
Evaluation of Aztreonam and Ceftazidime/Avibactam Synergism against Klebsiella pneumoniae by MALDI-TOF MS
 , and
, and
Abstract
1. Introduction
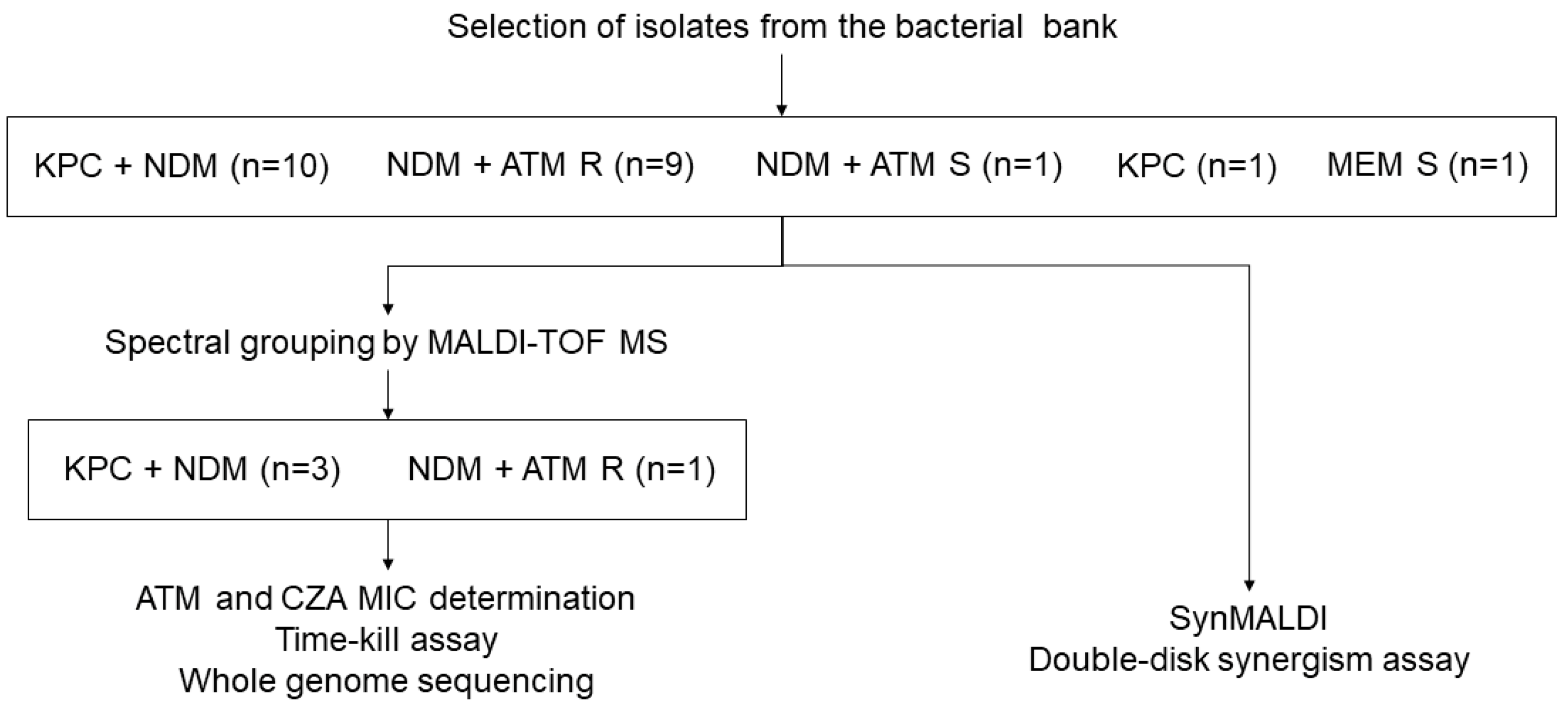
2. Materials and Methods
2.1. Bacterial Strains
