
Prevalence and Associated Factors of optrA-Positive-Enterococcus faecalis in Different Reservoirs around Farms in Vietnam

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Results
2.1. Demographic Characteristics
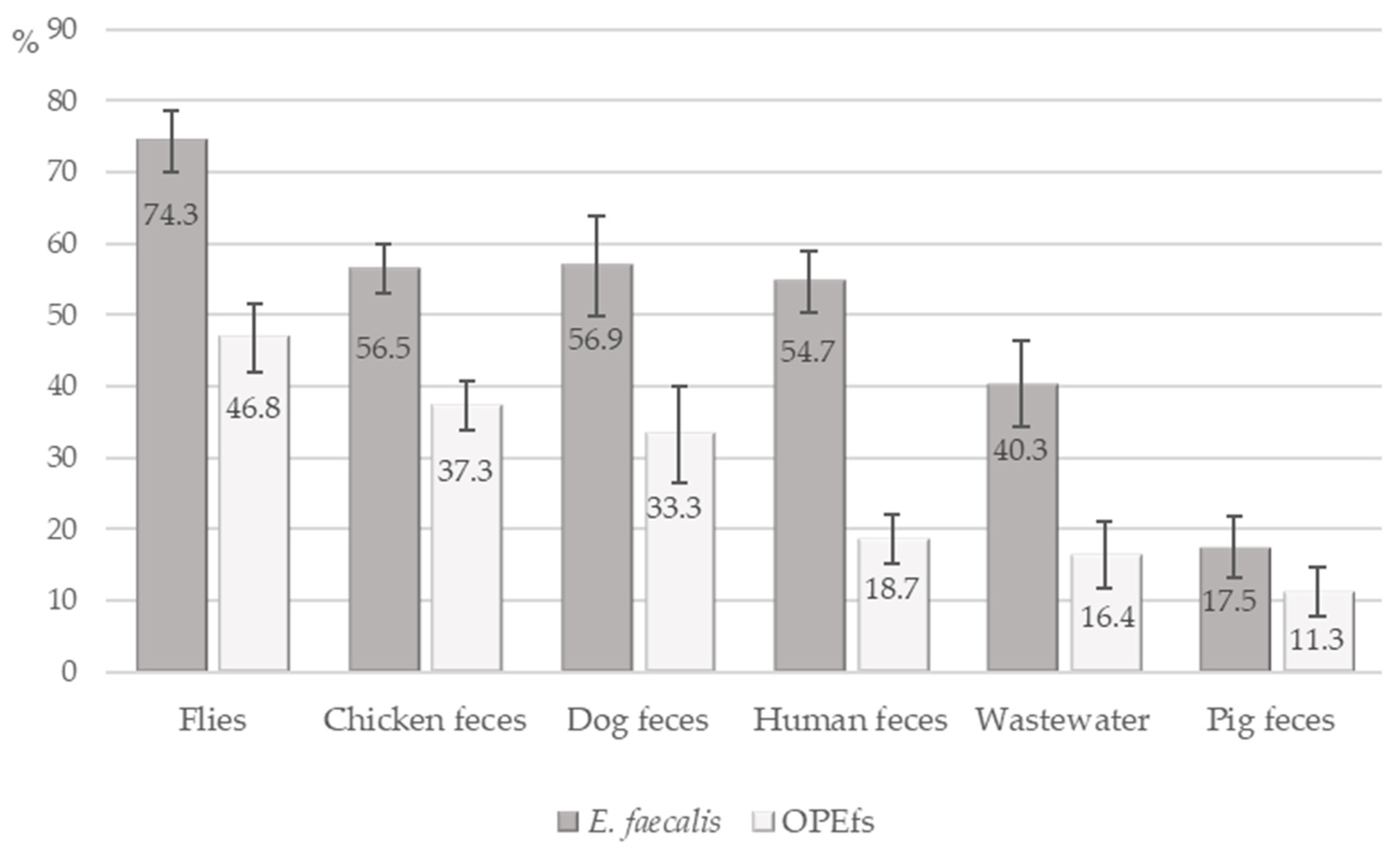
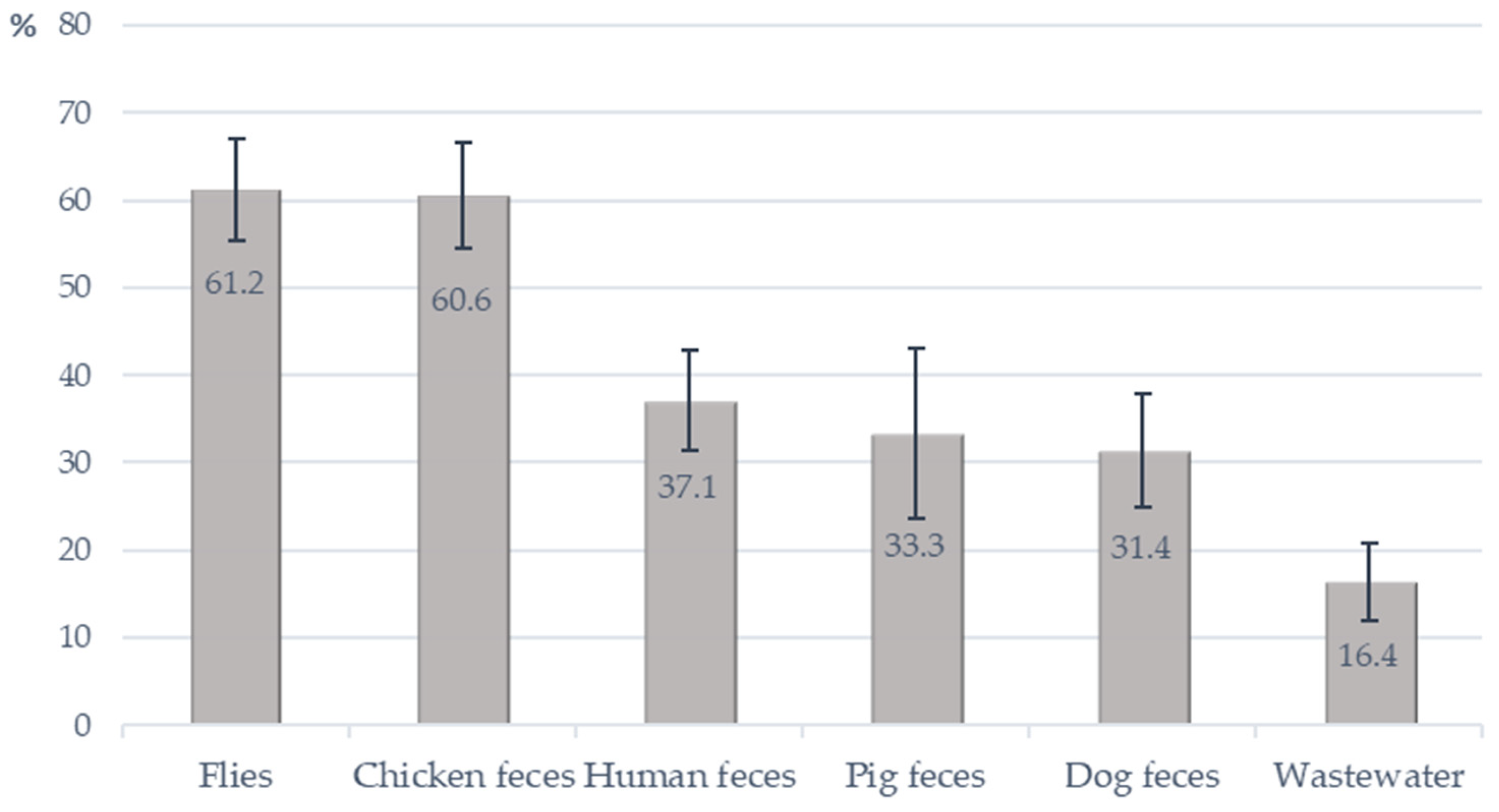
2.2. Prevalence of E. faecalis and OPEfs in Collected Samples
2.3. Associated Epidemiology Factors of OPEfs in Different Sample Types
2.4. OptrA-Positive E. faecalis Status and Antibiotic Resistance
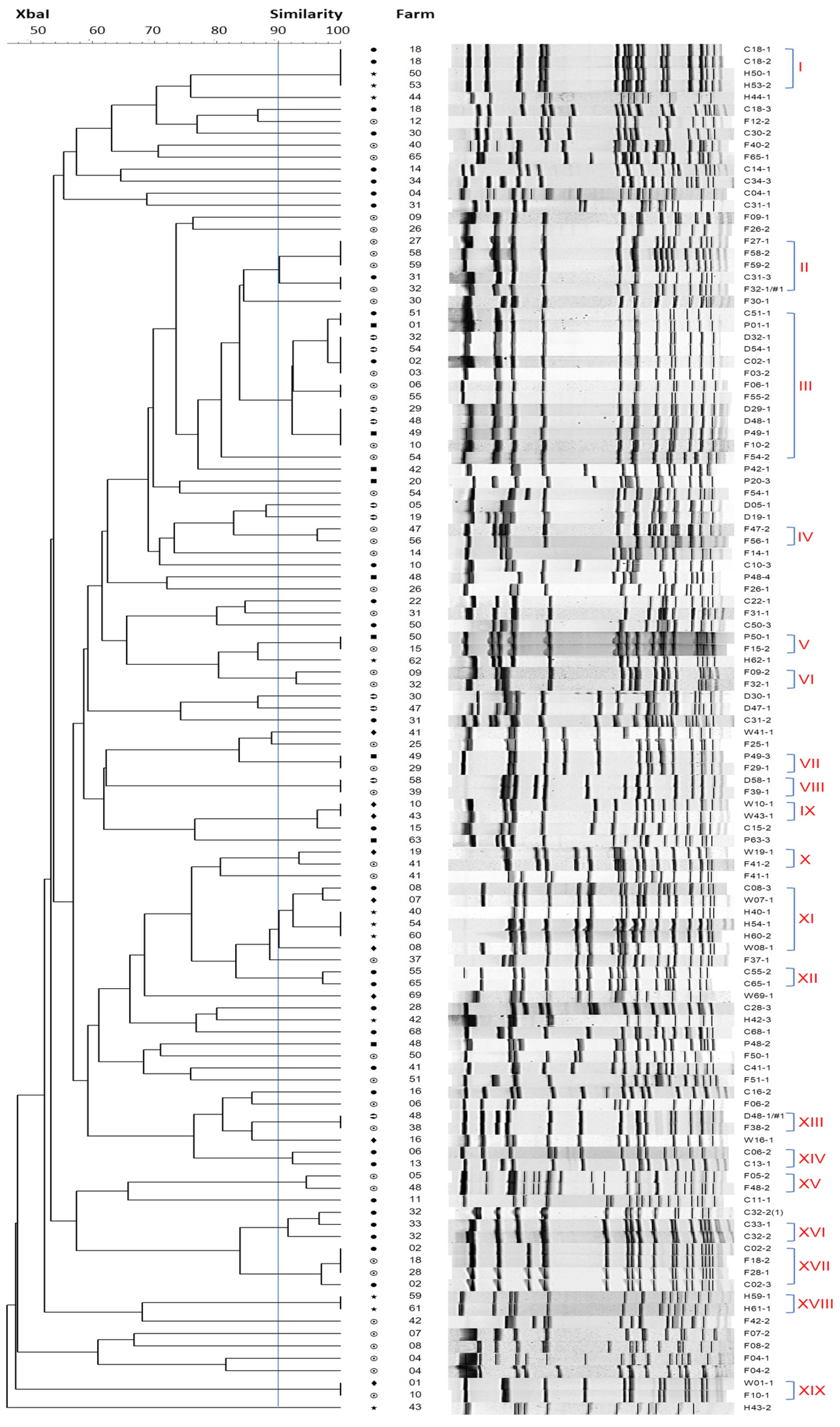
2.5. Molecular Typing of optrA-Positive E. faecalis by PFGE
3. Discussion
4. Materials and Methods
4.1. Setting and Population
4.2. Sample Collection and Laboratory Analysis
4.3. Antimicrobial Susceptibility Testing
4.4. Epidemiological Data Collection and Analysis
4.5. Pulsed-Field Gel Electrophoresis (PFGE)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, D.N.; Schluenzen, F.; Harms, J.M.; Starosta, A.L.; Connell, S.R.; Fucini, P. The oxazolidinone antibiotics perturb the ribosomal peptidyl-transferase center and effect tRNA positioning. Proc. Natl. Acad. Sci. USA 2008, 105, 13339–13344. [Google Scholar] [CrossRef]
- Moellering, R.C. Linezolid: The first oxazolidinone antimicrobial. Ann. Intern. Med. 2003, 138, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Moon, D.C.; Kim, S.J.; Mechesso, A.F.; Song, H.J.; Kang, H.Y.; Choi, J.H.; Yoon, S.S.; Lim, S.K. Nationwide Surveillance on Antimicrobial Resistance Profiles of Enterococcus faecium and Enterococcus faecalis Isolated from Healthy Food Animals in South Korea, 2010 to 2019. Microorganisms 2021, 9, 925. [Google Scholar] [CrossRef] [PubMed]
- Hammerum, A.M. Enterococci of animal origin and their significance for public health. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2012, 18, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, L.L.; Bisgaard, M.; Son, N.T.; Trung, N.V.; An, H.M.; Dalsgaard, A. Enterococcus faecalis clones in poultry and in humans with urinary tract infections, Vietnam. Emerg. Infect. Dis. 2012, 18, 1096–1100. [Google Scholar] [CrossRef]
- Gaglio, R.; Couto, N.; Marques, C.; de Fatima Silva Lopes, M.; Moschetti, G.; Pomba, C.; Settanni, L. Evaluation of antimicrobial resistance and virulence of enterococci from equipment surfaces, raw materials, and traditional cheeses. Int. J. Food Microbiol. 2016, 236, 107–114. [Google Scholar] [CrossRef]
- Fozouni, L.; Askari, H.; Pordeli, H.R. Frequency Distribution of Fluoroquinolones-Resistant Enterococcus faecalis Isolates from Patients with Prostatitis in Golestan Province, Iran. Med. Lab. J. 2019, 13, 29–33. [Google Scholar] [CrossRef]
