
Effect of Phenylalanine–Arginine Beta-Naphthylamide on the Values of Minimum Inhibitory Concentration of Quinolones and Aminoglycosides in Clinical Isolates of Acinetobacter baumannii
 , , ,
, , ,
Abstract
1. Introduction
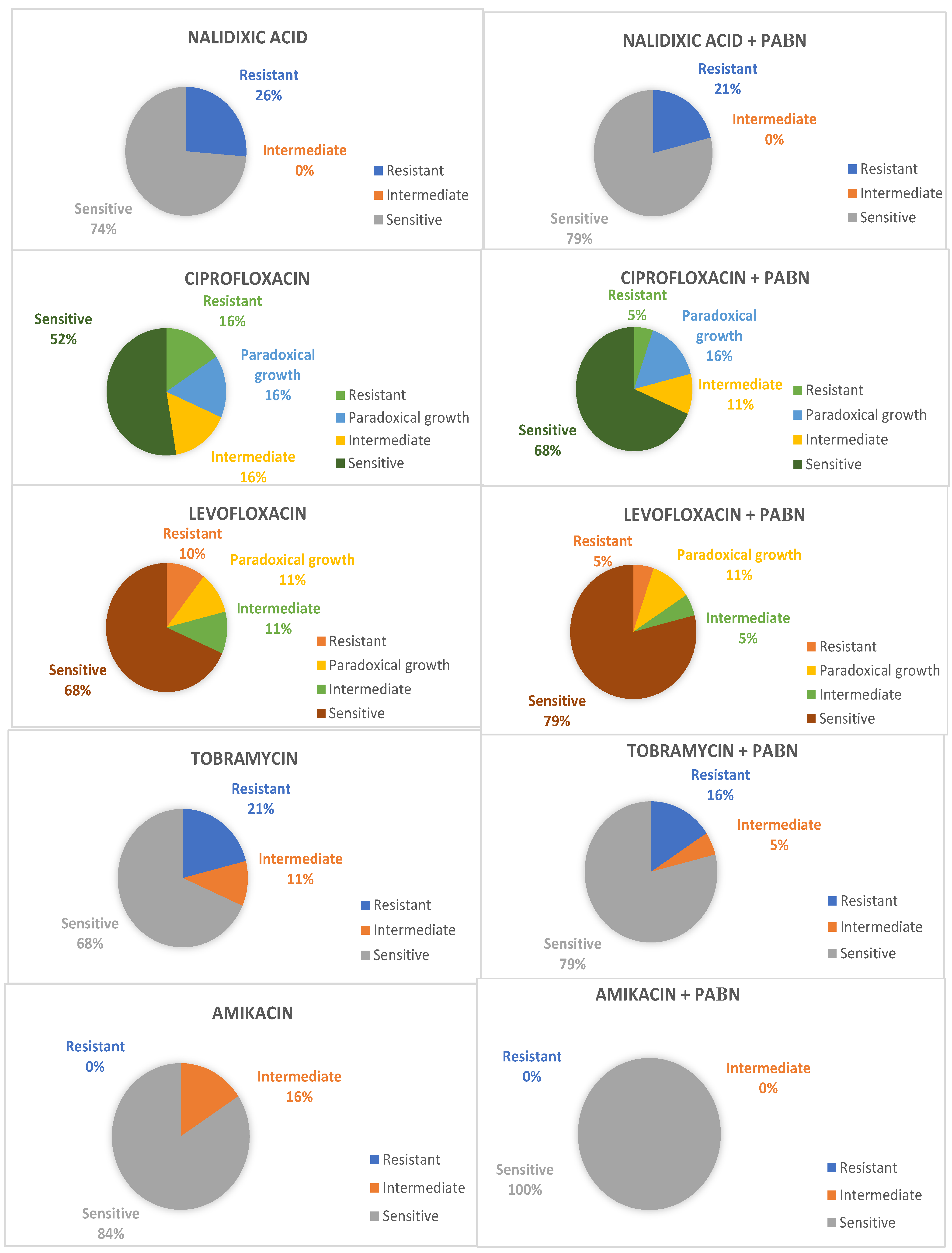
2. Results
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Bacterial Culture Conditions and Identification
4.3. Antimicrobial Susceptibility Testing
4.4. Phenylalanine–Arginine beta-Naphthylamide (PAβN)
4.5. Ethics Statement
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karageorgopoulos, D.E.; Falagas, M.E. Current control and treatment of multidrug-resistant Acinetobacter baumannii infections. Lancet Infect. Dis. 2008, 8, 751–762. [Google Scholar] [CrossRef]
- Nemec, A.; Krizova, L.; Maixnerova, M.; van der Reijden, T.J.; Deschaght, P.; Passet, V.; Vaneechoutte, M.; Brisse, S.; Dijkshoorn, L. Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov. (formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov. (formerly Acinetobacter genomic species 13TU). Res. Microbiol. 2011, 162, 393–404. [Google Scholar]
- Vijayakumar, S.; Biswas, I.; Veeraraghavan, B. Accurate identification of clinically important Acinetobacter spp.: An update. Future Sci. OA 2019, 5, FSO395. [Google Scholar] [CrossRef]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef]
- Maragakis, L.L.; Perl, T.M. Acinetobacter baumannii: Epidemiology, antimicrobial resistance, and treatment options. Clin. Infect. Dis. 2008, 46, 1254–1263. [Google Scholar] [CrossRef]
- Park, S.; Lee, K.M.; Yoo, Y.S.; Yoo, J.S.; Yoo, J.I.; Kim, H.S.; Lee, Y.S.; Chung, G.T. Alterations of gyrA, gyrB, and parC and Activity of Efflux Pump in Fluoroquinolone-resistant Acinetobacter baumannii. Osong Public Health Res. Perspect. 2011, 2, 164–170. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef]
- Chittawatanarat, K.; Jaipakdee, W.; Chotirosniramit, N.; Chandacham, K.; Jirapongcharoenlap, T. Microbiology, resistance patterns, and risk factors of mortality in ventilator-associated bacterial pneumonia in a Northern Thai tertiary-care university based general surgical intensive care unit. Infect. Drug. Resist. 2014, 7, 203–210. [Google Scholar] [CrossRef]
- Cisneros, J.M.; Rodriguez-Bano, J.; Fernandez-Cuenca, F.; Ribera, A.; Vila, J.; Pascual, A.; Martínez-Martínez, L.; Bou, G.; Pachón, J.; on behalf of the Spanish Group for Nosocomial Infection (GEIH) for the Spanish Society of Infectious Diseases and Clinical Microbiology (SEIMC). Risk-factors for the acquisition of imipenem-resistant Acinetobacter baumannii in Spain: A nationwide study. Clinical microbiology and infection: The official publication of the European Society of Clinical Microbiology and Infectious Diseases. Clin. Microbiol. Infect. 2005, 11, 874–879. [Google Scholar] [CrossRef]
