. Water 2021, 13, 2122. [Google Scholar] [CrossRef]
- Matsui, K.; Miki, T. Microbial Community Composition and Function in an Urban Waterway with Combined Sewer Overflows before and after Implementation of a Stormwater Storage Pipe. PeerJ 2023, 11, e14684. [Google Scholar] [CrossRef] [PubMed]
- Honda, R.; Tachi, C.; Yasuda, K.; Hirata, T.; Noguchi, M.; Hara-Yamamura, H.; Yamamoto-Ikemoto, R.; Watanabe, T. Estimated Discharge of Antibiotic-Resistant Bacteria from Combined Sewer Overflows of Urban Sewage System. Npj Clean Water 2020, 3, 15. [Google Scholar] [CrossRef]
- Collignon, P.; Beggs, J.J.; Walsh, T.R.; Gandra, S.; Laxminarayan, R. Anthropological and Socioeconomic Factors Contributing to Global Antimicrobial Resistance: A Univariate and Multivariable Analysis. Lancet Planet. Health 2018, 2, e398–e405. [Google Scholar] [CrossRef]
- Di Cesare, A.; Eckert, E.M.; Rogora, M.; Corno, G. Rainfall Increases the Abundance of Antibiotic Resistance Genes within a Riverine Microbial Community. Environ. Pollut. 2017, 226, 473–478. [Google Scholar] [CrossRef]
- Ouyang, W.-Y.; Huang, F.-Y.; Zhao, Y.; Li, H.; Su, J.-Q. Increased Levels of Antibiotic Resistance in Urban Stream of Jiulongjiang River, China. Appl. Microbiol. Biotechnol. 2015, 99, 5697–5707. [Google Scholar] [CrossRef]
- Xu, Y.; Guo, C.; Luo, Y.; Lv, J.; Zhang, Y.; Lin, H.; Wang, L.; Xu, J. Occurrence and Distribution of Antibiotics, Antibiotic Resistance Genes in the Urban Rivers in Beijing, China. Environ. Pollut. 2016, 213, 833–840. [Google Scholar] [CrossRef]
- Zhou, Z.-C.; Zheng, J.; Wei, Y.-Y.; Chen, T.; Dahlgren, R.A.; Shang, X.; Chen, H. Antibiotic Resistance Genes in an Urban River as Impacted by Bacterial Community and Physicochemical Parameters. Environ. Sci. Pollut. Res. 2017, 24, 23753–23762. [Google Scholar] [CrossRef]
- Pan, X.; Chen, L.; Zhang, L.; Zuo, J. Characteristics of Antibiotic Resistance Gene Distribution in Rainfall Runoff and Combined Sewer Overflow. Environ. Sci. Pollut. Res. 2022, 30, 30766–30778. [Google Scholar] [CrossRef]
- Petrie, B. A Review of Combined Sewer Overflows as a Source of Wastewater-Derived Emerging Contaminants in the Environment and Their Management. Environ. Sci. Pollut. Res. 2021, 28, 32095–32110. [Google Scholar] [CrossRef] [PubMed]
- United States Environmental Protection Agency. EPA Report to Congress on Impacts and Control of Combined Sewer Overflows and Sanitary Sewer Overflows; United States Environmental Protection Agency: Washington, DC, USA, 2004.
