
Comparative Genomics Identifies Novel Genetic Changes Associated with Oxacillin, Vancomycin and Daptomycin Susceptibility in ST100 Methicillin-Resistant Staphylococcus aureus

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Antimicrobial Susceptibility
2.2. General Genomic Features
2.3. Mutations Associated with hVISA/VISA
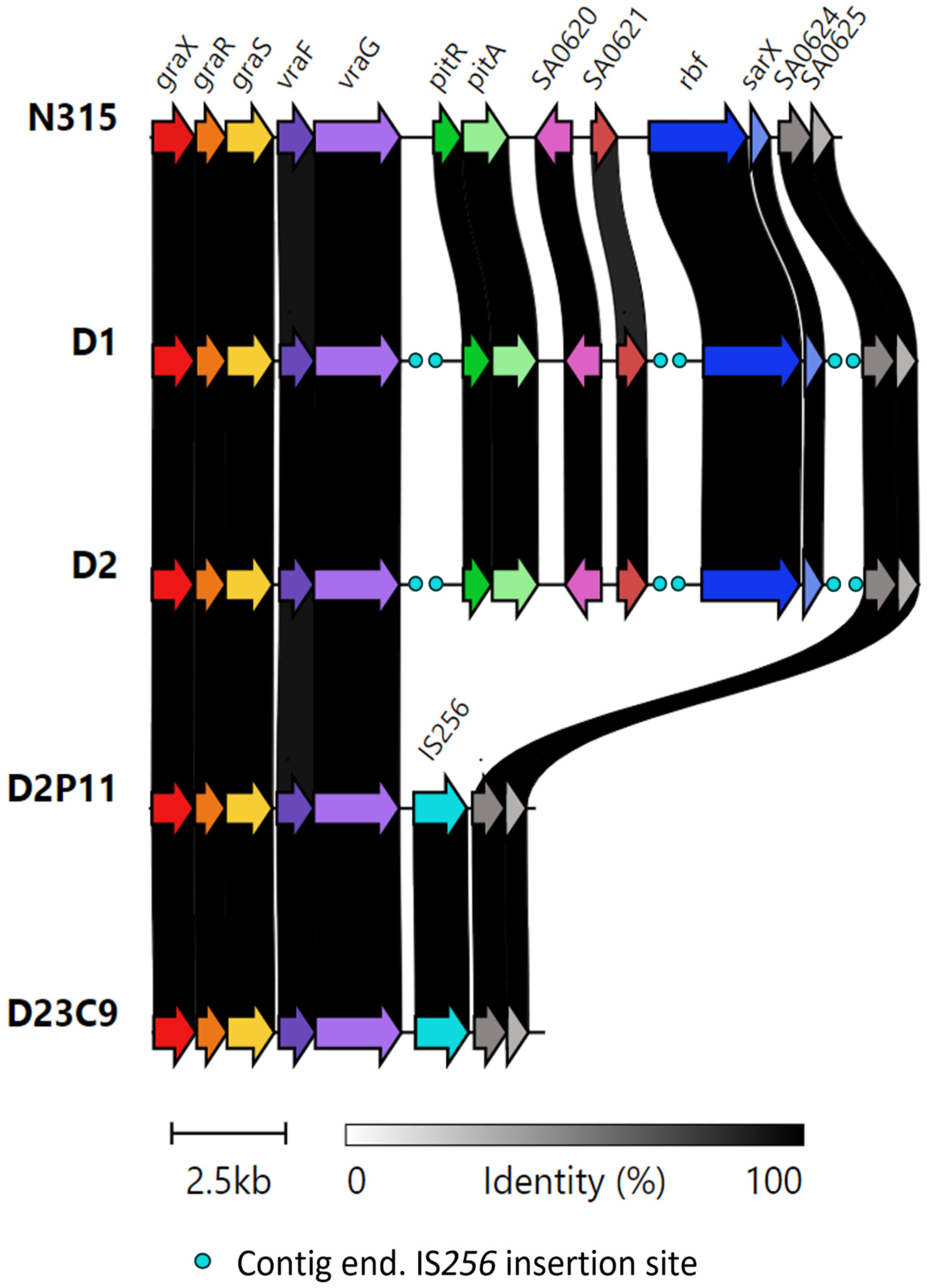
2.4. IS256-Mediated Genomic Rearrangements
2.5. Biofilm Formation
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Antimicrobial Susceptibility Testing
4.3. DNA Extraction and Whole Genome Sequencing
4.4. Whole Genome Sequencing Analysis
4.5. Growth Curves
4.6. Transmission Electron Microscopy (TEM)
4.7. Biofilm Production Assay
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 20 December 2022).