- Falagas, M.E.; Kopterides, P. Risk factors for the isolation of multi-drug-resistant Acinetobacter baumannii and Pseudomonas aeruginosa: A systematic review of the literature. J. Hosp. Infect. 2006, 64, 7–15. [Google Scholar] [CrossRef]
- Dijkshoorn, L.; Nemec, A.; Seifert, H. An increasing threat in hospitals: Multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 2007, 5, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.; O’Donoghue, M.; Feeney, A.; Sleator, R.D. Acinetobacter baumannii: An emerging opportunistic pathogen. Virulence 2012, 3, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Price, L.S.; Weinstein, R.A. Acinetobacter infection. N. Engl. J. Med. 2008, 358, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Bliziotis, I.A.; Siempos, I.I. Attributable mortality of Acinetobacter baumannii infections in critically ill patients: A systematic review of matched cohort and case-control studies. Crit. Care. 2006, 10, R48. [Google Scholar] [CrossRef] [PubMed]
- Shin, B.; Park, W. Antibiotic resistance of pathogenic Acinetobacter species and emerging combination therapy. J. Microbiol. 2017, 55, 837–849. [Google Scholar] [CrossRef]
- Ayoub Moubareck, C.; Hammoudi Halat, D. Insights into Acinetobacter baumannii: A Review of Microbiological, Virulence, and Resistance Traits in a Threatening Nosocomial Pathogen. Antibiotics 2020, 9, 119. [Google Scholar] [CrossRef]
- Giammanco, A.; Calà, C.; Fasciana, T.; Dowzicky, M.J. Global Assessment of the Activity of Tigecycline against Multidrug-Resistant Gram-Negative Pathogens between 2004 and 2014 as Part of the Tigecycline Evaluation and Surveillance Trial. MSphere 2017, 2, e00310-16. [Google Scholar] [CrossRef]
- Rodríguez, C.H.; Nastro, M.; Famiglietti, A. Carbapenemases in Acinetobacter baumannii. Review of their dissemination in Latin America. Rev. Argent. Microbiol. 2018, 50, 327–333. [Google Scholar] [CrossRef]
- Zhao, Y.; Hu, K.; Zhang, J.; Guo, Y.; Fan, X.; Wang, Y.; Mensah, S.D.; Zhang, X. Outbreak of carbapenem-resistant Acinetobacter baumannii carrying the carbapenemase OXA-23 in ICU of the eastern Heilongjiang Province, China. BMC Infect. Dis. 2019, 19, 452. [Google Scholar]
- Du, D.; Wang-Kan, X.; Neuberger, A.; van Veen, H.W.; Pos, K.M.; Piddock, L.J.V.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef]
- Roy, S.; Junghare, V.; Dutta, S.; Hazra, S.; Basu, S. Differential Binding of Carbapenems with the AdeABC Efflux Pump and Modulation of the Expression of AdeB Linked to Novel Mutations within Two-Component System AdeRS in Carbapenem-Resistant Acinetobacter baumannii. MSystems 2022, 7, e0021722. [Google Scholar] [CrossRef]