2.2. Determination of Susceptibility by Disk Diffusion and Minimum Inhibitory Concentration (MIC)
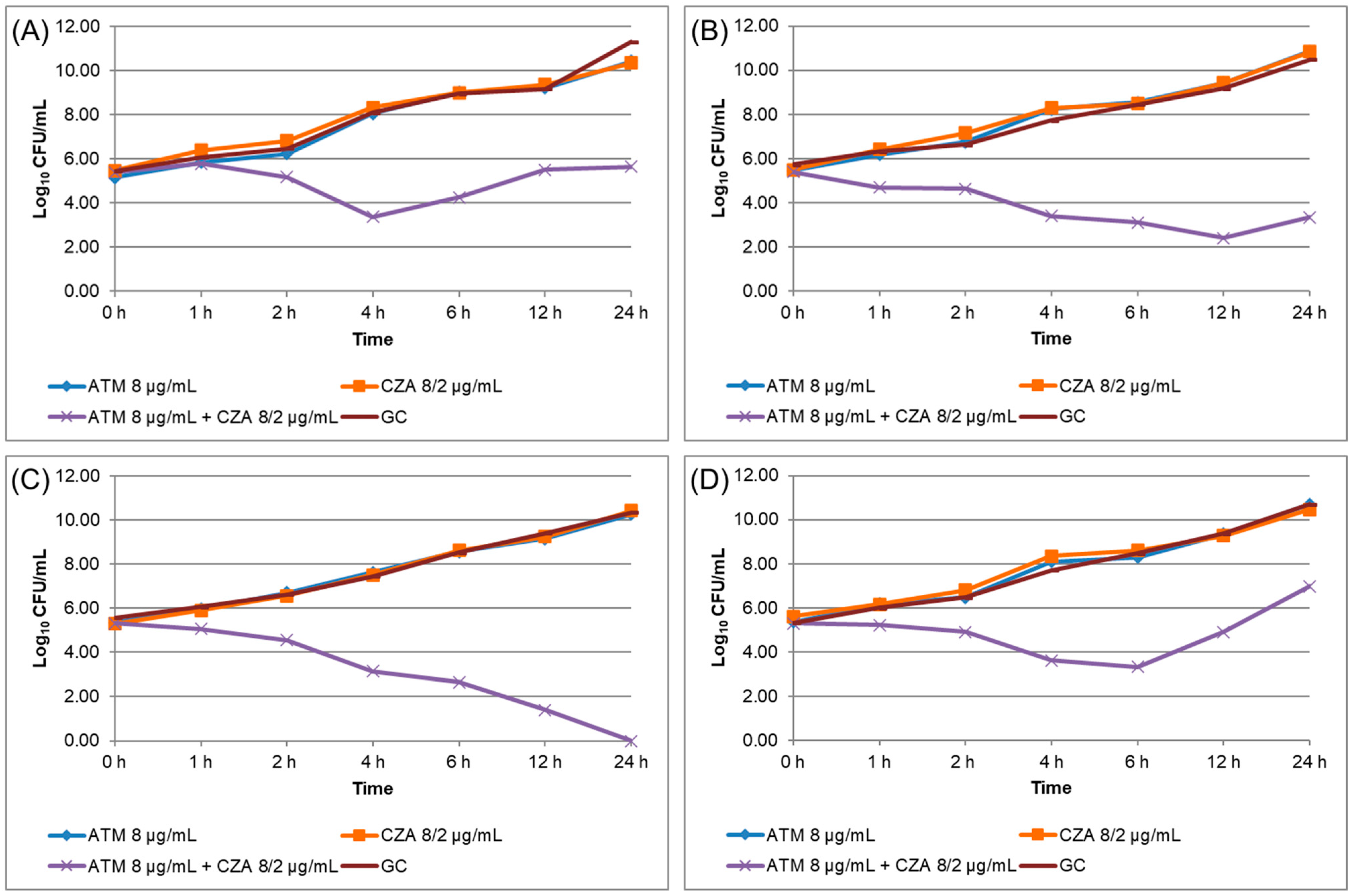
2.3. Time–Kill Curve (TKC) Assay
2.4. Whole Genome Sequencing (WGS)
2.5. Antibiotic Solution Preparation for Synergism by MALDI-TOF MS (SynMALDI)