- Zou, J.; Xia, Y. Molecular characteristics and risk factors associated with linezolid-resistant Enterococcus faecalis infection in Southwest China. J. Glob. Antimicrob. Resist. 2020, 22, 504–510. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, F.; Bai, B.; Lin, Z.; Xu, G.; Chen, Z.; Sun, X.; Zheng, J.; Deng, Q.; Yu, Z. Linezolid Resistance in Enterococcus faecalis Associated with Urinary Tract Infections of Patients in a Tertiary Hospitals in China: Resistance Mechanisms, Virulence, and Risk Factors. Front. Public Health 2021, 9, 570650. [Google Scholar] [CrossRef]
- Anh Tram, Q. Study on antibiotic resistance characteristics of gram-positive strains of bacteria causing urinary tract infections in Nghe An General Friendship Hospital. Tạp chí Y học Việt Nam 2022, 517, 257–261. [Google Scholar] [CrossRef]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti-Infect. Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef] [PubMed]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Dicuonzo, G.; Gherardi, G.; Lorino, G.; Angeletti, S.; Battistoni, F.; Bertuccini, L.; Creti, R.; Di Rosa, R.; Venditti, M.; Baldassarri, L. Antibiotic resistance and genotypic characterization by PFGE of clinical and environmental isolates of enterococci. FEMS Microbiol. Lett. 2001, 201, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Daniel, D.S.; Lee, S.M.; Gan, H.M.; Dykes, G.A.; Rahman, S. Genetic diversity of Enterococcus faecalis isolated from environmental, animal and clinical sources in Malaysia. J. Infect. Public Health 2017, 10, 617–623. [Google Scholar] [CrossRef]
- Usui, M.; Ozawa, S.; Onozato, H.; Kuge, R.; Obata, Y.; Uemae, T.; Ngoc, P.T.; Heriyanto, A.; Chalemchaikit, T.; Makita, K.; et al. Antimicrobial susceptibility of indicator bacteria isolated from chickens in Southeast Asian countries (Vietnam, Indonesia and Thailand). J. Vet. Med. Sci. 2014, 76, 685–692. [Google Scholar] [CrossRef]
- Ruiz-Ripa, L.; Feßler, A.T.; Hanke, D.; Eichhorn, I.; Azcona-Gutiérrez, J.M.; Pérez-Moreno, M.O.; Seral, C.; Aspiroz, C.; Alonso, C.A.; Torres, L.; et al. Mechanisms of Linezolid Resistance Among Enterococci of Clinical Origin in Spain-Detection of optrA- and cfr(D)-Carrying E. faecalis. Microorganisms 2020, 8, 1155. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, Y.; Cai, J.; Schwarz, S.; Cui, L.; Hu, Z.; Zhang, R.; Li, J.; Zhao, Q.; He, T.; et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin. J. Antimicrob. Chemother. 2015, 70, 2182–2190. [Google Scholar] [CrossRef]
- He, T.; Shen, Y.; Schwarz, S.; Cai, J.; Lv, Y.; Li, J.; Feßler, A.T.; Zhang, R.; Wu, C.; Shen, J.; et al. Genetic environment of the transferable oxazolidinone/phenicol resistance gene optrA in Enterococcus faecalis isolates of human and animal origin. J. Antimicrob. Chemother. 2016, 71, 1466–1473. [Google Scholar] [CrossRef]
