
Genomics Insight into cfr-Mediated Linezolid-Resistant LA-MRSA in Italian Pig Holdings
 , , , , and
, , , , and
Abstract
1. Introduction
2. Results
2.1. Antimicrobial Susceptibility Testing (AST)
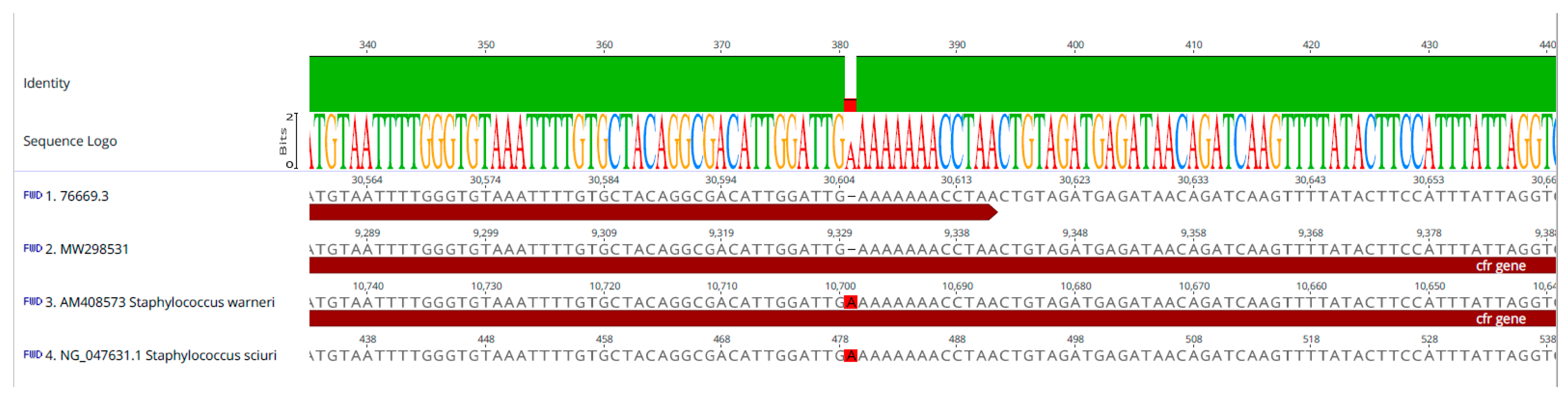
2.2. WGS and Bioinformatics Analysis (Illumina Short Reads)
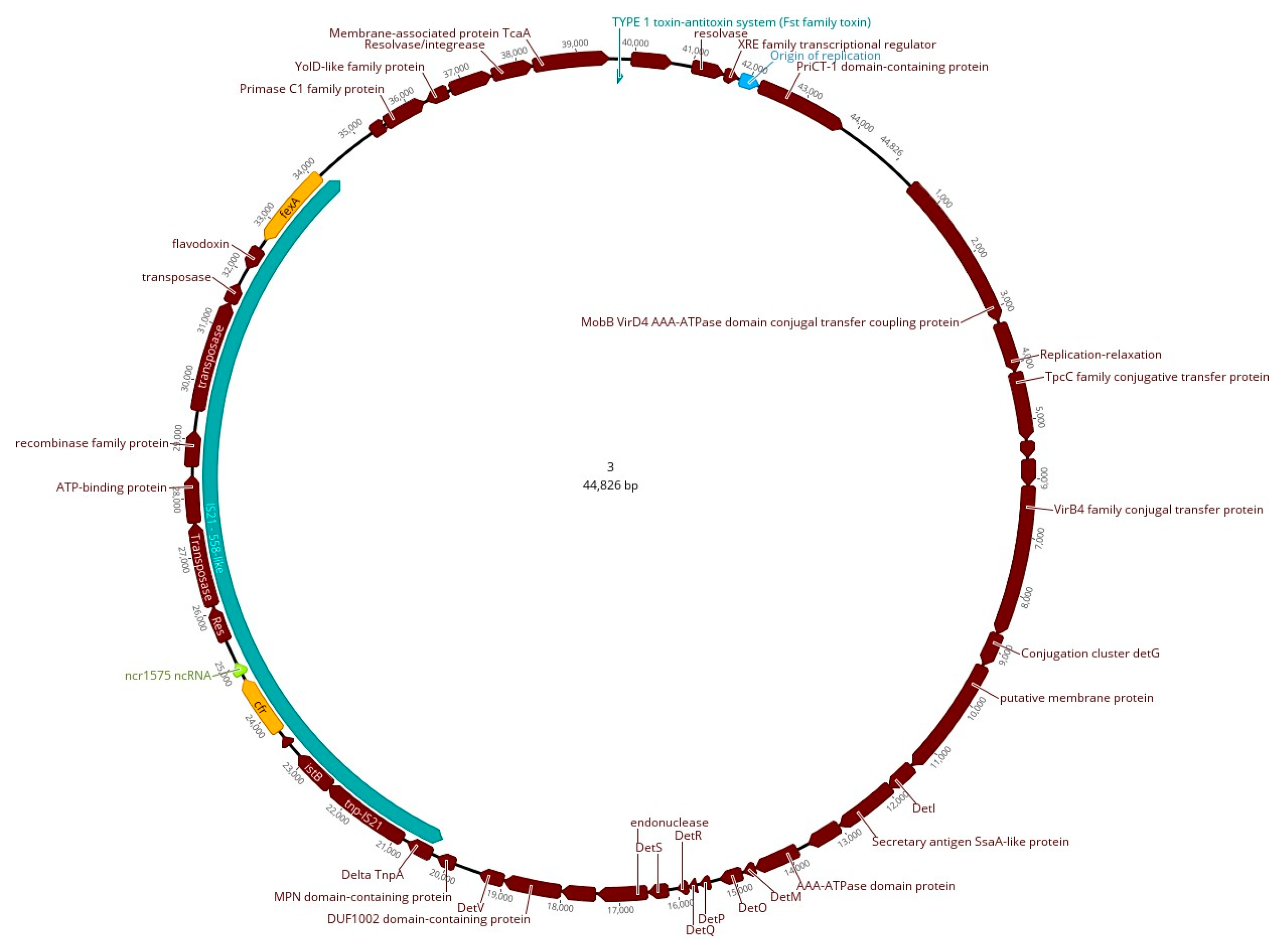
2.3. WGS and Bioinformatics Analysis (Hybrid Assembly)
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. LA-MRSA Isolates
5.2. Antimicrobial Susceptibility Testing (AST)
5.3. Library Preparation and Whole Genome Sequencing (WGS)
5.4. Bioinformatics Analysis (Illumina Short Reads)
5.5. Bioinformatics Analysis (Hybrid Assembly)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gu, B.; Kelesidis, T.; Tsiodras, S.; Hindler, J.; Humphries, R.M. The emerging problem of linezolid-resistant Staphylococcus. J. Antimicrob. Chemother. 2013, 68, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Zhang, W.; Du, X.D.; Krüger, H.; Feßler, A.T.; Ma, S.; Zhu, Y.; Wu, C.; Shen, J.; Wang, Y. Mobile Oxazolidinone Resistance Genes in Gram-Positive and Gram-Negative Bacteria. Clin. Microbiol. Rev. 2021, 34, e0018820. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Werckenthin, C.; Kehrenberg, C. Identification of a plasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococcus sciuri. Antimicrob. Agents Chemother. 2000, 44, 2530–2533. [Google Scholar] [CrossRef]
- Long, K.S.; Poehlsgaard, J.; Kehrenberg, C.; Schwarz, S.; Vester, B. The cfr rRNA methyltransferase confers resistance to phenicols, lincosamides, oxazolidinones, pleuromutilins, and streptogramin A antibiotics. Antimicrob. Agents Chemother. 2006, 50, 2500–2505. [Google Scholar] [CrossRef]
- Antonelli, A.; D’Andrea, M.M.; Galano, A.; Borchi, B.; Brenciani, A.; Vaggelli, G.; Cavallo, A.; Bartoloni, A.; Giovanetti, E.; Rossolini, G.M. Linezolid-resistant cfr-positive MRSA, Italy. J. Antimicrob. Chemother. 2016, 71, 2349–2351. [Google Scholar] [CrossRef]
- Youenou, B.; Martins Simoes, P.; Tristan, A.; Farfour, E.; Beauruelle, C.; Kolenda, C.; Ranc, A.G.; Vandenesch, F.; Laurent, F.; Dupieux, C. Linezolid resistance: Detection of the cfr(B) gene in French clinical MRSA strains. J. Antimicrob. Chemother. 2022, 13, dkac411. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Schwarz, S. Distribution of Florfenicol Resistance Genes fexA and cfr among Chloramphenicol-Resistant Staphylococcus Isolates. Antimicrob. Agents Chemother. 