- Howden, B.P.; Peleg, A.Y.; Stinear, T.P. The Evolution of Vancomycin Intermediate Staphylococcus Aureus (VISA) and Heterogenous-VISA. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2014, 21, 575–582. [Google Scholar] [CrossRef]
- Shariati, A.; Dadashi, M.; Moghadam, M.T.; van Belkum, A.; Yaslianifard, S.; Darban-Sarokhalil, D. Global Prevalence and Distribution of Vancomycin Resistant, Vancomycin Intermediate and Heterogeneously Vancomycin Intermediate Staphylococcus Aureus Clinical Isolates: A Systematic Review and Meta-Analysis. Sci. Rep. 2020, 10, 12689. [Google Scholar] [CrossRef]
- Gostev, V.V.; Popenko, L.N.; Chernen’kaia, T.V.; Naumenko, Z.S.; Voroshilova, T.M.; Zakharova, I.A.; Khokhlova, O.E.; Kruglov, A.N.; Ershova, M.G.; Angelova, S.N.; et al. Estimation of MRSA susceptibility to oxacillin, cefoxitine, vancomycin and daptomycin. Antibiot. Chemoterapy 2013, 58, 13–20. [Google Scholar]
- Di Gregorio, S.; Perazzi, B.; Ordoñez, A.M.; De Gregorio, S.; Foccoli, M.; Lasala, M.B.; García, S.; Vay, C.; Famiglietti, A.; Mollerach, M. Clinical, Microbiological, and Genetic Characteristics of Heteroresistant Vancomycin-Intermediate Staphylococcus Aureus Bacteremia in a Teaching Hospital. Microb. Drug Resist. 2015, 21, 25–34. [Google Scholar] [CrossRef]
- Hiramatsu, K.; Hanaki, H.; Ino, T.; Yabuta, K.; Oguri, T.; Tenover, F.C. Methicillin-Resistant Staphylococcus Aureus Clinical Strain with Reduced Vancomycin Susceptibility. J. Antimicrob. Chemother. 1997, 40, 135–136. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, K.; Aritaka, N.; Hanaki, H.; Kawasaki, S.; Hosoda, Y.; Hori, S.; Fukuchi, Y.; Kobayashi, I. Dissemination in Japanese Hospitals of Strains of Staphylococcus Aureus Heterogeneously Resistant to Vancomycin. Lancet 1997, 350, 1670–1673. [Google Scholar] [CrossRef]
- Adhikari, R.P.; Scales, G.C.; Kobayashi, K.; Smith, J.M.B.; Berger-Bächi, B.; Cook, G.M. Vancomycin-Induced Deletion of the Methicillin Resistance Gene MecA in Staphylococcus Aureus. J. Antimicrob. Chemother. 2004, 54, 360–363. [Google Scholar] [CrossRef]
- Reipert, A.; Ehlert, K.; Kast, T.; Bierbaum, G. Morphological and Genetic Differences in Two Isogenic Staphylococcus Aureus Strains with Decreased Susceptibilities to Vancomycin. Antimicrob. Agents Chemother. 2003, 47, 568–576. [Google Scholar] [CrossRef]
- Cui, L.; Tominaga, E.; Neoh, H.-M.; Hiramatsu, K. Correlation between Reduced Daptomycin Susceptibility and Vancomycin Resistance in Vancomycin-Intermediate Staphylococcus Aureus. Antimicrob. Agents Chemother. 2006, 50, 1079–1082. [Google Scholar] [CrossRef]
- Hiramatsu, K.; Kayayama, Y.; Matsuo, M.; Aiba, Y.; Saito, M.; Hishinuma, T.; Iwamoto, A. Vancomycin-Intermediate Resistance in Staphylococcus Aureus. J. Glob. Antimicrob. Resist. 2014, 2, 213–224. [Google Scholar] [CrossRef]
- Hu, Q.; Peng, H.; Rao, X. Molecular Events for Promotion of Vancomycin Resistance in Vancomycin Intermediate Staphylococcus Aureus. Front. Microbiol. 2016, 7, 1601. [Google Scholar] [CrossRef]
- Alam, M.T.; Petit, R.A.; Crispell, E.K.; Thornton, T.A.; Conneely, K.N.; Jiang, Y.; Satola, S.W.; Read, T.D. Dissecting Vancomycin-Intermediate Resistance in Staphylococcus Aureus Using Genome-Wide Association. Genome Biol. Evol. 2014, 6, 1174–1185. [Google Scholar] [CrossRef]
- Maki, H.; McCallum, N.; Bischoff, M.; Wada, A.; Berger-Bächi, B. TcaA Inactivation Increases Glycopeptide Resistance in Staphylococcus Aureus. Antimicrob. Agents Chemother. 2004, 48, 1953–1959. [Google Scholar] [CrossRef]