- Kaynak Onurdağ, F.; Kayış, U.; Ökten, S. Acinetobacter baumannii İzolatlarında Fenilalanin-arjinin-beta-naftilamidin Siprofloksasinin Minimal İnhibitör Konsantrasyon Değerleri ve Dışa Atım Pompası Genlerinin Ekspresyonu Üzerine Etkisi [Effect of Phenylalanine-arginine-beta-naphthylamide to Ciprofloxacin Minimum Inhibitory Concentration Values and Expression of Efflux Pump System Genes in Acinetobacter baumannii Isolates]. Mikrobiyol. Bülteni 2021, 55, 285–299. (In Turkish) [Google Scholar]
- Abdi, S.N.; Ghotaslou, R.; Ganbarov, K.; Mobed, A.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Kafil, H.S. Acinetobacter baumannii Efflux Pumps and Antibiotic Resistance. Infect. Drug. Resist. 2020, 13, 423–434. [Google Scholar] [CrossRef]
- Hassan, K.A.; Liu, Q.; Henderson, P.J.; Paulsen, I.T. Homologs of the Acinetobacter baumannii AceI transporter represent a new family of bacterial multidrug efflux systems. MBio 2015, 6, e01982-14. [Google Scholar] [CrossRef]
- Yoon, E.J.; Courvalin, P.; Grillot-Courvalin, C. RND-type efflux pumps in multidrug-resistant clinical isolates of Acinetobacter baumannii: Major role for AdeABC overexpression and AdeRS mutations. Antimicrob. Agents Chemother. 2013, 57, 2989–2995. [Google Scholar] [CrossRef]
- Abd El-Rahman, O.A.; Rasslan, F.; Hassan, S.S.; Ashour, H.M.; Wasfi, R. The RND Efflux Pump Gene Expression in the Biofilm Formation of Acinetobacter baumannii. Antibiotics 2023, 12, 419. [Google Scholar] [CrossRef]
- Coyne, S.; Courvalin, P.; Perichon, B. Efflux-mediated antibiotic resistance in Acinetobacter spp. Antimicrob Agents Chemother. 2011, 55, 947–953. [Google Scholar] [CrossRef]
- Ardebili, A.; Talebi, M.; Azimi, L.; Lari, A.R. Effect of Efflux Pump Inhibitor Carbonyl Cyanide 3-Chlorophenylhydrazone on the Minimum Inhibitory Concentration of Ciprofloxacin in Acinetobacter baumannii Clinical Isolates. Jundishapur. J. Microbiol. 2014, 7, e8691. [Google Scholar] [CrossRef]
- Park, Y.K.; Ko, K.S. Effect of carbonyl cyanide 3-chlorophenylhydrazone (CCCP) on killing Acinetobacter baumannii by colistin. J. Microbiol. 2015, 53, 53–59. [Google Scholar] [CrossRef]
- Yang, Y.; Chua, K.L. Assessment of the effect of efflux pump inhibitors on in vitro antimicrobial susceptibility of multidrug-resistant Acinetobacter baumannii. Int. J. Antimicrob. Agents. 2013, 42, 283–284. [Google Scholar] [CrossRef]