2006, 50, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Peeters, L.E.J.; Argudín, M.A.; Azadikhah, S.; Butaye, P. Antimicrobial resistance and population structure of Staphylococcus aureus recovered from pigs farms. Vet. Microbiol. 2015, 180, 151–156. [Google Scholar] [CrossRef]
- Ruiz-Ripa, L.; Bellés-Bellés, A.; Fernández-Fernández, R.; García, M.; Vilaró, A.; Zarazaga, M.; Torres, C. Linezolid-resistant MRSA-CC398 carrying the cfr gene, and MRSA-CC9 isolates from pigs with signs of infection in Spain. J. Appl. Microbiol. 2021, 131, 615–622. [Google Scholar] [CrossRef]
- Leão, C.; Clemente, L.; Cara d’Anjo, M.; Albuquerque, T.; Amaro, A. Emergence of Cfr-Mediated Linezolid Resistance among Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) from Healthy Pigs in Portugal. Antibiotics 2022, 11, 1439. [Google Scholar] [CrossRef]
- Schouls, L.M.; Veldman, K.; Brouwer, M.S.M.; Dierikx, C.; Witteveen, S.; van Santen-Verheuvel, M.; Hendrickx, A.P.A.; Landman, F.; Hengeveld, P.; Wullings, B.; et al. Dutch MRSA surveillance study group. cfr and fexA genes in methicillin-resistant Staphylococcus aureus from humans and livestock in the Netherlands. Commun. Med. 2022, 2, 135. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-M.; Zhou, Y.-F.; Li, L.; Fang, L.-X.; Duan, J.-H.; Liu, F.-R.; Liang, H.-Q.; Wu, Y.-T.; Gu, W.-Q.; Liao, X.-P.; et al. Characterization of the multi-drug resistance gene cfr in methicillin-resistant Staphylococcus aureus (MRSA) strains isolated from animals and humans in China. Front. Microbiol. 2018, 9, 2925. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.Y.; Moon, D.C.; Mechesso, A.F.; Choi, J.-H.; Kim, S.-J.; Song, H.-J.; Kim, M.H.; Yoon, S.-S.; Lim, S.-K. Emergence of cfr-mediated linezolid resistance in Staphylococcus aureus isolated from pig carcasses. Antibiotics 2020, 9, 769. [Google Scholar] [CrossRef] [PubMed]
- Osman, K.M.; Amer, A.M.; Badr, J.M.; Helmy, N.M.; Elhelw, R.A.; Orabi, A.; Bakry, M.; Saad, A.S.A. Antimicrobial resistance, biofilm formation and mecA characterization of methicillin-susceptible S. aureus and non-S. aureus of beef meat origin in Egypt. Front. Microbiol. 2016, 7, 222. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wang, Y.; Schwarz, S. Presence and dissemination of the multiresistance gene cfr in Gram-positive and Gram-negative bacteria. J. Antimicrob. Chemother. 