- Yi, M.; Zou, J.; Zhao, J.; Tang, Y.; Yuan, Y.; Yang, B.; Huang, J.; Xia, P.; Xia, Y. Emergence of optrA-Mediated Linezolid Resistance in Enterococcus faecium: A Molecular Investigation in a Tertiary Hospital of Southwest China from 2014–2018. Infect. Drug Resist. 2022, 15, 13–20. [Google Scholar] [CrossRef]
- Sadowy, E. Linezolid resistance genes and genetic elements enhancing their dissemination in enterococci and streptococci. Plasmid 2018, 99, 89–98. [Google Scholar] [CrossRef]
- Freitas, A.R.; Tedim, A.P.; Novais, C.; Lanza, V.F.; Peixe, L. Comparative genomics of global optrA-carrying Enterococcus faecalis uncovers a common chromosomal hotspot for optrA acquisition within a diversity of core and accessory genomes. Microb. Genom. 2020, 6, e000350. [Google Scholar] [CrossRef]
- Yoon, S.; Kim, Y.B.; Seo, K.W.; Ha, J.S.; Noh, E.B.; Lee, Y.J. Characteristics of linezolid-resistant Enterococcus faecalis isolates from broiler breeder farms. Poult. Sci. 2020, 99, 6055–6061. [Google Scholar] [CrossRef]
- Almeida, L.M.; Lebreton, F.; Gaca, A.; Bispo, P.M.; Saavedra, J.T.; Calumby, R.N.; Grillo, L.M.; Nascimento, T.G.; Filsner, P.H.; Moreno, A.M.; et al. Transferable Resistance Gene optrA in Enterococcus faecalis from Swine in Brazil. Antimicrob. Agents Chemother. 2020, 64, e00142-20. [Google Scholar] [CrossRef]
- Kim, D.P.; Saegerman, C.; Douny, C.; Dinh, T.V.; Xuan, B.; Vu, B.D.; Hong, N.P.; Scippo, M.-L. First Survey on the Use of Antibiotics in Pig and Poultry Production in the Red River Delta Region of Vietnam. Food Public Health 2013, 3, 247–256. [Google Scholar]
- Di, K.N.; Pham, D.T.; Tee, T.S.; Binh, Q.A.; Nguyen, T.C. Antibiotic usage and resistance in animal production in Vietnam: A review of existing literature. Trop. Anim. Health Prod. 2021, 53, 340. [Google Scholar] [CrossRef]
- Luu, Q.H.; Nguyen, T.L.A.; Pham, T.N.; Vo, N.G.; Padungtod, P. Antimicrobial use in household, semi-industrialized, and industrialized pig and poultry farms in Viet Nam. Prev. Vet. Med. 2021, 189, 105292. [Google Scholar] [CrossRef]
- Tamang, M.D.; Moon, D.C.; Kim, S.R.; Kang, H.Y.; Lee, K.; Nam, H.M.; Jang, G.C.; Lee, H.S.; Jung, S.C.; Lim, S.K. Detection of novel oxazolidinone and phenicol resistance gene optrA in enterococcal isolates from food animals and animal carcasses. Vet. Microbiol. 2017, 201, 252–256. [Google Scholar] [CrossRef]
- Rousham, E.K.; Unicomb, L.; Islam, M.A. Human, animal and environmental contributors to antibiotic resistance in low-resource settings: Integrating behavioural, epidemiological and One Health approaches. Proc. Biol. Sci. 2018, 285, 20180332. [Google Scholar] [CrossRef]
- Nguyen, P.T.L.; Tran, H.T.M.; Tran, H.A.; Pham, T.D.; Luong, T.M.; Nguyen, T.H.; Nguyen, L.T.P.; Nguyen, T.T.T.; Hoang, H.T.A.; Nguyen, C.; et al. Carriage of Plasmid-Mediated Colistin Resistance-1-Positive Escherichia coli in Humans, Animals, and Environment on Farms in Vietnam. Am. J. Trop. Med. Hyg. 2022, 107, 65–71. [Google Scholar] [CrossRef]