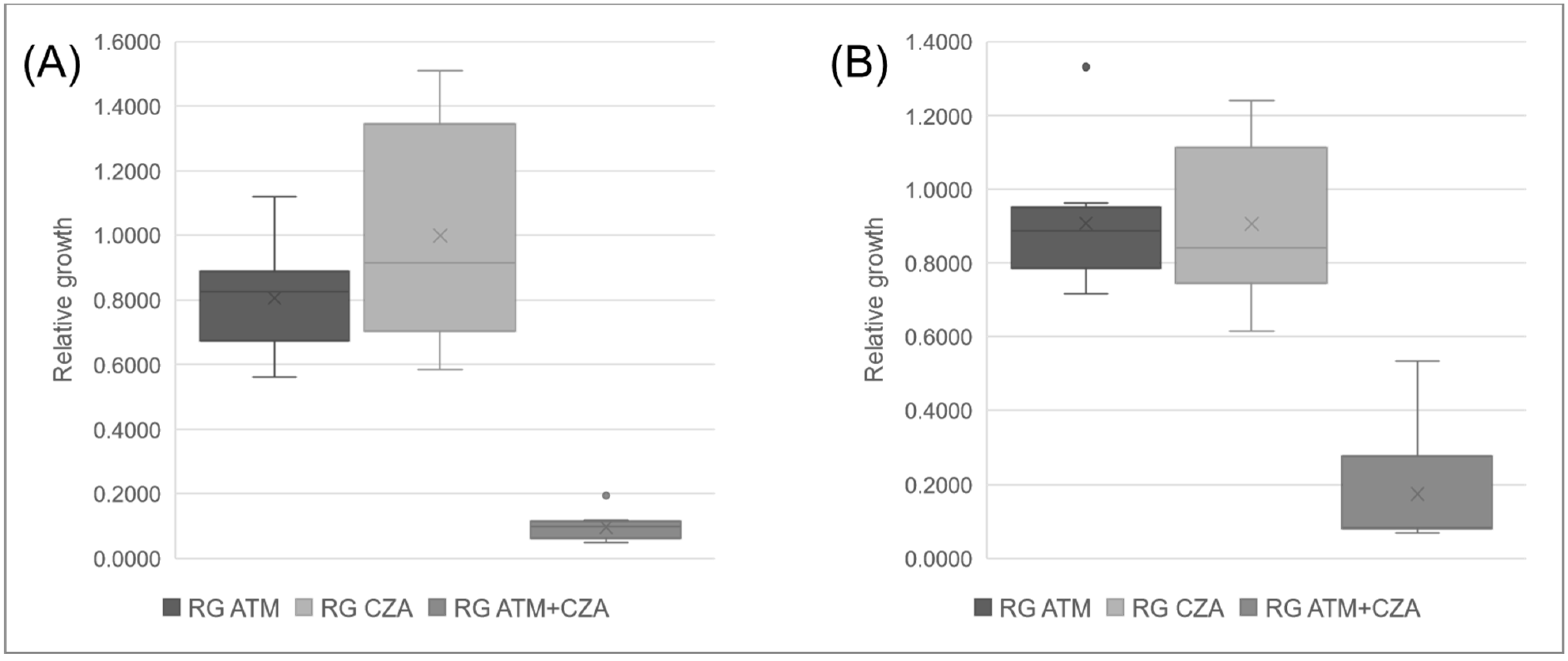
2.6. Synergism by MALDI-TOF MS (SynMALDI)
2.7. Double-Disk Synergism Assay
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CDC Antibiotic Resistance Threats in the United States. 2019. Available online: www.cdc.gov (accessed on 9 July 2022).