- Jansen, A.; Türck, M.; Szekat, C.; Nagel, M.; Clever, I.; Bierbaum, G. Role of Insertion Elements and YycFG in the Development of Decreased Susceptibility to Vancomycin in Staphylococcus Aureus. Int. J. Med. Microbiol. IJMM 2007, 297, 205–215. [Google Scholar] [CrossRef]
- McEvoy, C.R.E.; Tsuji, B.; Gao, W.; Seemann, T.; Porter, J.L.; Doig, K.; Ngo, D.; Howden, B.P.; Stinear, T.P. Decreased Vancomycin Susceptibility in Staphylococcus Aureus Caused by IS256 Tempering of WalKR Expression. Antimicrob. Agents Chemother. 2013, 57, 3240–3249. [Google Scholar] [CrossRef]
- Kuroda, M.; Sekizuka, T.; Matsui, H.; Ohsuga, J.; Ohshima, T.; Hanaki, H. IS256-Mediated Overexpression of the WalKR Two-Component System Regulon Contributes to Reduced Vancomycin Susceptibility in a Staphylococcus Aureus Clinical Isolate. Front. Microbiol. 2019, 10, 1882. [Google Scholar] [CrossRef]
- Yin, Y.; Chen, H.; Li, S.; Gao, H.; Sun, S.; Li, H.; Wang, R.; Jin, L.; Liu, Y.; Wang, H. Daptomycin Resistance in Methicillin-Resistant Staphylococcus Aureus Is Conferred by IS256 Insertion in the Promoter of MprF along with Mutations in MprF and WalK. Int. J. Antimicrob. Agents 2019, 54, 673–680. [Google Scholar] [CrossRef]
- Casagrande Proietti, P.; Bietta, A.; Coletti, M.; Marenzoni, M.L.; Scorza, A.V.; Passamonti, F. Insertion Sequence IS256 in Canine Pyoderma Isolates of Staphylococcus Pseudintermedius Associated with Antibiotic Resistance. Vet. Microbiol. 2012, 157, 376–382. [Google Scholar] [CrossRef]
- He, T.; Wang, Y.; Schwarz, S.; Zhao, Q.; Shen, J.; Wu, C. Genetic Environment of the Multi-Resistance Gene Cfr in Methicillin-Resistant Coagulase-Negative Staphylococci from Chickens, Ducks, and Pigs in China. Int. J. Med. Microbiol. IJMM 2014, 304, 257–261. [Google Scholar] [CrossRef]
- Byrne, M.E.; Rouch, D.A.; Skurray, R.A. Nucleotide Sequence Analysis of IS256 from the Staphylococcus Aureus Gentamicin-Tobramycin-Kanamycin-Resistance Transposon Tn4001. Gene 1989, 81, 361–367. [Google Scholar] [CrossRef]
- Kleinert, F.; Kallies, R.; Hort, M.; Zweynert, A.; Szekat, C.; Nagel, M.; Bierbaum, G. Influence of IS256 on Genome Variability and Formation of Small-Colony Variants in Staphylococcus Aureus. Antimicrob. Agents Chemother. 2017, 61, e00144-17. [Google Scholar] [CrossRef]
- Benson, M.A.; Ohneck, E.A.; Ryan, C.; Alonzo, F.; Smith, H.; Narechania, A.; Kolokotronis, S.-O.; Satola, S.W.; Uhlemann, A.-C.; Sebra, R.; et al. Evolution of Hypervirulence by a MRSA Clone through Acquisition of a Transposable Element. Mol. Microbiol. 2014, 93, 664–681. [Google Scholar] [CrossRef]
- Di Gregorio, S.; Fernandez, S.; Perazzi, B.; Bello, N.; Famiglietti, A.; Mollerach, M. Increase in IS256 Transposition in Invasive Vancomycin Heteroresistant Staphylococcus Aureus Isolate Belonging to ST100 and Its Derived VISA Mutants. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 43, 197–202. [Google Scholar] [CrossRef]
- Loessner, I.; Dietrich, K.; Dittrich, D.; Hacker, J.; Ziebuhr, W. Transposase-Dependent Formation of Circular IS256 Derivatives in Staphylococcus Epidermidis and Staphylococcus Aureus. J. Bacteriol. 2002, 184, 4709–4714. [Google Scholar] [CrossRef]
- Valle, J.; Vergara-Irigaray, M.; Merino, N.; Penadés, J.R.; Lasa, I. SigmaB Regulates IS256-Mediated Staphylococcus Aureus Biofilm Phenotypic Variation. J. Bacteriol. 2007, 189, 2886–2896. [Google Scholar] [CrossRef]
- Cafiso, V.; Bertuccio, T.; Santagati, M.; Demelio, V.; Spina, D.; Nicoletti, G.; Stefani, S. Agr-Genotyping and Transcriptional Analysis of Biofilm-Producing Staphylococcus Aureus. FEMS Immunol. Med. Microbiol. 2007, 51, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.E.; Gillespie, M.T.; Skurray, R.A. Molecular Analysis of a Gentamicin Resistance Transposonlike Element on Plasmids Isolated from North American Staphylococcus Aureus Strains. Antimicrob. Agents Chemother. 1990, 34, 2106–2113. [Google Scholar] [CrossRef]