- Abdellatif, M.; Atherton, W.; Alkhaddar, R.M.; Osman, Y.Z. Quantitative Assessment of Sewer Overflow Performance with Climate Change in Northwest England. Hydrol. Sci. J. 2015, 60, 636–650. [Google Scholar] [CrossRef]
- Li, J.; Li, X.; Liu, H.; Gao, L.; Wang, W.; Wang, Z.; Zhou, T.; Wang, Q. Climate Change Impacts on Wastewater Infrastructure: A Systematic Review and Typological Adaptation Strategy. Water Res. 2023, 120282. [Google Scholar] [CrossRef] [PubMed]
- Karkman, A.; Pärnänen, K.; Larsson, D.G.J. Fecal Pollution Can Explain Antibiotic Resistance Gene Abundances in Anthropogenically Impacted Environments. Nat. Commun. 2019, 10, 80. [Google Scholar] [CrossRef]
- Zhang, X.; Zhi, X.; Chen, L.; Shen, Z. Spatiotemporal Variability and Key Influencing Factors of River Fecal Coliform within a Typical Complex Watershed. Water Res. 2020, 178, 115835. [Google Scholar] [CrossRef]
- Vitro, K.A.; BenDor, T.K.; Jordanova, T.V.; Miles, B. A Geospatial Analysis of Land Use and Stormwater Management on Fecal Coliform Contamination in North Carolina Streams. Sci. Total Environ. 2017, 603–604, 709–727. [Google Scholar] [CrossRef] [PubMed]
- Weyrauch, P.; Matzinger, A.; Pawlowsky-Reusing, E.; Plume, S.; Von Seggern, D.; Heinzmann, B.; Schroeder, K.; Rouault, P. Contribution of Combined Sewer Overflows to Trace Contaminant Loads in Urban Streams. Water Res. 2010, 44, 4451–4462. [Google Scholar] [CrossRef]
- Wang, J. Combined Sewer Overflows (CSOs) Impact on Water Quality and Environmental Ecosystem in the Harlem River. JEP 2014, 5, 1373–1389. [Google Scholar] [CrossRef]
- Diaz-Fierros, T.F.; Puerta, J.; Suarez, J.; Diaz-Fierros, V.F. Contaminant Loads of CSOs at the Wastewater Treatment Plant of a City in NW Spain. Urban Water 2002, 4, 291–299. [Google Scholar] [CrossRef]
- Damashek, J.; Westrich, J.R.; McDonald, J.M.B.; Teachey, M.E.; Jackson, C.R.; Frye, J.G.; Lipp, E.K.; Capps, K.A.; Ottesen, E.A. Non-Point Source Fecal Contamination from Aging Wastewater Infrastructure Is a Primary Driver of Antibiotic Resistance in Surface Waters. Water Res. 2022, 222, 118853. [Google Scholar] [CrossRef]
- Zhang, X.-X.; Zhang, T.; Fang, H.H.P. Antibiotic Resistance Genes in Water Environment. Appl. Microbiol. Biotechnol. 2009, 82, 397–414. [Google Scholar] [CrossRef]
- Patterson, A.J.; Rincon, M.T.; Flint, H.J.; Scott, K.P. Mosaic Tetracycline Resistance Genes Are Widespread in Human and Animal Fecal Samples. Antimicrob. Agents Chemother. 2007, 51, 1115–1118. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Auerbach, E.A.; Seyfried, E.E.; McMahon, K.D. Tetracycline Resistance Genes in Activated Sludge Wastewater Treatment Plants. Water Res. 2007, 41, 1143–1151. [Google Scholar] [CrossRef]
- Pruden, A.; Larsson, D.G.J.; Amézquita, A.; Collignon, P.; Brandt, K.K.; Graham, D.W.; Lazorchak, J.M.; Suzuki, S.; Silley, P.; Snape, J.R.; et al. Management Options for Reducing the Release of Antibiotics and Antibiotic Resistance Genes to the Environment. Environ. Health Perspect. 2013, 121, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Laird, E.; Gentry, T.J.; Brooks, J.P.; Karthikeyan, R. Increased Antimicrobial and Multidrug Resistance Downstream of Wastewater Treatment Plants in an Urban Watershed. Front. Microbiol. 2021, 12, 657353. [Google Scholar] [CrossRef] [PubMed]
- Keely, S.P.; Brinkman, N.E.; Wheaton, E.A.; Jahne, M.A.; Siefring, S.D.; Varma, M.; Hill, R.A.; Leibowitz, S.G.; Martin, R.W.; Garland, J.L.; et al. Geospatial Patterns of Antimicrobial Resistance Genes in the US EPA National Rivers and Streams Assessment Survey. Environ. Sci. Technol. 2022, 56, 14960–14971. [Google Scholar] [CrossRef]
- Thakali, O.; Tandukar, S.; Brooks, J.; Sherchan, S.; Sherchand, J.; Haramoto, E. The Occurrence of Antibiotic Resistance Genes in an Urban River in Nepal. Water 2020, 12, 450. [Google Scholar] [CrossRef]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.A.; Dantas, G. The Shared Antibiotic Resistome of Soil Bacteria and Human Pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef]