- Rawat, N.; Anjali; Shreyata; Sabu, B.; Jamwal, R.; Devi, P.P.; Yadav, K.; Raina, H.S.; Rajagopal, R. Understanding the role of insects in the acquisition and transmission of antibiotic resistance. Sci. Total Environ. 2023, 858, 159805. [Google Scholar] [CrossRef]
- Fletcher, S. Understanding the contribution of environmental factors in the spread of antimicrobial resistance. Environ. Health Prev. Med. 2015, 20, 243–252. [Google Scholar] [CrossRef] [PubMed]
- WHO. Critically Important Antimicrobials for Human Medicine, 6th Revision. 2018, p. 14. Available online: https://apps.who.int/iris/bitstream/handle/10665/312266/9789241515528-eng.pdf (accessed on 18 February 2023).
- Daniel, D.S.; Lee, S.M.; Dykes, G.A.; Rahman, S. Public Health Risks of Multiple-Drug-Resistant Enterococcus spp. in Southeast Asia. Appl. Environ. Microbiol. 2015, 81, 6090–6097. [Google Scholar] [CrossRef] [PubMed]
- Gião, J.; Leão, C.; Albuquerque, T.; Clemente, L.; Amaro, A. Antimicrobial Susceptibility of Enterococcus Isolates from Cattle and Pigs in Portugal: Linezolid Resistance Genes optrA and poxtA. Antibiotics 2022, 11, 615. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Pan, H.; Lou, Y.; Wu, Z.; Zhang, J.; Huang, Y.; Yu, W.; Qiu, Y. Epidemiological characteristics and genetic structure of linezolid-resistant Enterococcus faecalis. Infect. Drug Resist. 2018, 11, 2397–2409. [Google Scholar] [CrossRef]
- MRAD. Vietnam Ministry of Agriculture & Rural Development, Circulars 10/2016TT-BNNPTN, List of Antibiotics Allowed to Be Used in Animal Feed. Available online: https://vanban.chinhphu.vn/default.aspx?pageid=27160&docid=186403 (accessed on 22 March 2023).
- Nguyen, V.T.; Carrique-Mas, J.J.; Ngo, T.H.; Ho, H.M.; Ha, T.T.; Campbell, J.I.; Nguyen, T.N.; Hoang, N.N.; Pham, V.M.; Wagenaar, J.A.; et al. Prevalence and risk factors for carriage of antimicrobial-resistant Escherichia coli on household and small-scale chicken farms in the Mekong Delta of Vietnam. J. Antimicrob. Chemother. 2015, 70, 2144–2152. [Google Scholar] [CrossRef]
- Nguyen, T.K.; Nguyen, L.T.; Chau, T.T.H.; Nguyen, T.T.; Tran, B.N.; Taniguchi, T.; Hayashidani, H.; Ly, K.T.L. Prevalence and antibiotic resistance of Salmonella isolated from poultry and its environment in the Mekong Delta, Vietnam. Vet. World 2021, 14, 3216–3223. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Jeggo, M. The One Health Approach-Why Is It So Important? Trop. Med. Infect. Dis. 2019, 4, 88. [Google Scholar] [CrossRef]
- CDC. One Health Basics. Available online: https://www.cdc.gov/onehealth/basics/index.html#print (accessed on 26 April 2023).