2013, 68, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Alba, P.; Feltrin, F.; Cordaro, G.; Porrero, M.C.; Kraushaar, B.; Argudín, M.A.; Nykäsenoja, S.; Monaco, M.; Stegger, M.; Aarestrup, F.M.; et al. Livestock-Associated Methicillin Resistant and Methicillin Susceptible Staphylococcus aureus Sequence Type (CC)1 in European Farmed Animals: High Genetic Relatedness of Isolates from Italian Cattle Herds and Humans. PLoS ONE 2015, 10, e0137143. [Google Scholar] [CrossRef]
- Feltrin, F.; Alba, P.; Amoruso, R.; Argudin, M.; Lauzat, B.; Iurescia, M.; Sorbara, L.; Dottarelli, S.; Carfora, V.; Menichini, E.; et al. Molecular characterization of the Staphylococcus aureus CC398 population in Italy. In Proceedings of the 11th International Meeting on Microbial Epidemiological Markers (ESCMID-ESGEM Conference), Estoril, Portugal, 9–12 March 2016. [Google Scholar]
- Fioriti, S.; Coccitto, S.N.; Morroni, G.; Simoni, S.; Cinthi, M.; Magi, G.; Sante, L.D.; Alvarez-Suarez, J.M.; Mingoia, M.; Giovanetti, E.; et al. Detection of a chromosomal truncated cfr gene in a linezolid-susceptible LA-MRSA ST398 isolate of porcine origin, Italy. J. Glob. Antimicrob. Resist. 2021, 26, 199–201. [Google Scholar] [CrossRef]
- Mendes, R.E.; Deshpande, L.M.; Bonilla, H.F.; Schwarz, S.; Huband, M.D.; Jones, R.N.; Quinn, J.P. Dissemination of a pSCFS3-like cfr-carrying plasmid in Staphylococcus aureus and Staphylococcus epidermidis clinical isolates recovered from hospitals in Ohio. Antimicrob. Agents Chemother. 2013, 57, 2923–2928. [Google Scholar] [CrossRef]
- Battisti, A.; (National Reference Laboratory for Antimicrobial Resistance, Department of General Diagnostics, Istituto Zooprofilattico Sperimentale del Lazio e della Toscana “M. Aleandri”, Rome, Italy). Personal Communication, 2021.
- Gales, A.C.; Deshpande, L.M.; de Souza, A.G.; Pignatari, A.C.; Mendes, R.E. MSSA ST398/t034 carrying a plasmid-mediated Cfr and Erm(B) in Brazil. J. Antimicrob. Chemother. 2015, 70, 303–305. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Aarestrup, F.M.; Schwarz, S. IS21-558 insertion sequences are involved in the mobility of the multiresistance gene cfr. Antimicrob. Agents Chemother. 2007, 51, 483–487. [Google Scholar] [CrossRef]