- Pannek, S.; Higgins, P.G.; Steinke, P.; Jonas, D.; Akova, M.; Bohnert, J.A.; Seifert, H.; Kern, W.V. Multidrug efflux inhibition in Acinetobacter baumannii: Comparison between 1-(1-naphthylmethyl)-piperazine and phenyl-arginine-beta-naphthylamide. J. Antimicrob. Chemother. 2006, 57, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Bowers, D.R.; Cao, H.; Zhou, J.; Ledesma, K.R.; Sun, D.; Lomovskaya, O.; Tam, V.H. Assessment of minocycline and polymyxin B combination against Acinetobacter baumannii. Antimicrob. Agents Chemother. 2015, 59, 2720–2725. [Google Scholar] [CrossRef] [PubMed]
- Gholami, M.; Hashemi, A.; Hakemi-Vala, M.; Goudarzi, H.; Hallajzadeh, M. Efflux Pump Inhibitor Phenylalanine-Arginine Beta-Naphthylamide Effect on the Minimum Inhibitory Concentration of Imipenem in Acinetobacter baumannii Strains Isolated From Hospitalized Patients in Shahid Motahari Burn Hospital, Tehran, Iran. Jundishapur. J. Microbiol. 2015, 8, e19048. [Google Scholar] [CrossRef] [PubMed]
- Lamers, R.P.; Cavallari, J.F.; Burrows, L.L. The efflux inhibitor phenylalanine-arginine beta-naphthylamide (PAbetaN) permeabilizes the outer membrane of gram-negative bacteria. PLoS ONE 2013, 8, e60666. [Google Scholar] [CrossRef]
- CLSI. CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018; ISBN 1-56238-839-8. [Google Scholar]
- Ruppé, É.; Woerther, P.L.; Barbier, F. Mechanisms of antimicrobial resistance in Gram-negative bacilli. Ann. Intensive Care. 2015, 5, 61. [Google Scholar] [CrossRef]
- Gautam, D.; Dolma, K.G.; Khandelwal, B.; Mitsuwan, W.; Mahboob, T.; Pereira, M.L.; Nawaz, M.; Wiart, C.; Ardebili, A.; Siyadatpanah, A.; et al. Acinetobacter baumannii: An overview of emerging multidrug-resistant pathogen. Med. J. Malaysia. 2022, 77, 357–370. [Google Scholar]
- Sharma, S.; Kaushik, V.; Kulshrestha, M.; Tiwari, V. Different Efflux Pump Systems in Acinetobacter baumannii and Their Role in Multidrug Resistance. Adv. Exp. Med. Biol. 2023, 1370, 155–168. [Google Scholar]
- Meyer, C.; Lucaβen, K.; Gerson, S.; Xanthopoulou, K.; Wille, T.; Seifert, H.; Higgins, P.G. Contribution of RND-Type Efflux Pumps in Reduced Susceptibility to Biocides in Acinetobacter baumannii. Antibiotics 2022, 11, 1635. [Google Scholar] [CrossRef]
- Mittal, K.R.; Jain, N.; Srivastava, P.; Jain, C.K. Multidrug-Resistant Acinetobacter baumannii: An Emerging Aspect of New Drug Discovery. Recent. Adv. Antiinfect. Drug. Discov. 2023, 18, 29–41. [Google Scholar]
- Li, X.-Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef]
- Cortez-Cordova, J.; Kumar, A. Activity of the efflux pump inhibitor phenylalanine-arginine β-naphthylamide against the AdeFGH pump of Acinetobacter baumannii. Int. J. Antimicrob. Agents. 2011, 37, 420–424. [Google Scholar] [CrossRef]
