
Recent Evolution of Susceptibility to Beta-Lactams in Neisseria meningitidis


Abstract
1. Introduction
2. Results
2.1. Beta-Lactam Susceptibility Phenotypes
2.2. Distribution of penA Alleles
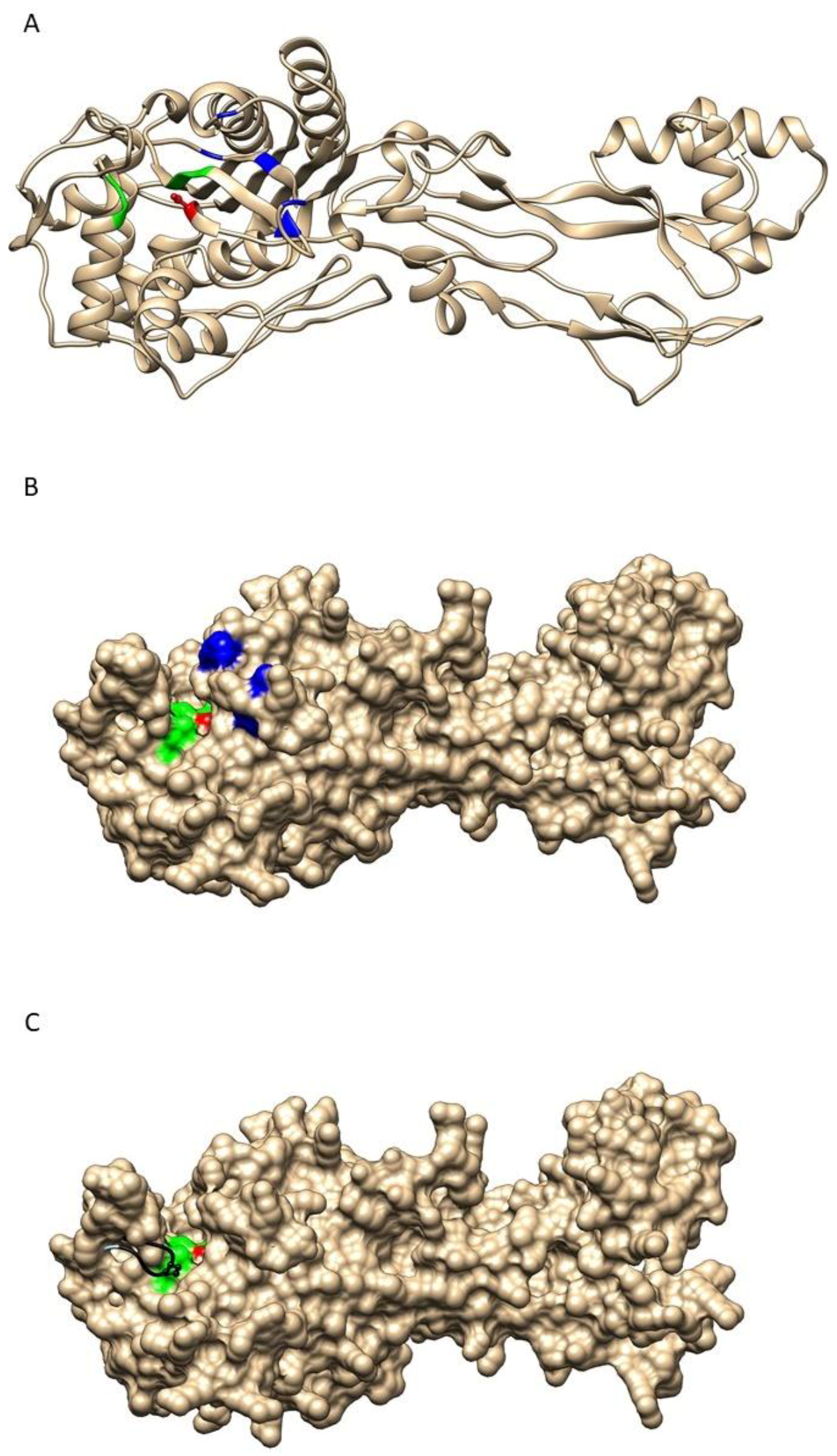
2.3. Structure-Function Analysis of Wild-Type and Modified PBP2
3. Discussion
4. Materials and Methods
4.1. Bacterial Methods
4.2. DNA Sequencing
4.3. Structure-Function Analysis
5. Limitations and Future Implications
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yazdankhah, S.P.; Caugant, D.A. Neisseria meningitidis: An overview of the carriage state. J. Med. Microbiol. 2004, 53, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.; Dias, J.G.; Ramliden, M.; Kodmon, C.; Economopoulou, A.; Beer, N.; Pastore Celentano, L.; Kanitz, E.; Richter, L.; Mattheus, W.; et al. The epidemiology of invasive meningococcal disease in EU/EEA countries, 2004-2014. Vaccine 2017, 35, 2034–2041. [Google Scholar] [CrossRef]
- Cartwright, K.A.; Reilly, S.; White, D.; Stuart, J. Early treatment with parenteral penicillin in meningococcal disease. Bmj 1992, 305, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Rosenstein, N.E.; Perkins, B.A.; Stephens, D.S.; Popovic, T.; Hughes, J.M. Meningococcal disease. N. Engl. J. Med. 2001, 344, 1378–1388. [Google Scholar] [CrossRef]
- Nadel, S. Treatment of Meningococcal Disease. J. Adolesc. Health 2016, 59 (Suppl. S2), S21–S28. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, J.A.; Arreaza, L.; Block, C.; Ehrhard, I.; Gray, S.J.; Heuberger, S.; Hoffmann, S.; Kriz, P.; Nicolas, P.; Olcen, P.; et al. Interlaboratory comparison of agar dilution and Etest methods for determining the MICs of antibiotics used in management of Neisseria meningitidis infections. Antimicrob. Agents Chemother. 2003, 47, 3430–3434. [Google Scholar] [CrossRef] [PubMed]
- Zapun, A.; Morlot, C.; Taha, M.K. Resistance to beta-Lactams in Neisseria ssp Due to Chromosomally Encoded Penicillin-Binding Proteins. Antibiotics 2016, 5, 35. [Google Scholar] [CrossRef] [PubMed]
- Vacca, P.; Fazio, C.; Neri, A.; Ambrosio, L.; Palmieri, A.; Stefanelli, P. Neisseria meningitidis Antimicrobial Resistance in Italy, 2006 to 2016. Antimicrob. Agents Chemother. 2018, 62, e00207-18. [Google Scholar] [CrossRef] [PubMed]