- EFSA. Analysis of the baseline survey on the prevalence of methicillin-resistant Staphylococcus aureus (MRSA) in holdings with breeding pigs, in the EU, 2008. Part A: MRSA prevalence estimates; on request from the European Commission. EFSA J. 2009, 7, 1376. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority); Aerts, M.; Battisti, A.; Hendriksen, R.; Larsen, J.; Nilsson, O.; Cortinas Abrahantes, J.; Guerra, B.; Papanikolaou, A.; Beloeil, P.-A. Technical specifications for a baseline survey on the prevalence of Methicillin Resistant Staphylococcus aureus (MRSA) in pigs. EFSA J. 2022, 20, e07620. [Google Scholar] [CrossRef] [PubMed]
- Battisti, A.; Franco, A.; Merialdi, G.; Hasman, H.; Iurescia, M.; Lorenzetti, R.; Feltrin, F.; Zini, M.; Aarestrup, F.M. Heterogeneity among methicillin-resistant Staphylococcus aureus from Italian pig finishing holdings. Vet. Microbiol. 2010, 142, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Cortimiglia, C.; Luini, M.; Bianchini, V.; Marzagalli, L.; Vezzoli, F.; Avisani, D.; Bertoletti, M.; Ianzano, A.; Franco, A.; Battisti, A. Prevalence of Staphylococcus aureus and of methicillin-resistant S. aureus clonal complexes in bulk tank milk from dairy cattle herds in Lombardy Region (Northern Italy). Epidemiol. Infect. 2016, 144, 3046–3051. [Google Scholar] [CrossRef]
- Feltrin, F.; Alba, P.; Kraushaar, B.; Ianzano, A.; Argudín, M.A.; Di Matteo, P.; Porrero, M.C.; Aarestrup, F.M.; Butaye, P.; Franco, A.; et al. A Livestock-Associated, Multidrug-Resistant, Methicillin-Resistant Staphylococcus aureus Clonal Complex 97 Lineage Spreading in Dairy Cattle and Pigs in Italy. Appl. Environ. Microbiol. 2015, 82, 816–821. [Google Scholar] [CrossRef]
- Tomao, P.; Pirolo, M.; Agnoletti, F.; Pantosti, A.; Battisti, A.; Di Martino, G.; Visaggio, D.; Monaco, M.; Franco, A.; Pimentel de Araujo, F.; et al. Molecular epidemiology of methicillin-resistant Staphylococcus aureus from dairy farms in North-eastern Italy. Int. J. Food Microbiol. 2020, 332, 108817. [Google Scholar] [CrossRef]
- Zoppi, S.; Dondo, A.; Di Blasio, A.; Vitale, N.; Carfora, V.; Goria, M.; Chiavacci, L.; Giorgi, I.; D’Errico, V.; Irico, L.; et al. Livestock-Associated Methicillin-Resistant Staphylococcus aureus and Related Risk Factors in Holdings of Veal Calves in Northwest Italy. Microb. Drug Resist. 2021, 27, 1136–1143. [Google Scholar] [CrossRef]