- Alam, M.M.; Kobayashi, N.; Uehara, N.; Watanabe, N. Analysis on Distribution and Genomic Diversity of High-Level Antiseptic Resistance Genes QacA and QacB in Human Clinical Isolates of Staphylococcus Aureus. Microb. Drug Resist. 2003, 9, 109–121. [Google Scholar] [CrossRef]
- Di Gregorio, S.; Fernandez, S.; Cuirolo, A.; Verlaine, O.; Amoroso, A.; Mengin-Lecreulx, D.; Famiglietti, A.; Joris, B.; Mollerach, M. Different Vancomycin-Intermediate Staphylococcus Aureus Phenotypes Selected from the Same ST100-HVISA Parental Strain. Microb. Drug Resist. 2017, 23, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Passalacqua, K.D.; Satola, S.W.; Crispell, E.K.; Read, T.D. A Mutation in the PP2C Phosphatase Gene in a Staphylococcus Aureus USA300 Clinical Isolate with Reduced Susceptibility to Vancomycin and Daptomycin. Antimicrob. Agents Chemother. 2012, 56, 5212–5223. [Google Scholar] [CrossRef]
- Kolar, S.L.; Nagarajan, V.; Oszmiana, A.; Rivera, F.E.; Miller, H.K.; Davenport, J.E.; Riordan, J.T.; Potempa, J.; Barber, D.S.; Koziel, J.; et al. NsaRS Is a Cell-Envelope-Stress-Sensing Two-Component System of Staphylococcus Aureus. Microbiology 2011, 157, 2206–2219. [Google Scholar] [CrossRef]
- Wheeler, R.; Turner, R.D.; Bailey, R.G.; Salamaga, B.; Mesnage, S.; Mohamad, S.A.S.; Hayhurst, E.J.; Horsburgh, M.; Hobbs, J.K.; Foster, S.J. Bacterial Cell Enlargement Requires Control of Cell Wall Stiffness Mediated by Peptidoglycan Hydrolases. mBio 2015, 6, e00660. [Google Scholar] [CrossRef]
- Hiron, A.; Falord, M.; Valle, J.; Débarbouillé, M.; Msadek, T. Bacitracin and Nisin Resistance in Staphylococcus Aureus: A Novel Pathway Involving the BraS/BraR Two-Component System (SA2417/SA2418) and Both the BraD/BraE and VraD/VraE ABC Transporters. Mol. Microbiol. 2011, 81, 602–622. [Google Scholar] [CrossRef]
- Wood, A.; Irving, S.E.; Bennison, D.J.; Corrigan, R.M. The (p)PpGpp-Binding GTPase Era Promotes RRNA Processing and Cold Adaptation in Staphylococcus Aureus. PLoS Genet. 2019, 15, e1008346. [Google Scholar] [CrossRef] [PubMed]
- Kelliher, J.L.; Radin, J.N.; Kehl-Fie, T.E. PhoPR Contributes to Staphylococcus Aureus Growth during Phosphate Starvation and Pathogenesis in an Environment-Specific Manner. Infect. Immun. 2018, 86, e00371-18. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, F.; Szekat, C.; Josten, M.; Sahl, H.-G.; Bierbaum, G. Antibiotic-Induced Autoactivation of IS256 in Staphylococcus Aureus. Antimicrob. Agents Chemother. 2013, 57, 6381–6384. [Google Scholar] [CrossRef]
- Cue, D.; Lei, M.G.; Lee, C.Y. Activation of SarX by Rbf Is Required for Biofilm Formation and IcaADBC Expression in Staphylococcus Aureus. J. Bacteriol. 2013, 195, 1515–1524. [Google Scholar] [CrossRef]
- Schilcher, K.; Horswill, A.R. Staphylococcal Biofilm Development: Structure, Regulation, and Treatment Strategies. Microbiol. Mol. Biol. Rev. MMBR 2020, 84, e00026-19. [Google Scholar] [CrossRef]
- Luong, T.T.; Lei, M.G.; Lee, C.Y. Staphylococcus Aureus Rbf Activates Biofilm Formation In Vitro and Promotes Virulence in a Murine Foreign Body Infection Model. Infect. Immun. 2009, 77, 335–340. [Google Scholar] [CrossRef]
- Silveira, A.C.O.; Caierão, J.; Silva, C.I.; Anzai, E.K.; McCulloch, J.A.; d’Azevedo, P.A.; Sincero, T.C.M. Impact of Mutations in HVISA Isolates on Decreased Susceptibility to Vancomycin, through Population Analyses Profile—Area under Curve (PAP-AUC). Diagn. Microbiol. Infect. Dis. 2019, 95, 114854. [Google Scholar] [CrossRef]