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery and Development of New Antibiotics; WHO: Rome, Italy, 2017; pp. 1–7. [Google Scholar]
- Coleman, K. Diazabicyclooctanes (DBOs): A potent new class of non-β-lactam β-lactamase inhibitors. Curr. Opin. Microbiol. 2011, 14, 550–555. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. B-lactams and B-lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef] [PubMed]
- Algammal, A.; Hetta, H.F.; Mabrok, M.; Behzadi, P. Editorial: Emerging multidrug-resistant bacterial pathogens “‘superbugs’”: A rising public health threat. Front. Microbiol. 2023, 14, 1135614. [Google Scholar] [CrossRef]
- Karampatakis, T.; Tsergouli, K.; Behzadi, P. Carbapenem-resistant Klebsiella pneumoniae: Virulence factors, molecular mpidemiology and latest updates in treatment options. Antibiotics 2023, 12, 234. [Google Scholar] [CrossRef]
- Castanheira, M.; Deshpande, L.M.; Mendes, R.E.; Canton, R.; Sader, H.S.; Jones, R.N. Variations in the occurrence of resistance phenotypes and carbapenemase genes among enterobacteriaceae isolates in 20 years of the SENTRY antimicrobial surveillance program. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2019; Volume 6, pp. S23–S33. [Google Scholar]
- Bush, K.; Bradford, P.A. Epidemiology of β-lactamase-producing pathogens. Clin. Microbiol. Rev. 2020, 33, e00047-19. [Google Scholar] [CrossRef]
- Wink, P.L.; Martins, A.S.; Volpato, F.; Zavascki, A.P.; Barth, A.L. Increased frequency of bla NDM in a tertiary care hospital in southern Brazil. Braz. J. Microbiol. 2021, 52, 299–301. [Google Scholar] [CrossRef]
- Zhang, B.; Zhu, Z.; Jia, W.; Qu, F.; Huang, B.; Shan, B.; Yu, H.; Tang, Y.; Chen, L.; Du, H. In vitro activity of aztreonam–avibactam against metallo-β-lactamase-producing Enterobacteriaceae—A multicenter study in China. Int. J. Infect. Dis. 2020, 97, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Veeraraghavan, B.; Bakthavatchalam, Y.D.; Soman, R.; Swaminathan, S.; Manesh, A.; Nagvekar, V.; Nangia, V. Management of serious infections caused by metallo β-lactamases with or without OXA-48-like expressing Enterobacterales with aztreonam and ceftazidime/avibactam combination: Dosing strategy for better clinical outcome. Indian. J. Med. Microbiol. 2021, 39, 286–288. [Google Scholar] [CrossRef]
- Perrotta, F.; Perrini, M.P. Successful Treatment of Klebsiella pneumoniae NDM Sepsis and Intestinal Decolonization with Ceftazidime/Avibactam Plus Aztreonam Combination in a Patient with TTP Complicated by SARS-CoV-2 Nosocomial Infection. Medicina 2021, 57, 424. [Google Scholar] [CrossRef]
- Alghoribi, M.F.; Alqurashi, M.; Okdah, L.; Alalwan, B.; AlHebaishi, Y.S.; Almalki, A.; Alzayer, M.A.; Alswaji, A.A.; Doumith, M.; Barry, M. Successful treatment of infective endocarditis due to pandrug-resistant Klebsiella pneumoniae with ceftazidime-avibactam and aztreonam. Sci. Rep. 2021, 11, 9684. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.; Carrara, E.; Retamar, P.; Tängdén, T.; Bitterman, R.; Bonomo, R.A.; de Waele, J.; Daikos, G.L.; Akova, M.; Harbarth, S.; et al. European Society of Clinical Microbiology and Infectious Diseases (ESCMID) guidelines for the treatment of infections caused by multidrug-resistant Gram-negative bacilli (endorsed by European society of intensive care medicine). Clin. Microbiol. Infect. 2022, 28, 521–547. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Infectious Diseases Society of America 2022 Guidance on the Treatment of Extended-Spectrum β-lactamase Producing Enterobacterales (ESBL-E), Carbapenem-Resistant Enterobacterales (CRE), and Pseudomonas aeruginosa with Difficult-to-Treat Resistance (DTR-P. aeruginosa). Clin. Infect. Dis. 2022, 75, 187–212. [Google Scholar] [PubMed]