- Spratt, B.G.; Bowler, L.D.; Zhang, Q.Y.; Zhou, J.; Smith, J.M. Role of interspecies transfer of chromosomal genes in the evolution of penicillin resistance in pathogenic and commensal Neisseria species. J. Mol. Evol. 1992, 34, 115–125. [Google Scholar] [CrossRef]
- Powell, A.J.; Tomberg, J.; Deacon, A.M.; Nicholas, R.A.; Davies, C. Crystal structures of penicillin-binding protein 2 from penicillin-susceptible and -resistant strains of Neisseria gonorrhoeae reveal an unexpectedly subtle mechanism for antibiotic resistance. J. Biol. Chem. 2009, 284, 1202–1212. [Google Scholar] [CrossRef] [PubMed]
- Tipper, D.J.; Strominger, J.L. Mechanism of action of penicillins: A proposal based on their structural similarity to acyl-D-alanyl-D-alanine. Proc. Natl. Acad. Sci. USA 1965, 54, 1133–1141. [Google Scholar] [CrossRef]
- Bennett, D.E.; Meyler, K.L.; Cafferkey, M.T.; Cunney, R.J. Antibiotic susceptibility and molecular analysis of invasive Neisseria meningitidis recovered in the Republic of Ireland, 1996 to 2016. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Deghmane, A.E.; Hong, E.; Taha, M.K. Emergence of meningococci with reduced susceptibility to third-generation cephalosporins. J. Antimicrob. Chemother. 2017, 72, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Hong, E.; Deghmane, A.E.; Taha, M.K. Acquisition of beta-lactamase by Neisseria meningitidis through possible horizontal gene transfer. Antimicrob. Agents Chemother. 2018, 62, e00831-18. [Google Scholar] [CrossRef]
- Tsang, R.S.W.; Ahmad, T.; Jamieson, F.B.; Tyrrell, G.J. WGS analysis of a penicillin-resistant Neisseria meningitidis strain containing a chromosomal ROB-1 beta-lactamase gene. J. Antimicrob. Chemother. 2019, 74, 22–28. [Google Scholar] [PubMed]
- McNamara, L.A.; Potts, C.; Blain, A.E.; Retchless, A.C.; Reese, N.; Swint, S.; Lonsway, D.; Karlsson, M.; Lunquest, K.; Sweitzer, J.J.; et al. Detection of Ciprofloxacin-Resistant, beta-Lactamase-Producing Neisseria meningitidis Serogroup Y Isolates—United States, 2019–2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 735–739. [Google Scholar] [CrossRef]