- Salemi, R.; Zega, A.; Aguglia, E.; Lo Verde, F.; Pigola, G.; Stefani, S.; Cafiso, V. Balancing the Virulence and Antimicrobial Resistance in VISA DAP-R CA-MRSA Superbug. Antibiotics 2022, 11, 1159. [Google Scholar] [CrossRef] [PubMed]
- Kant, S.; Asthana, S.; Missiakas, D.; Pancholi, V. A Novel STK1-Targeted Small-Molecule as an “Antibiotic Resistance Breaker” against Multidrug-Resistant Staphylococcus Aureus. Sci. Rep. 2017, 7, 5067. [Google Scholar] [CrossRef] [PubMed]
- Blake, K.L.; Randall, C.P.; O’Neill, A.J. In Vitro Studies Indicate a High Resistance Potential for the Lantibiotic Nisin in Staphylococcus Aureus and Define a Genetic Basis for Nisin Resistance▿. Antimicrob. Agents Chemother. 2011, 55, 2362–2368. [Google Scholar] [CrossRef]
- Chan, Y.G.Y.; Frankel, M.B.; Missiakas, D.; Schneewind, O. SagB Glucosaminidase Is a Determinant of Staphylococcus Aureus Glycan Chain Length, Antibiotic Susceptibility, and Protein Secretion. J. Bacteriol. 2016, 198, 1123–1136. [Google Scholar] [CrossRef]
- Ishii, K.; Tabuchi, F.; Matsuo, M.; Tatsuno, K.; Sato, T.; Okazaki, M.; Hamamoto, H.; Matsumoto, Y.; Kaito, C.; Aoyagi, T.; et al. Phenotypic and Genomic Comparisons of Highly Vancomycin-Resistant Staphylococcus Aureus Strains Developed from Multiple Clinical MRSA Strains by in Vitro Mutagenesis. Sci. Rep. 2015, 5, 17092. [Google Scholar] [CrossRef]
- Santos-Beneit, F.; Ordóñez-Robles, M.; Martín, J.F. Glycopeptide Resistance: Links with Inorganic Phosphate Metabolism and Cell Envelope Stress. Biochem. Pharmacol. 2017, 133, 74–85. [Google Scholar] [CrossRef]
- Mechler, L.; Herbig, A.; Paprotka, K.; Fraunholz, M.; Nieselt, K.; Bertram, R. A Novel Point Mutation Promotes Growth Phase-Dependent Daptomycin Tolerance in Staphylococcus Aureus. Antimicrob. Agents Chemother. 2015, 59, 5366–5376. [Google Scholar] [CrossRef]
- Bakthavatchalam, Y.D.; Babu, P.; Munusamy, E.; Dwarakanathan, H.T.; Rupali, P.; Zervos, M.; John Victor, P.; Veeraraghavan, B. Genomic Insights on Heterogeneous Resistance to Vancomycin and Teicoplanin in Methicillin-Resistant Staphylococcus Aureus: A First Report from South India. PLoS ONE 2019, 14, e0227009. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Davis, M.H.; Peterson, J.; Solis-Lemus, C.; Satola, S.W.; Read, T.D. Effect of Genetic Background on the Evolution of Vancomycin-Intermediate Staphylococcus Aureus (VISA). PeerJ 2021, 9, e11764. [Google Scholar] [CrossRef]
- Myers, C.L.; Li, F.K.K.; Koo, B.-M.; El-Halfawy, O.M.; French, S.; Gross, C.A.; Strynadka, N.C.J.; Brown, E.D. Identification of Two Phosphate Starvation-Induced Wall Teichoic Acid Hydrolases Provides First Insights into the Degradative Pathway of a Key Bacterial Cell Wall Component. J. Biol. Chem. 2016, 291, 26066–26082. [Google Scholar] [CrossRef] [PubMed]
- Delauné, A.; Dubrac, S.; Blanchet, C.; Poupel, O.; Mäder, U.; Hiron, A.; Leduc, A.; Fitting, C.; Nicolas, P.; Cavaillon, J.-M.; et al. The WalKR System Controls Major Staphylococcal Virulence Genes and Is Involved in Triggering the Host Inflammatory Response. Infect. Immun. 2012, 80, 3438–3453. [Google Scholar] [CrossRef]
- Falord, M.; Mäder, U.; Hiron, A.; Débarbouillé, M.; Msadek, T. Investigation of the Staphylococcus Aureus GraSR Regulon Reveals Novel Links to Virulence, Stress Response and Cell Wall Signal Transduction Pathways. PLoS ONE 2011, 6, e21323. [Google Scholar] [CrossRef]