- Marshall, S.; Hujer, A.M.; Rojas, L.J.; Papp-Wallace, K.M.; Humphries, R.M.; Spellberg, B.; Hujer, K.M.; Marshall, E.K.; Rudin, S.D.; Perez, F.; et al. Can ceftazidime-avibactam and aztreonam overcome B-lactam resistance conferred by metallo-B-lactamases in Enterobacteriaceae? Antimicrob. Agents Chemother. 2017, 61, e02243-16. [Google Scholar] [CrossRef]
- Monogue, M.L.; Abbo, L.M.; Rosa, R.; Camargo, J.F.; Martinez, O.; Bonomo, R.A.; Nicolau, D.P. In Vitro Discordance with In Vivo Activity: Humanized Exposures of Ceftazidime-Avibactam, Aztreonam, and Tigecycline Alone and in Combination against New Delhi Metallo-β-Lactamase- Producing Klebsiella pneumoniae in a Murine Lung Infection Model. Antimicrob. Agents Chemother. 2017, 61, e00486-17. [Google Scholar] [CrossRef]
- Wenzler, E.; Deraedt, M.F.; Harrington, A.T.; Danizger, L.H. Synergistic activity of ceftazidime-avibactam and aztreonam against serine and metallo-β-lactamase-producing gram-negative pathogens. Diagn. Microbiol. Infect. Dis. 2017, 88, 352–354. [Google Scholar] [CrossRef]
- Falcone, M.; Daikos, G.L.; Tiseo, G.; Bassoulis, D.; Giordano, C.; Galfo, V.; Leonildi, A.; Tagliaferri, E.; Barnini, S.; Sani, S.; et al. Efficacy of ceftazidime-avibactam plus aztreonam in patients with bloodstream infections caused by MBL-producing Enterobacterales. Clin. Infect. Dis. 2020, 586, 1871–1878. [Google Scholar]
- Sahu, C.; Pal, S.; Patel, S.S.; Singh, S.; Gurjar, M.; Ghoshal, U. Phenotypic synergy testing of ceftazidime–avibactam with aztreonam in a university hospital having high number of metallobetalactamase producing bacteria. Infect. Dis. 2020, 52, 801–807. [Google Scholar] [CrossRef]
- Khan, A.; Erickson, S.G.; Pettaway, C.; Arias, C.A.; Miller, W.R.; Bhatti, M.M. Evaluation of Susceptibility Testing Methods for Aztreonam and Ceftazidime-Avibactam Combination Therapy on Extensively Drug-Resistant Gram-Negative Organisms. Antimicrob. Agents Chemother. 2021, 65, e00846-21. [Google Scholar] [CrossRef]
- Avery, L.M.; Nicolau, D.P. Assessing the in vitro activity of ceftazidime/avibactam and aztreonam among carbapenemase-producing Enterobacteriaceae: Defining the zone of hope. Int. J. Antimicrob. Agents 2018, 52, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Maraki, S.; Mavromanolaki, V.E.; Moraitis, P.; Stafylaki, D.; Kasimati, A.; Magkafouraki, E.; Scoulica, E. Ceftazidime-avibactam, meropenen-vaborbactam, and imipenem-relebactam in combination with aztreonam against multidrug-resistant, metallo-β-lactamase-producing Klebsiella pneumoniae. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1755–1759. [Google Scholar] [CrossRef]
- Biagi, M.; Wu, T.; Lee, M.; Patel, S.; Butler, D.; Wenzler, E. Exploring aztreonam in combination with ceftazidime-avibactam or meropenem-vaborbactam as potential treatments for metallo- and serine-B-lactamase-producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2019, 63, e01426-19. [Google Scholar] [CrossRef] [PubMed]