- Magnet, S.; Courvalin, P.; Lambert, T. Resistance–nodulation–cell division-type efflux pump involved in aminoglycoside resistance in Acinetobacter baumannii strain BM4454. Antimicrob. Agents Chemother. 2001, 45, 3375–3380. [Google Scholar] [CrossRef]
- Damier-Piolle, L.; Magnet, S.; Bremont, S.; Lambert, T.; Courvalin, P. AdeIJK, a resistance–nodulation–cell division pump effluxing multiple antibiotics in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2008, 52, 557–562. [Google Scholar] [CrossRef]
- Coyne, S.; Rosenfeld, N.; Lambert, T.; Courvalin, P.; Perichon, B. Overexpression of resistance–nodulation–cell division pump AdeFGH confers multidrug resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 4389–4393. [Google Scholar] [CrossRef]
- Zheng, W.; Huang, Y.; Wu, W.; Zhu, J.; Zhang, T. Analysis of Efflux Pump System and Other Drug Resistance Related Gene Mutations in Tigecycline-Resistant Acinetobacter baumannii. Comput. Math. Methods Med. 2023, 2023, 8611542. [Google Scholar] [CrossRef]
- Chau, S.L.; Chu, Y.W.; Houang, E.T. Novel resistance–nodulation–cell division efflux system AdeDE in Acinetobacter genomic DNA group 3. Antimicrob. Agents Chemother. 2004, 48, 4054–4055. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Potoski, B.A.; Rea, R.; Adams, J.; Sethi, J.; Capitano, B.; Husain, S.; Kwak, E.J.; Bhat, S.V.; Paterson, D.L. Acinetobacter baumannii bloodstream infection while receiving tigecycline: A cautionary report. J. Antimicrob. Chemother. 2007, 59, 128–131. [Google Scholar] [CrossRef]
- Valentine, S.C.; Contreras, D.; Tan, S.; Real, L.J.; Chu, S.; Xu, H.H. Phenotypic and molecular characterization of Acinetobacter baumannii clinical isolates from nosocomial outbreaks in Los Angeles County, California. J. Clin. Microbiol. 2008, 46, 2499–2507. [Google Scholar] [CrossRef]
- Khayat, H.; Sadeghifard, N.; Pakzad, I.; Azimi, L.; Delfani, S.; Sayehmiri, K.; Soroush, S.; Bogdanovic, L.; Taherikalani, M. Determination of Different Fluoroquinolone Mechanisms Among Clinical Isolates of Acinetobacter baumannii in Tehran, Iran. Iran. Red. Crescent. Med. J. 2017, 19, e58798.37. [Google Scholar] [CrossRef]
- Golanbar, G.D.; Lam, C.K.; Chu, Y.M.; Cueva, C.; Tan, S.W.; Silva, I.; Xu, H.H. Phenotypic and molecular characterization of Acinetobacter clinical isolates obtained from inmates of California correctional facilities. J. Clin. Microbiol. 2011, 49, 2121–2131. [Google Scholar] [CrossRef]
- Ribera, A.; Ruiz, J.; Jiminez de Anta, M.T.; Vila, J. Effect of an efflux pump inhibitor on the MIC of nalidixic acid for Acinetobacter baumannii and Stenotrophomonas maltophilia clinical isolates. J. Antimicrob. Chemother. 2002, 49, 697–698. [Google Scholar] [CrossRef] [PubMed]