- Dubrac, S.; Msadek, T. Identification of Genes Controlled by the Essential YycG/YycF Two-Component System of Staphylococcus Aureus. J. Bacteriol. 2004, 186, 1175–1181. [Google Scholar] [CrossRef]
- Cameron, D.R.; Jiang, J.-H.; Kostoulias, X.; Foxwell, D.J.; Peleg, A.Y. Vancomycin Susceptibility in Methicillin-Resistant Staphylococcus Aureus Is Mediated by YycHI Activation of the WalRK Essential Two-Component Regulatory System. Sci. Rep. 2016, 6, 30823. [Google Scholar] [CrossRef]
- Cafiso, V.; Stracquadanio, S.; Lo Verde, F.; De Guidi, I.; Zega, A.; Pigola, G.; Stefani, S. Genomic and Long-Term Transcriptomic Imprints Related to the Daptomycin Mechanism of Action Occurring in Daptomycin- and Methicillin-Resistant Staphylococcus Aureus Under Daptomycin Exposure. Front. Microbiol. 2020, 11, 1893. [Google Scholar] [CrossRef]
- Su, M.; Satola, S.W.; Read, T.D. Genome-Based Prediction of Bacterial Antibiotic Resistance. J. Clin. Microbiol. 2019, 57, e01405-18. [Google Scholar] [CrossRef]
- Kim, J.I.; Maguire, F.; Tsang, K.K.; Gouliouris, T.; Peacock, S.J.; McAllister, T.A.; McArthur, A.G.; Beiko, R.G. Machine Learning for Antimicrobial Resistance Prediction: Current Practice, Limitations, and Clinical Perspective. Clin. Microbiol. Rev. 2022, 35, e0017921. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 22nd ed.; M100-S22; CLSI: San Antonio, TX, USA.
- Katz, B.D.; Luperchio, S.A.; Thorne, G.M. Detection of Daptomycin-Nonsusceptible Strains Using the Neo-Sensitab Prediffusion Method. Diagn. Microbiol. Infect. Dis. 2008, 61, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Rosco Diagnostica A/S Supplement User’s Guide. Neo-SensitabsTM Susceptibility Testing, Supplement 2010. Available online: http://www.rosco.dk/ (accessed on 22 December 2022).
- Wootton, M.; Howe, R.A.; Hillman, R.; Walsh, T.R.; Bennett, P.M.; MacGowan, A.P. A Modified Population Analysis Profile (PAP) Method to Detect Hetero-Resistance to Vancomycin in Staphylococcus Aureus in a UK Hospital. J. Antimicrob. Chemother. 2001, 47, 399–403. [Google Scholar] [CrossRef]
- Andrews, S. FastQC a Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 22 December 2022).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, K.D.; Tatusova, T.; Brown, G.R.; Maglott, D.R. NCBI Reference Sequences (RefSeq): Current Status, New Features and Genome Annotation Policy. Nucleic Acids Res. 2012, 40, D130–D135. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Snippy: Rapid Haploid Variant Calling and Core Genome Alignment. Available online: https://github.com/tseemann/snippy (accessed on 22 December 2022).
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An Integrated Platform for Visualization and Analysis of High-Throughput Sequence-Based Experimental Data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H.; Hasman, H.; Larsen, J.; Stegger, M.; Johannesen, T.B.; Allesøe, R.L.; Lemvigh, C.K.; Aarestrup, F.M.; Lund, O.; Larsen, A.R. SCCmecFinder, a Web-Based Tool for Typing of Staphylococcal Cassette Chromosome Mec in Staphylococcus Aureus Using Whole-Genome Sequence Data. mSphere 2018, 3, e00612-17. [Google Scholar] [CrossRef]
- Hunt, M.; Mather, A.E.; Sánchez-Busó, L.; Page, A.J.; Parkhill, J.; Keane, J.A.; Harris, S.R. ARIBA: Rapid Antimicrobial Resistance Genotyping Directly from Sequencing Reads. Microb. Genomics 2017, 3, e000131. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A Novel Web Tool for WGS-Based Detection of Antimicrobial Resistance Associated with Chromosomal Point Mutations in Bacterial Pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and Model-Centric Curation of the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.-M. ARG-ANNOT, a New Bioinformatic Tool to Discover Antibiotic Resistance Genes in Bacterial Genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef]