- World Health Organization. Global Action Plan on Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Tyson, G.H.; Sabo, J.L.; Hoffmann, M.; Hsu, C.H.; Mukherjee, S.; Hernandez, J.; Tillman, G.; Wasilenko, J.L.; Haro, J.; Simmons, M.; et al. Novel linezolid resistance plasmids in Enterococcus from food animals in the USA. J. Antimicrob. Chemother. 2018, 73, 3254–3258. [Google Scholar] [CrossRef]
- McHugh, M.P.; Parcell, B.J.; Pettigrew, K.A.; Toner, G.; Khatamzas, E.; El Sakka, N.; Karcher, A.M.; Walker, J.; Weir, R.; Meunier, D.; et al. Presence of optrA-mediated linezolid resistance in multiple lineages and plasmids of Enterococcus faecalis revealed by long read sequencing. Microbiology 2022, 168, 001137. [Google Scholar] [CrossRef]
- Cai, J.; Schwarz, S.; Chi, D.; Wang, Z.; Zhang, R.; Wang, Y. Faecal carriage of optrA-positive enterococci in asymptomatic healthy humans in Hangzhou, China. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2019, 25, 630.e1–630.e6. [Google Scholar] [CrossRef]
- Càmara, J.; Camoez, M.; Tubau, F.; Pujol, M.; Ayats, J.; Ardanuy, C.; Domínguez, M. Detection of the Novel optrA Gene Among Linezolid-Resistant Enterococci in Barcelona, Spain. Microb. Drug Resist. 2019, 25, 87–93. [Google Scholar] [CrossRef]
- Park, K.; Jeong, Y.S.; Chang, J.; Sung, H.; Kim, M.N. Emergence of optrA-Mediated Linezolid-Nonsusceptible Enterococcus faecalis in a Tertiary Care Hospital. Ann. Lab. Med. 2020, 40, 321–325. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, R.; Li, J.; Wu, Z.; Yin, W.; Schwarz, S.; Tyrrell, J.M.; Zheng, Y.; Wang, S.; Shen, Z.; et al. Comprehensive resistome analysis reveals the prevalence of NDM and MCR-1 in Chinese poultry production. Nat. Microbiol. 2017, 2, 16260. [Google Scholar] [CrossRef]
- Ström, G.; Halje, M.; Karlsson, D.; Jiwakanon, J.; Pringle, M.; Fernström, L.L.; Magnusson, U. Antimicrobial use and antimicrobial susceptibility in Escherichia coli on small- and medium-scale pig farms in north-eastern Thailand. Antimicrob. Resist. Infect. Control 2017, 6, 75. [Google Scholar] [CrossRef]
- Nkansa, M.; Agbekpornu, H.; Kikimoto, B.; Chandler, C. Antibiotic Use Among Poultry Farmers in the Dormaa Municipality, Ghana; Report for Fleming Fund Fellowship Programme; London School of Hygiene and Tropical Medicine: London, UK, 2020. [Google Scholar]
- Paintsil, E.K.; Ofori, L.A.; Akenten, C.W.; Fosu, D.; Ofori, S.; Lamshöft, M.; May, J.; Danso, K.O.; Krumkamp, R.; Dekker, D. Antimicrobial Usage in Commercial and Domestic Poultry Farming in Two Communities in the Ashanti Region of Ghana. Antibiotics 2021, 10, 800. [Google Scholar] [CrossRef]
- Doud, C.W.; Scott, H.M.; Zurek, L. Role of house flies in the ecology of Enterococcus faecalis from wastewater treatment facilities. Microb. Ecol. 2014, 67, 380–391. [Google Scholar] [CrossRef]
- Sruthi, B.; Ibrahim, S.S. Insect pests of medical importance and their management. In Insect Pests of Crops and Their Eco-Friendly Management; Bhumi Publishing: Maharashtra, India, 2023; pp. 211–214. [Google Scholar]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef]
- Alves, T.D.S.; Lara, G.H.B.; Maluta, R.P.; Ribeiro, M.G.; Leite, D.D.S. Carrier flies of multidrug-resistant Escherichia coli as potential dissemination agent in dairy farm environment. Sci. Total Environ. 2018, 633, 1345–1351. [Google Scholar] [CrossRef]
- Onwugamba, F.C.; Fitzgerald, J.R.; Rochon, K.; Guardabassi, L.; Alabi, A.; Kühne, S.; Grobusch, M.P.; Schaumburg, F. The role of ‘filth flies’ in the spread of antimicrobial resistance. Travel Med. Infect. Dis. 2018, 22, 8–17. [Google Scholar] [CrossRef]
- Macovei, L.; Zurek, L. Ecology of antibiotic resistance genes: Characterization of enterococci from houseflies collected in food settings. Appl. Environ. Microbiol. 2006, 72, 4028–4035. [Google Scholar] [CrossRef]
- Baureder, M.; Reimann, R.; Hederstedt, L. Contribution of catalase to hydrogen peroxide resistance in Enterococcus faecalis. FEMS Microbiol. Lett. 2012, 331, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Cockerill, F.R.; Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically: Approved Standard, 9th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- CDC. The National Antimicrobial Resistance Monitoring System for Enteric Bacteria. Glossary of Terms Related to Antibiotic Resistance. Available online: https://www.cdc.gov/narms/resources/glossary.html#print (accessed on 22 March 2023).