- Coldiron, M.E.; Assao, B.; Page, A.L.; Hitchings, M.D.T.; Alcoba, G.; Ciglenecki, I.; Langendorf, C.; Mambula, C.; Adehossi, E.; Sidikou, F.; et al. Single-dose oral ciprofloxacin prophylaxis as a response to a meningococcal meningitis epidemic in the African meningitis belt: A 3-arm, open-label, cluster-randomized trial. PLoS Med. 2018, 15, e1002593. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.K.; Deghmane, A.E. Impact of COVID-19 pandemic and the lockdown on invasive meningococcal disease. BMC Res. Notes 2020, 13, 399. [Google Scholar] [CrossRef]
- Deghmane, A.-E.; Taha, M.-K. Changes in Invasive Neisseria meningitidis and Haemophilus influenzae Infections in France during the COVID-19 Pandemic. Microorganisms 2022, 10, 907. [Google Scholar] [CrossRef]
- Thulin, S.; Olcén, P.; Fredlund, H.; Unemo, M. Total variation in the penA gene of Neisseria meningitidis: Correlation between susceptibility to beta-lactam antibiotics and penA gene heterogeneity. Antimicrob. Agents Chemother. 2006, 50, 3317–3324. [Google Scholar] [CrossRef]
- Sauvage, E.; Kerff, F.; Terrak, M.; Ayala, J.A.; Charlier, P. The penicillin-binding proteins: Structure and role in peptidoglycan biosynthesis. FEMS Microbiol. Rev. 2008, 32, 234–258. [Google Scholar] [CrossRef]
- Potts, C.C.; Rodriguez-Rivera, L.D.; Retchless, A.C.; Hu, F.; Marjuki, H.; Blain, A.E.; McNamara, L.A.; Wang, X. Antimicrobial Susceptibility Survey of Invasive Neisseria meningitidis, United States 2012–2016. J. Infect. Dis. 2022. [Google Scholar] [CrossRef] [PubMed]
- Willerton, L.; Lucidarme, J.; Walker, A.; Lekshmi, A.; Clark, S.A.; Walsh, L.; Bai, X.; Lee-Jones, L.; Borrow, R. Antibiotic resistance among invasive Neisseria meningitidis isolates in England, Wales and Northern Ireland (2010/11 to 2018/19). PLoS ONE 2021, 16, e0260677. [Google Scholar] [CrossRef] [PubMed]
- Fedarovich, A.; Cook, E.; Tomberg, J.; Nicholas, R.A.; Davies, C. Structural effect of the Asp345a insertion in penicillin-binding protein 2 from penicillin-resistant strains of Neisseria gonorrhoeae. Biochemistry 2014, 53, 7596–7603. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, D.S., Jr.; Peacock, W.L., Jr.; Deacon, W.E.; Brown, L.; Pirkle, D.I. Neisseria gonorrhoeae. I. Virulence Genetically Linked to Clonal Variation. J. Bacteriol. 1963, 85, 1274–1279. [Google Scholar] [CrossRef]
- Jolley, K.A.; Maiden, M.C. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinform. 2010, 11, 595. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]