- Harkins, C.P.; Pichon, B.; Doumith, M.; Parkhill, J.; Westh, H.; Tomasz, A.; de Lencastre, H.; Bentley, S.D.; Kearns, A.M.; Holden, M.T.G. Methicillin-Resistant Staphylococcus Aureus Emerged Long before the Introduction of Methicillin into Clinical Practice. Genome Biol. 2017, 18, 130. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids Using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Goerke, C.; Pantucek, R.; Holtfreter, S.; Schulte, B.; Zink, M.; Grumann, D.; Bröker, B.M.; Doskar, J.; Wolz, C. Diversity of Prophages in Dominant Staphylococcus Aureus Clonal Lineages. J. Bacteriol. 2009, 191, 3462–3468. [Google Scholar] [CrossRef] [PubMed]
- Subedi, A.; Ubeda, C.; Adhikari, R.P.; Penadés, J.R.; Novick, R.P. Sequence Analysis Reveals Genetic Exchanges and Intraspecific Spread of SaPI2, a Pathogenicity Island Involved in Menstrual Toxic Shock. Microbiology 2007, 153, 3235–3245. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.D.; Bishop, B.; Wright, M.S. Quantitative Assessment of Insertion Sequence Impact on Bacterial Genome Architecture. Microb. Genomics 2016, 2, e000062. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, C.L.M.; Chooi, Y.-H. Clinker & Clustermap. Js: Automatic Generation of Gene Cluster Comparison Figures. Bioinformatics 2021, btab007. [Google Scholar] [CrossRef]
- Carver, T.J.; Rutherford, K.M.; Berriman, M.; Rajandream, M.-A.; Barrell, B.G.; Parkhill, J. ACT: The Artemis Comparison Tool. Bioinformatics 2005, 21, 3422–3423. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ 1997. Available online: https://imagej.nih.gov/ij/ (accessed on 22 December 2022).
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of Biofilm in Microtiter Plates: Overview of Testing Conditions and Practical Recommendations for Assessment of Biofilm Production by Staphylococci. APMIS Acta Pathol. Microbiol. Immunol. Scand. 2007, 115, 891–899. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| D1 | D2 | D23C9 | D2P11 | Reference | ||
|---|---|---|---|---|---|---|
| MIC (µg/mL) | VAN | 0.5 | 1 | 4 | 8 | [24] |
| OXA | 128 | 2 | 8 | 32 | [24] | |
| DAP | 0.094 | 0.38 | 0.5 | 1 | This Study | |
| DAP Pre-diffusion method (mm) | 30 | 30 | 22 | 18 | This Study | |
| VAN PAP-AUC ratio | 0.98 | 1.48 | 2.81 | 5.7 | [24] | |
| Phenotype | hVISA | hVISA | VISA | VISA | [24] | |
| Assembly | D1 | D2 | D2P11 | D23C9 |
|---|---|---|---|---|
| Assembly accession | JAPZAH000000000 | JAPZAI000000000 | JAPZAJ000000000 | JAPZAK000000000 |
| Genome coverage | 286.77 | 246.87 | 277.45 | 256.49 |
| # Contigs (≥0 bp) | 45 | 45 | 35 | 47 |
| # Contigs (≥1000 bp) | 38 | 38 | 31 | 40 |
| Total length (≥0 bp) | 2,795,159 | 2,795,155 | 2,785,355 | 2,787,646 |
| Total length (≥1000 bp) | 2,789,975 | 2,789,971 | 2,782,204 | 2,782,459 |
| # Contigs | 45 | 45 | 35 | 47 |
| Largest contig | 567,081 | 567,084 | 567,084 | 419,127 |
| Total length | 2,795,159 | 2,795,155 | 2,785,355 | 2,787,646 |
| GC (%) | 32.79 | 32.79 | 32.8 | 32.81 |
| N50 | 334,617 | 334,619 | 337,832 | 148,220 |
| N90 | 36,723 | 36,723 | 39,437 | 33,617 |
| L50 | 4 | 4 | 4 | 5 |
| L90 | 17 | 17 | 16 | 19 |
| # N’s per 100 kbp | 0 | 0 | 0 | 0 |
| Feature | |
|---|---|
| MLST | ST100 |
| SCCmec type | NT |
| AMR determinants | aac(6′)-aph(2″), blaZ, mecA, rpoB H481N |
| Insertion sequences | IS256, IS1181, IS431, ISSau6 |
| Transposon | Tn4001 |
| RM systems | S.Sau N315 I–M.Sau N315 I, S.Sau N315 II–M.Sau N315 II, SauUSI |