- Wang, S.; Gao, X.; Gao, Y.; Li, Y.; Cao, M.; Xi, Z.; Zhao, L.; Feng, Z. Tetracycline Resistance Genes Identified from Distinct Soil Environments in China by Functional Metagenomics. Front. Microbiol. 2017, 8, 1406. [Google Scholar] [CrossRef]
- Daghrir, R.; Drogui, P. Tetracycline Antibiotics in the Environment: A Review. Environ. Chem. Lett. 2013, 11, 209–227. [Google Scholar] [CrossRef]
- Barr, B.S.; Waldridge, B.M.; Morresey, P.R.; Reed, S.M.; Clark, C.; Belgrave, R.; Donecker, J.M.; Weigel, D.J. Antimicrobial-Associated Diarrhoea in Three Equine Referral Practices: Antimicrobial-Associated Diarrhoea in Three Equine Referral Practices. Equine Vet. J. 2013, 45, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Rule, E.K.; Boyle, A.G.; Redding, L.E. Antimicrobial Prescribing Patterns in Equine Ambulatory Practice. Prev. Vet. Med. 2021, 193, 105411. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, J.; Zhu, Y.; Chen, Q.; Shen, F.; Wu, Y.; Xu, S.; Fan, H.; Da, G.; Huang, R.; et al. Global Survey of Antibiotic Resistance Genes in Air. Environ. Sci. Technol. 2018, 52, 10975–10984. [Google Scholar] [CrossRef]
- Bajaj, P.; Singh, N.S.; Kanaujia, P.K.; Virdi, J.S. Distribution and Molecular Characterization of Genes Encoding CTX-M and AmpC β-Lactamases in Escherichia Coli Isolated from an Indian Urban Aquatic Environment. Sci. Total Environ. 2015, 505, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Lachmayr, K.L.; Kerkhof, L.J.; DiRienzo, A.G.; Cavanaugh, C.M.; Ford, T.E. Quantifying Nonspecific TEM β-Lactamase (BlaTEM) Genes in a Wastewater Stream. Appl. Environ. Microbiol. 2009, 75, 203–211. [Google Scholar] [CrossRef]
- Coertze, R.D.; Bezuidenhout, C.C. Detection and Quantification of Clinically Relevant Plasmid-Mediated AmpC Beta-Lactamase Genes in Aquatic Systems. Water Supply 2020, 20, 1745–1756. [Google Scholar] [CrossRef]
- Coertze, R.D.; Bezuidenhout, C.C. Relating the Prevalence of Plasmid-Mediated AmpC Beta-Lactamase Genes to Aquatic Environmental Factors. Sci. Total Environ. 2021, 763, 144119. [Google Scholar] [CrossRef]
- Martínez, J.L.; Coque, T.M.; Baquero, F. What Is a Resistance Gene? Ranking Risk in Resistomes. Nat. Rev. Microbiol. 2015, 13, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Coertze, R.D.; Bezuidenhout, C.C. Global Distribution and Current Research of AmpC Beta-Lactamase Genes in Aquatic Environments: A Systematic Review. Environ. Pollut. 2019, 252, 1633–1642. [Google Scholar] [CrossRef]
- Song, M.; Song, D.; Jiang, L.; Zhang, D.; Sun, Y.; Chen, G.; Xu, H.; Mei, W.; Li, Y.; Luo, C.; et al. Large-Scale Biogeographical Patterns of Antibiotic Resistome in the Forest Soils across China. J. Hazard. Mater. 2021, 403, 123990. [Google Scholar] [CrossRef]
- Scott, L.C.; Lee, N.; Aw, T.G. Antibiotic Resistance in Minimally Human-Impacted Environments. Int. J. Environ. Res. Public Health 2020, 17, 3939. [Google Scholar] [CrossRef] [PubMed]
- Adachi, F.; Yamamoto, A.; Takakura, K.-I.; Kawahara, R. Occurrence of Fluoroquinolones and Fluoroquinolone-Resistance Genes in the Aquatic Environment. Sci. Total Environ. 2013, 444, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Golovko, O.; Kumar, V.; Fedorova, G.; Randak, T.; Grabic, R. Seasonal Changes in Antibiotics, Antidepressants/Psychiatric Drugs, Antihistamines and Lipid Regulators in a Wastewater Treatment Plant. Chemosphere 2014, 111, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Shi, Y.; Li, W.; Niu, H.; Liu, J.; Cai, Y. Occurrence of Antibiotics in Eight Sewage Treatment Plants in Beijing, China. Chemosphere 2012, 86, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Janecko, N.; Pokludova, L.; Blahova, J.; Svobodova, Z.; Literak, I. Implications of Fluoroquinolone Contamination for the Aquatic Environment-A Review: Fluoroquinolone in the Aquatic Ecosystem-A Review. Environ. Toxicol. Chem. 2016, 35, 2647–2656. [Google Scholar] [CrossRef]
- Virginia Geographic Information Network. 2016 Download: Land Cover Dataset Download Application. Available online: https://www.arcgis.com/home/item.html?id=d3d51bb5431a4d26a313f586c7c2c848 (accessed on 1 July 2023).