- Laudy, A.E.; Mrowka, A.; Krajewska, J.; Tyski, S. The Influence of Efflux Pump Inhibitors on the Activity of Non-Antibiotic NSAIDS against Gram-Negative Rods. PLoS ONE 2016, 11, e0147131. [Google Scholar] [CrossRef] [PubMed]
- Coban, A.Y.; Guney, A.K.; Tanriverdi Cayci, Y.; Durupinar, B. Effect of 1-(1-Naphtylmethyl)-piperazine, an efflux pump inhibitor, on antimicrobial drug susceptibilities of clinical Acinetobacter baumannii isolates. Curr. Microbiol. 2011, 62, 508–511. [Google Scholar] [CrossRef] [PubMed]
- Cheng, V.C.; Chen, J.H.; So, S.Y.; Wong, S.C.; Yan, M.K.; Chau, P.H.; Lee, W.M.; To, K.K.W.; Chan, J.F.W.; Hung, I.F.N.; et al. Use of fluoroquinolones is the single most important risk factor for the high bacterial load in patients with nasal and gastrointestinal colonization by multidrug-resistant Acinetobacter baumannii. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 2359–2366. [Google Scholar] [CrossRef]
- Levy-Blitchtein, S.; Roca, I.; Plasencia-Rebata, S.; Vicente-Taboada, W.; Velasquez-Pomar, J.; Munoz, L.; Moreno-Morales, J.; Pons, M.J.; del Valle-Mendoza, J.; Vila, J. Emergence and spread of carbapenem-resistant Acinetobacter baumannii international clones II and III in Lima, Peru. Emerg. Microbes Infect. 2018, 7, 119. [Google Scholar] [CrossRef]
- Yamane, K.; Wachino, J.; Doi, Y.; Kurokawa, H.; Arakawa, Y. Global spread of multiple aminoglycoside resist genes. Emerg. Infect. Dis. 2005, 11, 951–953. [Google Scholar] [CrossRef]
- Xu, C.; Bilya, S.R.; Xu, W. adeABC efflux gene in Acinetobacter baumannii. New Microbes New Infect. 2019, 30, 100549. [Google Scholar] [CrossRef]
- Appleman, M.D.; Belzberg, H.; Citron, D.M.; Heseltine, P.N.; Yellin, A.E.; Murray, J.; Berne, T.V. In vitro activities of nontraditional antimicrobials against multiresistant Acinetobacter baumannii strains isolated in an intensive care unit outbreak. Antimicrob. Agents Chemother. 2000, 44, 1035–1040. [Google Scholar] [CrossRef]
- Ferrer-Espada, R.; Shahrour, H.; Pitts, B.; Stewart, P.S.; Sánchez-Gómez, S.; Martínez-de-Tejada, G. A permeability-increasing drug synergizes with bacterial efflux pump inhibitors and restores susceptibility to antibiotics in multi-drug resistant Pseudomonas aeruginosa strains. Sci. Rep. 2019, 9, 3452. [Google Scholar] [CrossRef]
- Prasetyoputri, A.; Jarrad, A.M.; Cooper, M.A.; Blaskovich, M.A.T. The Eagle Effect and Antibiotic-Induced Persistence: Two Sides of the Same Coin? Trends. Microbiol. 2019, 27, 339–354. [Google Scholar] [CrossRef]