- Alba, P.; Taddei, R.; Cordaro, G.; Fontana, M.C.; Toschi, E.; Gaibani, P.; Marani, I.; Giacomi, A.; Diaconu, E.L.; Iurescia, M.; et al. Carbapenemase IncF-borne blaNDM-5 gene in the E. coli ST167 high-risk clone from canine clinical infection, Italy. Vet. Microbiol. 2021, 256, 109045. [Google Scholar] [CrossRef]
- Branton, D.; Deamer, D.W.; Marziali, A.; Bayley, H.; Benner, S.A.; Butler, T.; Di Ventra, M.; Garaj, S.; Hibbs, A.; Huang, X.; et al. The potential and challenges of nanopore sequencing. Nat. Biotechnol. 2008, 26, 1146–1153. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef]
- International Working Group on The Classification of Staphylococcal Cassette Chromosome Elements (IWG-SCC). Classification of staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents Chemother. 2009, 56, 4961–4967. [Google Scholar]
- Kondo, Y.; Ito, T.; Ma, X.X.; Watanabe, S.; Kreiswirth, B.N.; Etienne, J.; Hiramatsu, K. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: Rapid identification system for mec, ccr and major difference in junkyard regions. Antimicrob. Agents Chemother. 2007, 51, 264–274. [Google Scholar] [CrossRef]
- Bartels, M.D.; Petersen, A.; Worning, P.; Nielsen, J.B.; Larner-Svensson, H.; Johansen, H.K.; Andersen, L.P.; Jarløv, J.O.; Boye, K.; Larsen, A.R.; et al. Comparing whole-genome sequencing with Sanger sequencing for spa typing of methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2014, 52, 4305–4308. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.F.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.R.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2020, 72, 2764–2768. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Diaconu, E.L.; Carfora, V.; Alba, P.; Di Matteo, P.; Stravino, F.; Buccella, C.; Dell’Aira, E.; Onorati, R.; Sorbara, L.; Battisti, A.; et al. Novel IncFII plasmid harbouring blaNDM-4 in a carbapenem-resistant Escherichia coli of pig origin, Italy. J. Antimicrob. Chemother. 2020, 75, 3475–3479. [Google Scholar] [CrossRef] [PubMed]
- Schwengers, O.; Jelonek, L.; Dieckmann, M.A.; Beyvers, S.; Blom, J.; Goesmann, A. Bakta: Rapid and standardized annotation of bacterial genomes via alignment-free sequence identification. Microb. Genom. 2021, 7, 000685. [Google Scholar] [CrossRef]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA Hybridization Values and Their Relationship to Whole-Genome Sequence Similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Ojo, K.K.; Schwarz, S. Nucleotide sequence and organization of the multiresistance plasmid pSCFS1 from Staphylococcus sciuri. J. Antimicrob. Chemother. 2004, 54, 936–939. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MRSA ID | Year of Isolation | Matrix of Origin | Host Production | Spa Type | ST | CC | SCCmec Type | AMR Phenotype | AMR Genotype |