- Loftus, M.J.; Everts, R.J.; Cheng, A.C.; Eti, P.; Fakasiieiki, T.; Isaia, L.; Isopo, E.; Jenney, A.W.J.; Lameko, V.; Leaupepe, H.; et al. Antimicrobial susceptibility of bacterial isolates from clinical specimens in four Pacific Island countries, 2017–2021. Lancet Reg. Health—West. Pac. 2023, 32, 100677. [Google Scholar] [CrossRef] [PubMed]
- CLSI. M100-ED32:2022 Performance Standards for Antimicrobial Susceptibility Testing, 32nd Edition. Available online: http://em100.edaptivedocs.net/GetDoc.aspx?doc=CLSI%20M100%20ED32:2022&xormat=SPDF&src=BB (accessed on 28 June 2022).
- Vietnamese Government. Decree 13/2020/ND-CP Detail Guideline of Livestock Law; Vietnamese Government: Hanoi, Vietnam, 2020.
- Centers for Disease Control and Prevention (CDC). Unified Pulsed-Field Gel Electrophoresis (PFGE) Protocol for Gram Positive Bacteria. Available online: https://www.cdc.gov/hai/pdfs/labsettings/unified_pfge_protocol.pdf (accessed on 10 February 2023).
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | n | % | 95% CI |
|---|---|---|---|
| Age groups (Years, N = 139) | |||
| 18–40 | 25 | 18.0 | 11.5–24.5 |
| 41–60 | 79 | 56.8 | 48.9–64.7 |
| >60 | 35 | 25.2 | 18.0–32.4 |
| Min = 22 | Max = 85 | Mean = 51.9 | |
| Sex (N = 139) | |||
| Male | 78 | 56.1 | 47.7–64.2 |
| Female | 61 | 43.9 | 35.8–52.3 |
| Education level (N = 139) | |||
| Primary school | 14 | 10.1 | 5.8–15.1 |
| Secondary school | 90 | 64.7 | 56.9–72.7 |
| High school and above | 35 | 25.2 | 18.0–32.4 |
| Total livestock unit of farm | Min = 0.22 | Max = 37.04 | Mean = 9.56 SD = 11.84 |
| Farm scales (N = 139) | |||
| Household | 98 | 70.5 | 62.3–77.6 |
| Small | 24 | 17.3 | 11.8–24.6 |
| Medium | 17 | 12.2 | 7.7–18.9 |
| Farm scales (N = 70) | |||
| Household | 51 | 72.9 | 61.0–82.2 |
| Small | 12 | 17.1 | 9.9–29.1 |
| Medium | 7 | 10.0 | 4.8–19.8 |
| Farm types (N = 70) | |||
| Chicken | 46 | 65.7 | 53.6–76.1 |
| Swine | 4 | 5.7 | 2.1–14.6 |
| Mixed | 20 | 28.6 | 19.0–40.5 |
| Factors | OPEfs in Sample | OR (95% CI) | p | Adjusted OR (95% CI) | p | |||
|---|---|---|---|---|---|---|---|---|
| Negative | Positive | |||||||
| n | % | n | % | |||||
| Human (N = 139) | ||||||||
| Age groups | ||||||||
| 18–40 | 22 | 88 | 3 | 12 | 1 | - | - | |
| 41–60 | 62 | 78.5 | 17 | 21.5 | 2.01 (0.54–7.53) | 0.30 | - | - |
| >60 | 29 | 82.9 | 6 | 17.1 | 1.52 (0.34–6.75) | 0.58 | - | - |
| Sex | ||||||||
| Female | 51 | 83.6 | 10 | 16.4 | 1 | |||
| Male | 62 | 79.5 | 16 | 17.1 | 1.32 (0.55–3.15) | 0.54 | - | - |
| Education level | ||||||||
| Primary school | 10 | 71.4 | 4 | 28.6 | 3.1 (0.65–14.73) | 0.16 | - | - |
| Secondary school | 72 | 80 | 18 | 20 | 1.94 (0.61–6.19) | 0.27 | - | - |
| High school and above | 31 | 88.6 | 4 | 11.4 | 1 | - | - | |
| Farm scales | ||||||||
| Household | 83 | 84.7 | 15 | 15.3 | 1 | - | - | |
| Small | 16 | 66.7 | 8 | 33.3 | 2.77 (1.01–7.61) | 0.049 | - | - |
| Medium | 14 | 82.4 | 3 | 17.6 | 1.19 (0.30–4.63) | 0.81 | - | - |
| Total livestock unit of farm | ||||||||
| Mean = 9.2 | Mean = 11.07 | 0.01 (0.98–1.05) | 0.47 | - | - | |||
| Farm types | ||||||||
| Chicken | 74 | 83.1 | 15 | 16.9 | 0.74 (0.30–1.87) | 0.53 | - | - |
| Swine | 6 | 75.0 | 2 | 25.0 | 1.22 (0.21–7.11) | 0.82 | - | - |
| Mixed | 33 | 78.6 | 9 | 21.4 | 1 | - | - | |
| Average income (per month) | ||||||||
| <1 million dong | 25 | 80.6 | 6 | 19.4 | 1 | - | - | |
| ≥1 million dong | 88 | 81.5 | 20 | 18.5 | 0.95 (0.34–2.61) | 0.92 | - | - |
| Feeding area | ||||||||
| <1000 m2 | 81 | 83.5 | 16 | 16.5 | 1 | - | - | |
| ≥1000 m2 | 32 | 76.2 | 10 | 23.8 | 1.58(0.65–3.85) | 0.31 | - | - |
| Used antibiotics in livestock | ||||||||
| No | 34 | 89.5 | 4 | 10.5 | 1 | - | - | |
| Yes | 79 | 78.2 | 22 | 21.8 | 2.37 (0.76–7.39) | 0.12 | - | - |
| Used industrial food in livestock | ||||||||
| No | 12 | 100 | 0 | 0 | - | - | - | - |
| Yes | 101 | 79.5 | 26 | 20.5 | - | - | - | - |
| Chicken (N = 193) | ||||||||
| Farm scales | ||||||||
| Household | 90 | 66.2 | 46 | 33.8 | 1 | - | - | |
| Small | 22 | 61.1 | 14 | 38.9 | 1.25 (0.58–2.66) | 0.57 | - | - |
| Medium | 9 | 41.9 | 12 | 75.1 | 2.61 (1.02–6.64) | 0.04 | - | - |
| Total livestock unit of farm | ||||||||
| Mean = 7.8 | Mean = 11.8 | 1.03 (1.00–1.05) | 0.02 | 1.01 (0.98–1.05) | 0.40 | |||
| Farm types | ||||||||
| Chicken | 85 | 61.6 | 53 | 38.4 | 1.18 (0.62–2.20) | 0.62 | - | - |
| Mixed | 36 | 65.5 | 19 | 34.5 | 1 | - | - | |
| Average income (per month) | ||||||||
| <1 million dong | 26 | 51.0 | 25 | 49.0 | 1.94 (1.01–3.73) | 0.04 | 1.40 (0.68–2.87) | 0.36 |
| ≥1 million dong | 95 | 66.9 | 47 | 33.1 | 1 | 1 | ||
| Feeding area | ||||||||
| <1000 m2 | 95 | 68.8 | 43 | 31.2 | 1 | 1 | ||
| ≥1000 m2 | 26 | 47.3 | 29 | 52.7 | 2.46 (1.30–4.67) | 0.006 | 1.77 (0.77–4.06) | 0.18 |