- Bocanegra-Ibarias, P.; Camacho-Ortiz, A.; Garza-González, E.; Flores-Treviño, S.; Kim, H.; Perez-Alba, E. Aztreonam plus ceftazidime-avibactam as treatment of NDM-1-producing Klebsiella pneumoniae bacteraemia in a neutropenic patient: Last resort therapy? J. Glob. Antimicrob. Resist. 2020, 23, 417–419. [Google Scholar] [CrossRef]
- Liu, Z.; Hang, X.; Yan, T.; Chu, W.; Gong, Z.; Liu, Y.; Dai, Y.; Yang, M.; Li, J.; Zhou, Q. A Simple Disk Stacking Plus Micro-Elution Method for Rapid Detection of the Synergistic Effect of Aztreonam and Ceftazidime/Avibactam Against Metallo-β-Lactamase Producing Enterobacterales. Infect. Drug. Resist. 2023, 16, 1537–1543. [Google Scholar] [CrossRef]
- Lange, C.; Schubert, S.; Jung, J.; Kostrzewa, M.; Sparbier, K. Quantitative matrix-assisted laser desorption ionization-time of flight mass spectrometry for rapid resistance detection. J. Clin. Microbiol. 2014, 52, 4155–4162. [Google Scholar] [CrossRef] [PubMed]
- Idelevich, E.A.; Becker, K. Matrix-assisted laser desorption ionization-time of flight mass spectrometry for antimicrobial susceptibility testing. J. Clin. Microbiol. 2021, 59, e0181419. [Google Scholar] [CrossRef] [PubMed]
- Sparbier, K.; Schubert, S.; Kostrzewa, M. MBT-ASTRA: A suitable tool for fast antibiotic susceptibility testing? Methods 2016, 104, 48–54. [Google Scholar] [CrossRef]
- Jung, J.S.; Hamacher, C.; Gross, B.; Sparbier, K.; Lange, C.; Kostrzewa, M.; Schubert, S. Evaluation of a semiquantitative matrix-assisted laser desorption ionization-time of flight mass spectrometry method for rapid antimicrobial susceptibility testing of positive blood cultures. J. Clin. Microbiol. 2016, 54, 2820–2824. [Google Scholar] [CrossRef] [PubMed]
- Axelsson, C.; Rehnstam-Holm, A.S.; Nilson, B. Rapid detection of antibiotic resistance in positive blood cultures by MALDI-TOF MS and an automated and optimized MBT-ASTRA protocol for Escherichia coli and Klebsiella pneumoniae. Infect. Dis. 2020, 52, 45–53. [Google Scholar] [CrossRef]
- Monteiro, J.; Widen, R.H.; Pignatari, A.C.C.; Kubasek, C.; Silbert, S. Rapid detection of carbapenemase genes by multiplex real-time PCR. J. Antimicrob. Chemother. 2012, 67, 906–909. [Google Scholar] [CrossRef]
- Sachse, S.; Bresan, S.; Erhard, M.; Edel, B.; Pfister, W.; Saupe, A.; Rödel, J. Comparison of multilocus sequence typing, RAPD, and MALDI-TOF mass spectrometry for typing of β-lactam-resistant Klebsiella pneumoniae strains. Diagn. Microbiol. Infect. Dis. 2014, 80, 267–271. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 30 June 2022).
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Routine Extended Internal Quality Control for MIC Determination Disk Diffusion as Recommended by EUCAST. Version 12.0. Available online: https://www.eucast.org/ast_of_bacteria/quality_control (accessed on 13 April 2022).
- Clinical Laboratory Standards Institute (CLSI). Methods for Determining Bactericidal Activity of Antimicrobial Agents: Approved Guideline; NCCLS Document M26-A; National Committee for Clinical Laboratory Standards: Albany, NY, USA, 1999. [Google Scholar]
- Wilhelm, C.M.; Nunes, L.D.S.; Martins, A.F.; Barth, A.L. In vitro antimicrobial activity of imipenem plus amikacin or polymyxin B against carbapenem-resistant Pseudomonas aeruginosa isolates. Diagn. Microbiol. Infect. Dis. 2018, 92, 152–154. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing. Antimicrobial Susceptibility Testing EUCAST Disk Diffusion Method Version 10.0. Available online: https://www.eucast.org/ast_of_bacteria/disk_diffusion_methodology (accessed on 12 December 2022).