{kind=link}
| penA Allele | N° of Isolates (%) | Presence of Mutation in the 5 Critical Residues | GM of Penicillin G MIC (95% CI) |
|---|---|---|---|
| 1 | 365 (29.3) | no | 0.05 (0.05–0.05) |
| 9 | 208 (17.7) | yes | 0.22 (0.21–0.24) |
| 22 | 148 (11.9) | no | 0.06 (0.05–0.07) |
| 2 | 69 (5.5) | no | 0.06 (0.05–0.06) |
| 386 | 52 (4.2) | yes | 0.14 (0.13–0.16) |
| 3 | 46 (3.7) | no | 0.05 (0.05–0.06) |
| 14 | 42 (3.4) | yes | 0.17 (0.15–0.19) |
| 33 | 39 (3.1) | yes | 0.22 (0.19–0.26) |
| 327 | 30 (2.4) | yes | 0.43 (0.38–0.49) |
| 7 | 18 (1.4) | yes | 0.19 (0.16–0.23) |
| 13 | 17 (1.4) | yes | 0.33 (0.29–0.37) |
| 5 | 14 (1.1) | no | 0.05 (0.04–0.06) |
| 34 | 14 (1.1) | no | 0.05 (0.04–0.07) |
| 12 | 13 (1.0) | yes | 0.20 (0.18–0.23) |
| 19 | 13 (1.0) | yes | 0.21 (0.15–0.3) |
| 27 | 12 (1.0) | no | 0.04 (0.03–0.05) |
| 36 | 12 (1.0) | yes | 0.25 (0.19–0.35) |
| 295 | 12 (1.0) | yes | 0.26 (0.22–0.31) |
| 81 | 9 (0.7) | yes | 0.25 (0.17–0.36) |
| 179 | 9 (0.7) | yes | 0.41 (0.29–0.57) |
| 10 | 6 (0.5) | yes | 0.21 (0.16–0.29) |
| 15 | 6 (0.5) | yes | 0.17 (0.11–0.27) |
| 248 | 6 (0.5) | yes | 0.19 (0.15–0.25) |
| 908 | 6 (0.5) | yes | 0.11 (0.04–0.33) |
| Parameter | Value |
|---|---|
| Sensitivity | 0.94 |
| Specificity | 0.97 |
| Positive predictive value | 0.96 |
| Negative predictive value | 0.96 |
| Accuracy | 0.96 |
| Kapp coefficient | 0.91 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deghmane, A.-E.; Hong, E.; Taha, M.-K. Recent Evolution of Susceptibility to Beta-Lactams in Neisseria meningitidis. Antibiotics 2023, 12, 992. https://doi.org/10.3390/antibiotics12060992
Deghmane A-E, Hong E, Taha M-K. Recent Evolution of Susceptibility to Beta-Lactams in Neisseria meningitidis. Antibiotics. 2023; 12(6):992. https://doi.org/10.3390/antibiotics12060992
Chicago/Turabian StyleDeghmane, Ala-Eddine, Eva Hong, and Muhamed-Kheir Taha. 2023. "Recent Evolution of Susceptibility to Beta-Lactams in Neisseria meningitidis" Antibiotics 12, no. 6: 992. https://doi.org/10.3390/antibiotics12060992
APA StyleDeghmane, A.-E., Hong, E., & Taha, M.-K. (2023). Recent Evolution of Susceptibility to Beta-Lactams in Neisseria meningitidis. Antibiotics, 12(6), 992. https://doi.org/10.3390/antibiotics12060992