| Used antibiotics in livestock | ||||||||
| No | 26 | 61.9 | 16 | 38.1 | 1 | - | - | |
| Yes | 95 | 62.9 | 56 | 37.1 | 0.96 (0.47–1.94) | 0.9 | - | - |
| Used industrial food in livestock | ||||||||
| No | 15 | 83.3 | 3 | 16.7 | 1 | - | - | |
| Yes | 106 | 60.6 | 69 | 39.4 | 3.26 (0.91–11.6) | 0.07 | - | - |
| Flies (N = 109) | ||||||||
| Farm scales | ||||||||
| Household | 50 | 61.7 | 31 | 38.3 | 1 | - | - | |
| Small | 6 | 35.3 | 11 | 64.7 | 2.96 (1.00–8.80) | 0.51 | - | - |
| Medium | 2 | 18.2 | 9 | 81.8 | 7.26 (1.47–35.82) | 0.02 | - | |
| Total livestock unit of farm | ||||||||
| Mean = 4.84 | Mean = 12.2 | 1.07 (1.03–1.12) | 0.002 | 1.06 (1.00–1.12) | 0.04 | |||
| Farm types | ||||||||
| Chicken | 30 | 45.5 | 36 | 54.5 | 2.22 (0.97–5.08) | 0.06 | - | - |
| Swine | 4 | 66.7 | 2 | 33.3 | 0.92 (0.15–5.73) | 0.93 | - | - |
| Mixed | 24 | 64.9 | 13 | 35.1 | 1 | - | - | |
| Average income (per month) | ||||||||
| <1 million dong | 10 | 40.0 | 15 | 60.0 | 2.0 (0.80–4.97) | 0.13 | - | - |
| ≥1 million dong | 48 | 57.1 | 36 | 42.9 | 1 | - | - | |
| Feeding area | ||||||||
| <1000 m2 | 48 | 60.0 | 32 | 40.0 | 1 | 1 | ||
| ≥1000 m2 | 10 | 34.5 | 10 | 65.5 | 2.85 (1.17–6.92) | 0.02 | 0.82 (0.23–3.00) | 0.77 |
| Used antibiotics in livestock | ||||||||
| No | 24 | 77.4 | 7 | 22.6 | 1 | 1 | ||
| Yes | 34 | 43.6 | 44 | 56.4 | 4.44 (1.71–11.51) | 0.002 | 3.35 (1.24–9.09) | 0.02 |
| Used industrial food in livestock | ||||||||
| No | 7 | 63.6 | 4 | 36.4 | 1 | - | ||
| Yes | 51 | 52.0 | 47 | 48.0 | 1.61 (0.44–1.86) | 0.47 | - | |
| Wastewater (N = 67) | ||||||||
| Farm scales | ||||||||
| Household | 43 | 89.6 | 5 | 10.4 | 1 | - | - | |
| Small | 10 | 83.3 | 2 | 16.7 | 1.72 (0.29–10.18) | 0.55 | - | - |
| Medium | 3 | 42.9 | 4 | 57.1 | 11.47 (1.97–66.66) | 0.007 | - | - |
| Total livestock unit of farm | ||||||||
| Mean = 7.14 | Mean = 18.66 | 1.08 (1.02–1.13) | 0.005 | 1.07 (0.99–1.13) | 0.052 | |||
| Farm types | ||||||||
| Chicken | 35 | 77.8 | 10 | 22.2 | 4.86 (0.57–41.10) | 0.15 | - | - |
| Swine | 4 | 100 | 0 | 0.0 | 0 | - | - | |
| Mixed | 17 | 94.4 | 1 | 5.6 | 1 | - | - | |
| Average income (per month) | ||||||||
| <1 million dong | 13 | 76.5 | 4 | 23.5 | 1.89 (0.48–7.48) | 0.36 | - | - |
| ≥1 million dong | 43 | 86.0 | 7 | 14.0 | 1 | - | - | |
| Feeding area | ||||||||
| <1000 m2 | 44 | 89.8 | 5 | 10.2 | 1 | 1 | ||
| ≥1000 m2 | 12 | 66.7 | 6 | 33.3 | 4.40 (1.14–16.9) | 0.03 | 1.7 (0.28–9.08) | 0.59 |
| Used antibiotics in livestock | ||||||||
| No | 16 | 94.1 | 1 | 5.9 | 1 | - | - | |
| Yes | 40 | 80 | 10 | 20 | 4.0 (0.47–33.85) | 0.20 | - | - |
| Used industrial food in livestock | ||||||||
| No | 4 | 80 | 1 | 20 | 1 | - | - | |
| Yes | 52 | 83.9 | 10 | 16.1 | 0.77 (0.08–7.62) | 0.82 | - | - |
| Sample Types | optrA | n | Resistance to Investigating Antimicrobials | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TE | MN | DXT | E | C | CIP | LEV | HLGR | HLSR | LNZ | |||||||||||||
| n | % | n | % | n | % | n | % | n | % | n | % | n | % | n | % | n | % | n | % | |||
| Human feces | Pos | 26 | 19 | 73.1 | 15 | 57.7 | 18 | 63.9 | 12 | 46.2 | 12 | 46.2 | 1 | 3.8 | 1 | 3.8 | 2 | 7.7 | 5 | 19.2 | 22 | 86.4 |
| Neg | 50 | 33 | 66.0 | 20 | 40.0 | 28 | 56.0 | 14 | 28.0 | 12 | 24.0 | 1 | 2.0 | 1 | 2.0 | 1 | 2.0 | 0 | 0.0 | 2 | 4.0 | |
| Total | 76 | 52 | 68.4 | 35 | 46.0 | 46 | 60.5 | 26 | 34.2 | 24 | 31.6 | 2 | 2.6 | 2 | 2.6 | 3 | 3.9 | 5 | 6.6 | 24 | 31.6 | |
| Chicken feces | Pos | 72 | 70 | 97.2 | 68 | 62.4 | 65 | 90.3 | 64 | 88.9 | 66 | 91.7 | 20 | 28.7 | 16 | 22.2 | 11 | 15.3 | 31 | 43.1 | 65 | 90.3 |
| Neg | 37 | 28 | 75.7 | 18 | 43.2 | 25 | 76.6 | 13 | 35.1 | 5 | 13.5 | 2 | 5.4 | 0 | 0.0 | 1 | 2.7 | 4 | 10.8 | 1 | 2.7 | |
| Total | 109 | 98 | 89.9 | 86 | 78.9 | 90 | 82.6 | 77 | 70.6 | 71 | 65.1 | 22 | 20.2 | 16 | 14.7 | 12 | 11.0 | 35 | 32.1 | 66 | 60.6 | |
| Pig feces | Pos | 9 | 9 | 100 | 6 | 66.7 | 8 | 88.9 | 8 | 88.9 | 6 | 66.7 | 2 | 22.2 | 2 | 22.2 | 2 | 22.2 | 7 | 77.8 | 9 | 100 |
| Neg | 5 | 5 | 100 | 3 | 60.0 | 4 | 80.0 | 4 | 80.0 | 1 | 20.0 | 1 | 20.0 | 1 | 20.0 | 0 | 0.0 | 1 | 20 | 0 | 0.0 | |
| Total | 14 | 14 | 100 | 9 | 64.3 | 12 | 85.7 | 12 | 85.7 | 7 | 50.0 | 3 | 21.4 | 3 | 21.4 | 2 | 14.3 | 8 | 57.1 | 9 | 64.3 | |
| Dog feces | Pos | 17 | 13 | 76.5 | 7 | 41.2 | 11 | 64.7 | 10 | 58.8 | 10 | 58.8 | 2 | 11.8 | 2 | 11.8 | 3 | 17.6 | 7 | 41.2 | 12 | 70.6 |
| Neg | 12 | 10 | 83.3 | 4 | 33.3 | 6 | 50.0 | 5 | 41.7 | 4 | 33.3 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 8.3 | 0 | 0.0 | |