- Wang, Y.; Wang, J.; Wang, R.; Cai, Y. Resistance to ceftazidime-avibactam and underlying mechanisms. J. Glob. Antimicrob. Resist. 2020, 22, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Taha, R.; Kader, O.; Shawky, S.; Rezk, S. Ceftazidime-avibactam plus aztreonam synergistic combination tested against carbapenem-resistant Enterobacterales characterized phenotypically and genotypically: A glimmer of hope. Ann. Clin. Microbiol. Antimicrob. 2023, 22, 21. [Google Scholar] [CrossRef] [PubMed]
- Bianco, G.; Boattini, M.; Comini, S.; Casale, R.; Iannaccone, M.; Cavallo, R.; Costa, C. Occurrence of multi-carbapenemases producers among carbapenemase-producing Enterobacterales and in vitro activity of combinations including cefiderocol, ceftazidime-avibactam, meropenem-vaborbactam, and aztreonam in the COVID-19 era. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Leber, A.L. Synergism testing: Broth microdilution checkerboard and broth macrodilution methods. In Clinical Microbiology Procedures Handbook, 4th ed.; ASM Press: Washington, DC, USA, 2016; pp. 1–23. [Google Scholar]
- Lu, G.; Tang, H.; Xia, Z.; Yang, W.; Xu, H.; Liu, Z.; Ni, S.; Wang, Z.; Shen, J. In vitro and in vivo antimicrobial activities of ceftazidime/avibactam alone or in combination with aztreonam against carbapenem-resistant Enterobacterales. Infect. Drug. Resist. 2022, 15, 7107–7116. [Google Scholar] [CrossRef] [PubMed]
- Pragasam, A.K.; Veeraraghavan, B.; Shankar, B.; Bakthavatchalam, Y.; Mathuram, A.; George, B.; Chacko, B.; Korula, P.; Anandan, S. Will ceftazidime/avibactam plus aztreonam be effective for NDM and OXA-48-Like producing organisms: Lessons learnt from in vitro study. Indian J. Med. Microbiol. 2019, 37, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Han, R.; Guo, Y.; Yang, Y.; Wu, S.; Ding, L.; Zhang, R.; Yin, D.; Hu, F. Multiple novel ceftazidime-avibactam-resistant variants of bla KPC-2-positive Klebsiella pneumoniae in two patients. Microbiol. Spectr. 2022, 10, e01714-21. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antibiotic | Disk Diffusion (mm) | MIC (mg/L) | ||
|---|---|---|---|---|
| S | R | S | R | |
| Aztreonam | ≥26 | <21 | ≤1 | >4 |
| Ceftazidime/avibactam | ≥13 | <13 | ≤8 | >8 |
| Isolate | Cp | Cluster | ATM | CZA | ||
|---|---|---|---|---|---|---|
| DD (mm) | Cat. | DD (mm) | Cat. | |||
| Kp01 | KPC, NDM | F | 8 | R | 7 | R |
| Kp02 | KPC, NDM | H | 10 | R | 6 | R |
| Kp03 | KPC, NDM | A | 6 | R | 8 | R |
| Kp04 | KPC, NDM | A | 6 | R | 8 | R |
| Kp05 | KPC, NDM | H | 6 | R | 6 | R |
| Kp06 | KPC, NDM | E | 6 | R | 9 | R |
| Kp07 | KPC, NDM | C | 6 | R | 6 | R |
| Kp08 | KPC, NDM | E | 6 | R | 8 | R |
| Kp09 | KPC, NDM | E | 6 | R | 9 | R |
| Kp10 | KPC, NDM | G | 6 | R | 8 | R |
| Kp11 | NDM | G | 6 | R | 9 | R |
| Kp12 | NDM | G | 12 | R | 7 | R |
| Kp13 | NDM | B | 8 | R | 8 | R |
| Kp14 | NDM | G | 9 | R | 10 | R |
| Kp15 | NDM | D | 6 | R | 6 | R |
| Kp16 | NDM | F | 6 | R | 6 | R |
| Kp17 | NDM | F | 6 | R | 6 | R |
| Kp18 | NDM | C | 6 | R | 6 | R |