- Maddox, T.W.; Clegg, P.D.; Williams, N.J.; Pinchbeck, G.L. Antimicrobial Resistance in Bacteria from Horses: Epidemiology of Antimicrobial Resistance. Equine Vet. J. 2015, 47, 756–765. [Google Scholar] [CrossRef]
- Dolejska, M.; Duskova, E.; Rybarikova, J.; Janoszowska, D.; Roubalova, E.; Dibdakova, K.; Maceckova, G.; Kohoutova, L.; Literak, I.; Smola, J.; et al. Plasmids Carrying BlaCTX-M-1 and Qnr Genes in Escherichia Coli Isolates from an Equine Clinic and a Horseback Riding Centre. J. Antimicrob. Chemother. 2011, 66, 757–764. [Google Scholar] [CrossRef]
- Lupo, A.; Haenni, M.; Saras, E.; Gradin, J.; Madec, J.-Y.; Börjesson, S. Is Bla CTX-M-1 Riding the Same Plasmid Among Horses in Sweden and France? Microb. Drug Resist. 2018, 24, 1580–1586. [Google Scholar] [CrossRef]
- Schmiedel, J.; Falgenhauer, L.; Domann, E.; Bauerfeind, R.; Prenger-Berninghoff, E.; Imirzalioglu, C.; Chakraborty, T. Multiresistant Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae from Humans, Companion Animals and Horses in Central Hesse, Germany. BMC Microbiol. 2014, 14, 187. [Google Scholar] [CrossRef]
- Li, J.; Wang, T.; Shao, B.; Shen, J.; Wang, S.; Wu, Y. Plasmid-Mediated Quinolone Resistance Genes and Antibiotic Residues in Wastewater and Soil Adjacent to Swine Feedlots: Potential Transfer to Agricultural Lands. Environ. Health Perspect. 2012, 120, 1144–1149. [Google Scholar] [CrossRef]
- FDA. Antimicrobials Sold or Distributed for Use in Food-Producing Animals; U.S. Food & Drug Administration: Center for Veterinary Medicine: Silver Spring, MD, USA, 2021.
- Stanton, I.C.; Murray, A.K.; Zhang, L.; Snape, J.; Gaze, W.H. Evolution of Antibiotic Resistance at Low Antibiotic Concentrations Including Selection below the Minimal Selective Concentration. Commun. Biol. 2020, 3, 467. [Google Scholar] [CrossRef] [PubMed]
- Turiel, E.; Martín-Esteban, A.; Bordin, G.; Rodríguez, A.R. Stability of Fluoroquinolone Antibiotics in River Water Samples and in Octadecyl Silica Solid-Phase Extraction Cartridges. Anal. Bioanal. Chem. 2004, 380. [Google Scholar] [CrossRef]
- EPA. Method 1603: Escherichia Coli (E. Coli) in Water by Membrane Filtration Using Modified Membrane-Thermotolerant Escherichia Coli Agar (Modified MTEC); United States Environmental Protection Agency: Washington, DC, USA, 2009.