| Total | 29 | 23 | 79.3 | 11 | 37.9 | 17 | 58.6 | 15 | 51.7 | 14 | 48.3 | 2 | 6.9 | 2 | 6.9 | 3 | 10.3 | 8 | 27.6 | 12 | 41.4 | |
| Flies | Pos | 51 | 51 | 100 | 50 | 98.0 | 50 | 98.0 | 49 | 96.1 | 43 | 84.3 | 20 | 39.2 | 17 | 33.3 | 10 | 19.6 | 24 | 47.1 | 43 | 84.3 |
| Neg | 30 | 26 | 86.7 | 20 | 66.7 | 23 | 76.7 | 13 | 43.3 | 6 | 20 | 1 | 3.3 | 1 | 3.3 | 0 | 0.0 | 0 | 0.0 | 1 | 3.3 | |
| Total | 81 | 77 | 95.1 | 70 | 86.4 | 73 | 90.1 | 62 | 76.5 | 49 | 60.5 | 21 | 25.9 | 18 | 22.2 | 10 | 12.3 | 24 | 29.6 | 44 | 54.3 | |
| Waste- water | Pos | 11 | 10 | 90.9 | 9 | 81.8 | 10 | 90.9 | 10 | 90.9 | 10 | 90.9 | 2 | 18.2 | 1 | 9.1 | 6 | 54.5 | 8 | 72.2 | 9 | 81.1 |
| Neg | 16 | 14 | 87.5 | 13 | 81.2 | 10 | 62.5 | 6 | 37.5 | 7 | 43.8 | 1 | 6.2 | 1 | 6.2 | 3 | 18.8 | 6 | 37.5 | 0 | 0.0 | |
| Total | 27 | 24 | 88.9 | 22 | 81.5 | 20 | 74.0 | 16 | 59.3 | 17 | 63.0 | 3 | 11.1 | 2 | 7.4 | 9 | 33.3 | 14 | 51.9 | 9 | 33.3 | |
| All samples | Pos | 186 | 172 | 92.5 | 139 | 74.7 | 162 | 87.1 | 153 | 82.3 | 147 | 79.0 | 47 | 25.3 | 39 | 21.0 | 34 | 18.3 | 82 | 44.1 | 160 | 86.0 |
| Neg | 150 | 116 | 77.3 | 76 | 50.7 | 96 | 64.0 | 55 | 36.7 | 35 | 23.3 | 6 | 4.0 | 4 | 2.7 | 5 | 3.3 | 12 | 8.0 | 4 | 2.7 | |
| Total | 336 | 288 | 85.7 | 215 | 64.0 | 258 | 76.8 | 208 | 61.9 | 182 | 54.2 | 53 | 15.8 | 43 | 12.8 | 39 | 11.6 | 94 | 28.0 | 164 | 48.8 | |
| Antibiotic Resistance | E. faecalis Harbouring optrA | OR (95% CI) | p | Adjusted OR (95% CI, p) | p | ||||
|---|---|---|---|---|---|---|---|---|---|
| Positive | Negative | ||||||||
| n | % | n | % | ||||||
| Vancomycin | NS | 34 | 64.2 | 19 | 35.8 | 1.5 (0.84–2.83) | 0.16 | - | - |
| S | 152 | 53.7 | 131 | 46.3 | 1 | - | - | ||
| Erythromycin | NS | 177 | 60.8 | 114 | 39.2 | 6.21 (2.88–13.38) | 0.000 | 1.01 (0.19–5.45) | 0.99 |
| S | 9 | 20 | 36 | 80 | 1 | ||||
| Tetracycline | NS | 172 | 59.3 | 118 | 40.7 | 3.33 (1.7–6.51) | 0.000 | 0.01 (0–0.32) | 0.01 |
| S | 14 | 30.4 | 32 | 69.6 | 1 | ||||
| Doxycycline | NS | 173 | 59.7 | 117 | 40.3 | 3.75 (1.89–3.73) | 0.000 | 8.07 (0.41–157.54) | 0.17 |
| S | 13 | 28.3 | 33 | 71.1 | 1 | ||||
| Minocycline | NS | 170 | 60.1 | 113 | 39.9 | 3.48 (1.85–6.55) | 0.000 | 16.99 (0.92–312.10) | 0.06 |
| S | 16 | 30.2 | 37 | 69.8 | 1 | ||||
| Chloramphenicol | NS | 158 | 68.7 | 72 | 31.3 | 6.11 (3.66–10.20) | 0.000 | 2.70 (0.73–9.99) | 0.14 |
| S | 28 | 26.4 | 78 | 73.6 | 1 | ||||
| Ciprofloxacin | NS | 112 | 64.4 | 62 | 35.6 | 2.15 (1.39–2.33) | 0.000 | 0.53 (0.15–1.92) | 0.33 |
| S | 74 | 45.7 | 88 | 54.3 | 1 | ||||
| Levofloxacin | NS | 64 | 76.2 | 20 | 23.8 | 3.40 (1.95–5.97) | 0.000 | 5.37 (1.29–22.38) | 0.02 |
| S | 122 | 48.4 | 130 | 51.6 | 1 | ||||
| HLGR | NS | 34 | 87.2 | 5 | 12.8 | 6.49 (2.47–17.04) | 0.000 | 1.60 (0.22–11.71) | 0.64 |
| S | 152 | 51.2 | 145 | 48.8 | 1 | ||||
| HLSR | NS | 82 | 87.2 | 12 | 12.8 | 9.07 (4.70–14.49) | 0.000 | 1.67 (0.33–8.43) | 0.54 |
| S | 104 | 43 | 138 | 57 | 1 | ||||
| Linezolid | NS | 172 | 97.7 | 4 | 2.3 | 448 (144–1392) | 0.000 | 540 (134–2175) | 0.000 |
| S | 14 | 8.8 | 146 | 91.2 | 1 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha, H.T.A.; Nguyen, P.T.L.; Hung, T.T.M.; Tuan, L.A.; Thuy, B.T.; Lien, T.H.M.; Thai, P.D.; Thanh, N.H.; Bich, V.T.N.; Anh, T.H.; et al. Prevalence and Associated Factors of optrA-Positive-Enterococcus faecalis in Different Reservoirs around Farms in Vietnam. Antibiotics 2023, 12, 954. https://doi.org/10.3390/antibiotics12060954
Ha HTA, Nguyen PTL, Hung TTM, Tuan LA, Thuy BT, Lien THM, Thai PD, Thanh NH, Bich VTN, Anh TH, et al. Prevalence and Associated Factors of optrA-Positive-Enterococcus faecalis in Different Reservoirs around Farms in Vietnam. Antibiotics. 2023; 12(6):954. https://doi.org/10.3390/antibiotics12060954
Chicago/Turabian StyleHa, Hoang Thi An, Phuong Thi Lan Nguyen, Tran Thi Mai Hung, Le Anh Tuan, Bui Thanh Thuy, Tran Hoang My Lien, Pham Duy Thai, Nguyen Ha Thanh, Vu Thi Ngoc Bich, Tran Hai Anh, and et al. 2023. "Prevalence and Associated Factors of optrA-Positive-Enterococcus faecalis in Different Reservoirs around Farms in Vietnam" Antibiotics 12, no. 6: 954. https://doi.org/10.3390/antibiotics12060954
APA StyleHa, H. T. A., Nguyen, P. T. L., Hung, T. T. M., Tuan, L. A., Thuy, B. T., Lien, T. H. M., Thai, P. D., Thanh, N. H., Bich, V. T. N., Anh, T. H., Hanh, N. T. H., Minh, N. T., Thanh, D. P., Mai, S.-N. T., The, H. C., Trung, N. V., Thu, N. H., Duong, T. N., Anh, D. D., ... Hoang, T. H. (2023). Prevalence and Associated Factors of optrA-Positive-Enterococcus faecalis in Different Reservoirs around Farms in Vietnam. Antibiotics, 12(6), 954. https://doi.org/10.3390/antibiotics12060954