- Leão, P.V.S.; Ferreira, A.L.D.S.; Oliveira, F.A.A.; Mesquita, A.B.S.; Lima-Net, J.S.; Gutierrez, S.J.C.; Nogueira, C.E.S.; Cruz-Martins, N.; Arcanjo, D.D.R.; Barreto, H.M.; et al. Riparin-B as a Potential Inhibitor of AdeABC Efflux System from Acinetobacter baumannii. Evid.-Based Complement. Altern. Med. 2023, 2023, 1780838. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef]
- Oh, S.Y.; Han, J.Y.; Lee, S.R.; Lee, H.T. Improved DNA Extraction Method for Molecular Diagnosis from Smaller numbers of Cells. Korean J. Clin. Lab. Sci. 2014, 46, 99–105. [Google Scholar] [CrossRef]
- Turton, J.F.; Woodford, N.; Glover, J.; Yarde, S.; Kaufmann, M.E.; Pitt, T.L. Identification of Acinetobacter baumannii by detection of the blaOXA-51-like carbapenemase gene intrinsic to this species. J. Clin. Microbiol. 2006, 44, 2974–2976. [Google Scholar] [CrossRef]
- Liu, Y.H.; Kuo, S.C.; Lee, Y.T.; Chang, I.C.; Yang, S.P.; Chen, T.L.; Fung, C.P. Amino acid substitutions of quinolone resistance determining regions in GyrA and ParC associated with quinolone resistance in Acinetobacter baumannii and Acinetobacter genomic species 13TU. J. Microbiol. Immunol. Infect. 2012, 45, 108–112. [Google Scholar] [CrossRef]


{kind=link}
| Strain | MIC (µg/mL) (Susceptibility Rating) for Indicated Drug | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Antibiotic | Antibiotic + PaβN | |||||||||
| TOB | CIP | LEV | NA | AK | TOB | CIP | LEV | NA | AK | |
| Ac2 | 0.5 (S) | 0.5 (S) | 8 (R) | 128 (R) | <0.25 (S) | 1 (S) | 0.25 (S) | 2 (S) i | 32 (R) | 2 (S) |
| Ac10 | 1 (S) | 0.5 (S) | 32 (R) | 32 (R) | <0.25 (S) | 2 (S) | 1 (S) | 32 (R) | 32 (R) | 1 (S) |
| Ac20 | 8 (I) | 2 (I) | 1 (S) | 1 (S) | <0.25 (S) | 4 (S) | 0.5 (S) i | 4 (I) | 4 (S) | <0.25 (S) |
| Ac21 | 8 (I) | <0.25 (S) | <0.25 (S) | 1 (S) | 8 (S) | 4 (S) | 32 (R) | 0.5 (S) | 2 (S) | 4 (S) |
| Ac22 | 0.5 (S) | 0.25 (S) | 2 (S) | 16 (S) | 4 (S) | 0.25 (S) | 0.5 (S) | 4 (I) | 1 (S) | 2 (S) |
| Ac23 | 64 (R) | 8 (R) | 0.25 (S) | 0.5 (S) | 16 (S) | 64 (R) | 2 (I) i | 0.25 (S) | 2 (S) | 4 (S) |
| Ac24 | 0.5 (S) | 2 (I) | 0.5 (S) | 1 (S) | 32 (I) | 1 (S) | 0.25 (S) i | 0.25 (S) | 0.5 (S) | 16 (S) |
| Ac25 | 32 (R) | 0.5 (S) | 1 (S) | 0.25 (S) | 32 (I) | 16 (R) | 0.25 (S) | 0.25 (S) | 0.5 (S) | 4 (S) i |
| Ac26 | 0.25 (S) | 8 (R) | 0.5 (S) | 64 (R) | <0.25 (S) | 2 (S) | 16 (R) | 1 (S) | 32 (R) | <0.25 (S) |
| Ac27 | 0.5 (S) | 1 (S) | 0.25 (S) | 64 (R) | 16 (S) | 4 (S) | 2 (I) | 0.25 (S) | 8 (S)i | 4 (S) |
| Ac28 | 0.25 (S) | 4 (R) | 2 (S) | 16 (S) | <0.25 (S) | 4 (S) | <0.25 (S) i | 0.5 (S) | 16 (S) | 1 (S) |
| Ac29 | 0.5 (S) | 2 (I) | 1 (S) | 64 (R) | 32 (I) | 4 (S) | 2 (I) | 1 (S) | 32 (R) | 4 (S) i |
| Ac30 | 2 (S) | 0.5 (S) | 1 (S) | 2 (S) | 0.5 (S) | 0.5 (S) | 0.25 (S) | <0.25 (S) | 0.5 (S) | 0.25 (S) |
| Ac39 | <0.25 (S) | <0.25 (S) | 2 (S) | 2 (S) | 1 (S) | <0.25 (S) | <0.25 (S) | 0.5 (S) | 0.25 (S) | 0.5 (S) |