- Bukaveckas, P.A.; Barry, L.E.; Beckwith, M.J.; David, V.; Lederer, B. Factors Determining the Location of the Chlorophyll Maximum and the Fate of Algal Production within the Tidal Freshwater James River. Estuaries Coasts 2011, 34, 569–582. [Google Scholar] [CrossRef]
- Morina, J.C.; Morrissey, E.M.; Franklin, R.B. Vegetation Effects on Bacteria and Denitrifier Abundance in the Soils of Two Tidal Freshwater Wetlands in Virginia. Appl. Environ. Soil. Sci. 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- CDC. Outpatient Antibiotic Prescriptions—United States, 2015; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2015. [Google Scholar]
- Thames, C.H.; Pruden, A.; James, R.E.; Ray, P.P.; Knowlton, K.F. Excretion of Antibiotic Resistance Genes by Dairy Calves Fed Milk Replacers with Varying Doses of Antibiotics. Front. Microbio. 2012, 3, 139. [Google Scholar] [CrossRef]
- Aminov, R.I.; Garrigues-Jeanjean, N.; Mackie, R.I. Molecular Ecology of Tetracycline Resistance: Development and Validation of Primers for Detection of Tetracycline Resistance Genes Encoding Ribosomal Protection Proteins. Appl. Environ. Microbiol. 2001, 67, 22–32. [Google Scholar] [CrossRef]
- Marti, E.; Variatza, E.; Balcázar, J.L. Bacteriophages as a Reservoir of Extended-Spectrum β -Lactamase and Fluoroquinolone Resistance Genes in the Environment. Clin. Microbiol. Infect. 2014, 20, O456–O459. [Google Scholar] [CrossRef]
- Shi, P.; Jia, S.; Zhang, X.-X.; Zhang, T.; Cheng, S.; Li, A. Metagenomic Insights into Chlorination Effects on Microbial Antibiotic Resistance in Drinking Water. Water Res. 2013, 47, 111–120. [Google Scholar] [CrossRef]
- Marti, E.; Balcázar, J.L. Real-Time PCR Assays for Quantification of Qnr Genes in Environmental Water Samples and Chicken Feces. Appl. Environ. Microbiol. 2013, 79, 1743–1745. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 3 March 2023).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| tetO | tetW | blaTEM | qnrA | ampC | |
|---|---|---|---|---|---|
| Fecal Contamination Indicator | |||||
| E. coli abundance | 0.85 ** | 0.58 ** | 0.48 ** | 0.15 | 0.23 |
| Sewage Overflow Indicators | |||||
| Δ TN | 0.60 * | 0.11 | −0.11 | −0.11 | −0.28 |
| Δ TP | 0.67 ** | 0.16 | 0.17 | 0.20 | 0.04 |
| Discharge and Precipitation | |||||
| Discharge | −0.07 | −0.10 | 0.19 | 0.09 | −0.16 |
| Sampling-day precipitation | 0.37 | 0.46 * | 0.38 | 0.29 | 0.05 |
| Prior day-precipitation | 0.41 | 0.49 * | 0.46 * | 0.43 * | 0.04 |
| Bacterial Abundance | −0.10 | 0.10 | 0.18 | −0.12 | 0.58 ** |
| Antibiotic Class | Gene | Primer Information | Thermal Conditions (°C) | ||
|---|---|---|---|---|---|
| Names | Source | µM | |||
| Tetracyclines | tetW | tetW-F; tetW-R | [70] | 0.10 | 95° for 4 min, 40 cycles of 95° for 30 s, 57.4° for 15 s, 72° for 15 s |
| tetO | tetO-FW; tetO-RW | [71] | 0.20 | 94° for 5 min, 45 cycles of 94° for 30 s, 60° for 30 s, 72° for 30 s | |
| β-lactams | blaTEM | blaTEM-FX; blaTEM-RX | [72] | 0.30 | 95° for 5 min, 40 cycles of 95° for 15 s, 61° for 30 s, 72° for 30 s |
| ampC | ampC-F; ampC-R | [73] | 0.20 | 94° for 3 min, 40 cycles of 94° for 20 s, 58° for 20 s, 72° for 45 s | |
| Quinolones | qnrA | qnrAf-RT; qnrAr-RT | [74] | 0.10 | 95° for 3 min, 45 cycles of 95° for 15 s, 59.9° for 20 s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morina, J.C.; Franklin, R.B. Drivers of Antibiotic Resistance Gene Abundance in an Urban River. Antibiotics 2023, 12, 1270. https://doi.org/10.3390/antibiotics12081270
Morina JC, Franklin RB. Drivers of Antibiotic Resistance Gene Abundance in an Urban River. Antibiotics. 2023; 12(8):1270. https://doi.org/10.3390/antibiotics12081270
Chicago/Turabian StyleMorina, Joseph C., and Rima B. Franklin. 2023. "Drivers of Antibiotic Resistance Gene Abundance in an Urban River" Antibiotics 12, no. 8: 1270. https://doi.org/10.3390/antibiotics12081270
APA StyleMorina, J. C., & Franklin, R. B. (2023). Drivers of Antibiotic Resistance Gene Abundance in an Urban River. Antibiotics, 12(8), 1270. https://doi.org/10.3390/antibiotics12081270