|---|---|---|---|---|---|---|---|---|---|
| 21554/1 § | 2008 | Dust sample | Pig breeding | t127 | 1 | 1 | V | CHL, CIP, TET, FOX, CLI, ERY, LIZ, PEN, TML, SYN | ant(9)-Ia °, aadD, bleO, mecA, blaZ, erm(A) °, cfr °°, fexA °°, tet(K), grlA (p.S80F), gyrA (p.S84L) |
| 76669/3 # | 2010 | Nasal swab | Pig finishing | t034 | 398 | 398 | Vc | CHL, CIP, TET, TRI, FOX, PEN, TML | mecA, blaZ, vga(A)V, cfr *°°, fexA °°, tet(K), tet(M), dfrG, grlA (p.S80F), gyrA (p.S84L) |
| 55864/23 § | 2011 | Dust sample | Pig breeding and finishing | t127 | 1 | 1 | V | CHL, CIP, GEN, KAN, TET, FOX, CLI, ERY, PEN | aac(6′)-aph(2″), mecA, blaZ, erm(A) °, erm(C), cfr *°°, fexA °°, tet(K), grlA (p.S80F), gyrA (p.S84L) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iurescia, M.; Diaconu, E.L.; Alba, P.; Feltrin, F.; Buccella, C.; Onorati, R.; Giacomi, A.; Caprioli, A.; Franco, A.; Battisti, A.; et al. Genomics Insight into cfr-Mediated Linezolid-Resistant LA-MRSA in Italian Pig Holdings. Antibiotics 2023, 12, 530. https://doi.org/10.3390/antibiotics12030530
Iurescia M, Diaconu EL, Alba P, Feltrin F, Buccella C, Onorati R, Giacomi A, Caprioli A, Franco A, Battisti A, et al. Genomics Insight into cfr-Mediated Linezolid-Resistant LA-MRSA in Italian Pig Holdings. Antibiotics. 2023; 12(3):530. https://doi.org/10.3390/antibiotics12030530
Chicago/Turabian StyleIurescia, Manuela, Elena Lavinia Diaconu, Patricia Alba, Fabiola Feltrin, Carmela Buccella, Roberta Onorati, Angelo Giacomi, Andrea Caprioli, Alessia Franco, Antonio Battisti, and et al. 2023. "Genomics Insight into cfr-Mediated Linezolid-Resistant LA-MRSA in Italian Pig Holdings" Antibiotics 12, no. 3: 530. https://doi.org/10.3390/antibiotics12030530
APA StyleIurescia, M., Diaconu, E. L., Alba, P., Feltrin, F., Buccella, C., Onorati, R., Giacomi, A., Caprioli, A., Franco, A., Battisti, A., & Carfora, V. (2023). Genomics Insight into cfr-Mediated Linezolid-Resistant LA-MRSA in Italian Pig Holdings. Antibiotics, 12(3), 530. https://doi.org/10.3390/antibiotics12030530