| Ac41 | 16 (R) | <0.25 (S) | 1 (S) | 4 (S) | 2 (S) | 16 (R) | <0.25 (S) | 2 (S) | 2 (S) | <0.25 (S) |
| Ac46 | 16 (R) | <0.25 (S) | 0.25 (S) | 4 (S) | <0.25 (S) | 8 (I) | 0.25 (S) | <0.25 (S) | 1 (S) | 0.25 (S) |
| Ac50 | 1 (S) | <0.25 (S) | <0.25 (S) | 0.25 (S) | 16 (S) | 0.5 (S) | 4 (R) | <0.25 (S) | 0.25 (S) | <0.25 (S) |
| Ac54 | 4 (S) | 1 (S) | 4 (I) | 2 (S) | <0.25 (S) | 1 (S) | 1 (S) | 2 (S) | 0.25 (S) | 4 (S) |
| Ac55 | 4 (S) | 0.5 (S) | 4 (I) | 2 (S) | 1 (S) | 4 (S) | 0.5 (S) | 4 (I) | 2 (S) | <0.25 (S) |
| Antimicrobial Agent | Disk Content | Interpretive Categories and MIC Breakpoints (µg/mL) | ||
|---|---|---|---|---|
| S | I | R | ||
| Tobramycin | 10 µg | ≤4 | 8 | ≥16 |
| Amikacin | 30 µg | ≤16 | 32 | ≥64 |
| Ciprofloxacin | 5 µg | ≤1 | 2 | ≥4 |
| Levofloxacin | 5 µg | ≤2 | 4 | ≥8 |
| Nalidixic acid | 30 µg | ≤16 | - | ≥32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plasencia-Rebata, S.; Levy-Blitchtein, S.; del Valle-Mendoza, J.; Silva-Caso, W.; Peña-Tuesta, I.; Vicente Taboada, W.; Barreda Bolaños, F.; Aguilar-Luis, M.A. Effect of Phenylalanine–Arginine Beta-Naphthylamide on the Values of Minimum Inhibitory Concentration of Quinolones and Aminoglycosides in Clinical Isolates of Acinetobacter baumannii. Antibiotics 2023, 12, 1071. https://doi.org/10.3390/antibiotics12061071
Plasencia-Rebata S, Levy-Blitchtein S, del Valle-Mendoza J, Silva-Caso W, Peña-Tuesta I, Vicente Taboada W, Barreda Bolaños F, Aguilar-Luis MA. Effect of Phenylalanine–Arginine Beta-Naphthylamide on the Values of Minimum Inhibitory Concentration of Quinolones and Aminoglycosides in Clinical Isolates of Acinetobacter baumannii. Antibiotics. 2023; 12(6):1071. https://doi.org/10.3390/antibiotics12061071
Chicago/Turabian StylePlasencia-Rebata, Stefany, Saul Levy-Blitchtein, Juana del Valle-Mendoza, Wilmer Silva-Caso, Isaac Peña-Tuesta, William Vicente Taboada, Fernando Barreda Bolaños, and Miguel Angel Aguilar-Luis. 2023. "Effect of Phenylalanine–Arginine Beta-Naphthylamide on the Values of Minimum Inhibitory Concentration of Quinolones and Aminoglycosides in Clinical Isolates of Acinetobacter baumannii" Antibiotics 12, no. 6: 1071. https://doi.org/10.3390/antibiotics12061071
APA StylePlasencia-Rebata, S., Levy-Blitchtein, S., del Valle-Mendoza, J., Silva-Caso, W., Peña-Tuesta, I., Vicente Taboada, W., Barreda Bolaños, F., & Aguilar-Luis, M. A. (2023). Effect of Phenylalanine–Arginine Beta-Naphthylamide on the Values of Minimum Inhibitory Concentration of Quinolones and Aminoglycosides in Clinical Isolates of Acinetobacter baumannii. Antibiotics, 12(6), 1071. https://doi.org/10.3390/antibiotics12061071