| Kp19 | NDM | D | 6 | R | 8 | R |
| Kp20 | NDM | D | 30 | S | 8 | R |
| Kp21 | KPC | F | 6 | R | 20 | S |
| Kp22 | NT | F | 30 | S | 24 | S |
| Isolate | Beta-Lactamase Genes | ATM | CZA | ATM + CZA | ||
|---|---|---|---|---|---|---|
| MIC (mg/L) | TKC * | MIC (mg/L) | TKC * | TKC * | ||
| Kp04 | NDM-1, KPC-2, CTX-M-15, SHV-187, TEM-181 | >1024 | 10.42 | >256 | 10.34 | 5.65 |
| Kp08 | NDM-1, KPC-2, OXA-1, SHV-187, TEM-181 | >1024 | 10.86 | >256 | 10.83 | 3.34 |
| Kp10 | NDM-1, KPC-2, OXA-1, CTX-M-15, SHV-187, TEM-181 | >1024 | 10.28 | >256 | 10.42 | 0.00 |
| Kp16 | NDM-1, OXA-1, CTX-M-15, SHV-187, TEM-181 | >1024 | 10.70 | >256 | 10.46 | 6.98 |
| Isolate | ATM + CZA | ATM | CZA | ||
|---|---|---|---|---|---|
| RG | RG | Dif. * | RG | Dif. * | |
| Kp01 | 0.1082 | 1.1196 | 1.0114 | 1.4997 | 1.3915 |
| Kp02 | 0.0544 | 0.8271 | 0.7726 | 0.9160 | 0.8616 |
| Kp03 | 0.1190 | 0.7915 | 0.6725 | 1.1907 | 1.0717 |
| Kp04 | 0.0793 | 0.9360 | 0.8566 | 0.9123 | 0.8330 |
| Kp05 | 0.1938 | 0.5617 | 0.3679 | 0.6647 | 0.4709 |
| Kp06 | 0.0704 | 0.7019 | 0.6316 | 0.7411 | 0.6708 |
| Kp07 | 0.0966 | 0.8282 | 0.7315 | 0.9710 | 0.8744 |
| Kp08 | 0.1042 | 0.8389 | 0.7346 | 1.5111 | 1.4068 |
| Kp09 | 0.0486 | 0.6440 | 0.5955 | 0.5841 | 0.5356 |
| Kp10 | 0.0841 | 0.8880 | 0.8040 | 0.7833 | 0.6993 |
| Kp11 | 0.0874 | 0.8129 | 0.7255 | 0.6141 | 0.5267 |
| Kp12 | 0.1127 | 0.7160 | 0.6033 | 0.7209 | 0.6082 |
| Kp13 | 0.4411 | 1.3313 | 0.8902 | 0.9592 | 0.5181 |
| Kp14 | 0.0912 | 0.7752 | 0.6840 | 0.8864 | 0.7952 |
| Kp15 | 0.0821 | 0.7618 | 0.6797 | 0.7712 | 0.6891 |
| Kp16 | 0.0696 | 0.9395 | 0.8699 | 1.0962 | 1.0266 |
| Kp17 | 0.0796 | 0.9623 | 0.8827 | 0.8398 | 0.7602 |
| Kp18 | 0.0813 | 0.8581 | 0.7768 | 1.2394 | 1.1581 |
| Kp19 | 0.5338 | 0.8967 | 0.3630 | 1.1305 | 0.5967 |
| Kp20 | 0.0528 | 0.3416 | 0.2888 | 0.6501 | 0.5973 |
| Kp21 | 0.0585 | 0.8503 | 0.7918 | 0.0647 | 0.0062 |
| Kp22 | 0.0311 | 0.2195 | 0.1884 | 0.0314 | 0.0003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilhelm, C.M.; Inamine, E.; Martins, A.F.; Barth, A.L. Evaluation of Aztreonam and Ceftazidime/Avibactam Synergism against Klebsiella pneumoniae by MALDI-TOF MS. Antibiotics 2023, 12, 1063. https://doi.org/10.3390/antibiotics12061063
Wilhelm CM, Inamine E, Martins AF, Barth AL. Evaluation of Aztreonam and Ceftazidime/Avibactam Synergism against Klebsiella pneumoniae by MALDI-TOF MS. Antibiotics. 2023; 12(6):1063. https://doi.org/10.3390/antibiotics12061063
Chicago/Turabian StyleWilhelm, Camila Mörschbächer, Everton Inamine, Andreza Francisco Martins, and Afonso Luís Barth. 2023. "Evaluation of Aztreonam and Ceftazidime/Avibactam Synergism against Klebsiella pneumoniae by MALDI-TOF MS" Antibiotics 12, no. 6: 1063. https://doi.org/10.3390/antibiotics12061063
APA StyleWilhelm, C. M., Inamine, E., Martins, A. F., & Barth, A. L. (2023). Evaluation of Aztreonam and Ceftazidime/Avibactam Synergism against Klebsiella pneumoniae by MALDI-TOF MS. Antibiotics, 12(6), 1063. https://doi.org/10.3390/antibiotics12061063