| Plasmidic rep genes | rep21.13_SAP101A (GQ900495.1), rep20.3_pTW20 (FN433597.1) |
| Chromosome Position | Gene Name | Product | Predicted Amino Acid Change |
|---|---|---|---|
| 46,301 | mecA | Penicillin-binding protein 2 prime | Gly246Glu |
| 297,502 | tarF_1 | CDP-glycerol:poly(glycerophosphate) glycerophosphotransferase | Asn236Ser |
| 581,060 | rpoB | DNA-directed RNA polymerase subunit beta | His481Asn |
| 583,428 | rpoC | DNA-directed RNA polymerase beta’ chain protein | Pro41Ala |
| 1,595,731 | rpoD | RNA polymerase sigma-70 factor RpoD | Val253Ile |
| 687,696 | tagG | Teichoic acid ABC superfamily ATP-binding cassette transporter | Val220Ala |
| 732,508 | cydD | ABC superfamily ATP-binding cassette transporter | Asn174Asp |
| 902,157 | dltD | D-alanine lipoteichoic acid and wall teichoic acid esterification secreted protein | Ile264Lys |
| 993,796 | murE | UDP-N-acetylmuramoyl-L-alanyl-D-glutamate--2 | Ala436Ser |
| 1,157,903 | ftsL | Cell division and chromosome partitioning protein | Asp78Asn |
| 1,203,919 | pknB_2(stk1) | Non-specific serine/threonine protein kinase | Lys512Asn |
| 1,487,245 | pbp2 | Glycosyl transferase family protein | Cys197Tyr |
| 1,487,506 | pbp2 | Glycosyl transferase family protein | Thr284Ile |
| 1,758,442 | ezrA | Septation ring formation regulator | Val545Ala |
| 2,009,774–2,009,775 | ami | N-acetylmuramoyl-L-alanine amidase | Ala250Val |
| 2,010,074 | ami | N-acetylmuramoyl-L-alanine amidase | Asn150Lys |
| 2,010,079 | ami | N-acetylmuramoyl-L-alanine amidase | Ile149Val |
| 2,138,921 | murF | Putative UDP-N-acetylmuramoylalanyl-D-glutamyl-2 | Ser126Asn |
| 2,171,885 | rho | Putative methicillin resistance expression factor | Ile48Leu |
| 2,412,317 | tcaA | Teicoplanin resistance-associated membrane protein TcaA protein | Leu218Pro |
| Chromosome Position | Gene Name | Product | Type | Predicted Aminoacid Change | |||
|---|---|---|---|---|---|---|---|
| D1 | D2 | D23C9 | D2P11 | ||||
| 1,506,816 | Intergenic (upstream aroC) | - | SNP | C63217A * | wt | wt | wt |
| 1,003,592 | Intergenic (upstream comK) | - | SNP | T19402C * | wt | wt | wt |
| 170,209 | cap5D | Capsular polysaccharide biosynthesis protein Cap5D | SNP | Thr215Ile | wt | wt | wt |
| 1,201,763 | stp1 | Protein phosphatase 2C domain-containing protein | INDEL | Gly41_Lys43dup | wt | wt | wt |
| 2,713,713 | nsaS/braS | Integral membrane sensor signal transduction histidine kinase | INDEL | Gln63del | wt | wt | wt |
| 1,832,949 | sagB | Beta-N-acetylglucosaminidase | INDEL | Wt | His142fs | His142fs | His142fs |
| 12,058(NODE_18) *2 | SAD1_02353 | Bacteriophage tail tape measure protein | SNP | Wt | Asp1737Gly | Asp1737Gly | Asp1737Gly |
| 27,119(NODE 19) *2 | SAD1_02404 | GNAT family acetyl transferase | SNP | Wt | Tyr158Tyr | Tyr158Tyr | Tyr158Tyr |
| 24,307(NODE 21) *2 | ccrB | Cassette chromosome recombinase B | SNP | Wt | Asn10Ser | Asn10Ser | Asn10Ser |
| 1,728,074 | phoR | Alkaline phosphatase synthesis sensor protein PhoR | SNP | Wt | wt | Arg200 * STOP | wt |
| 1,603,348 | era | GTP-binding protein Era | INDEL | Wt | wt | wt | Asn81fs |
| IS | D1 | D2 | D23C9 | D2P11 |
|---|---|---|---|---|
| IS256 | 14 | 14 | 17 | 13 |
| IS1181 | 7 + 1 * | 7 + 1 * | 7 + 1 * | 7 + 1 * |
| IS1182 | 0 | 0 | 0 | 0 |
| IS1272 | 1 * | 1* | 1 * | 1 * |
| IS431 | 1 | 1 | 1 | 1 |
| ISSau1 | 0 | 0 | 0 | 0 |
| ISSau2 | 3 * | 3 * | 3 * | 3 * |
| ISSau3 | 10 * | 10 * | 10 * | 10 * |
| ISSau4 | 0 | 0 | 0 | 0 |
| ISSau5 | 1 * | 1 * | 1 * | 1 * |
| ISSau6 | 6 | 6 | 6 | 6 |
| ISSau8 | 2 * | 2 * | 2 * | 2 * |
| ISSau9 | 0 | 0 | 0 | 0 |
| ISSep1 | 1 * | 1 * | 1 * | 1 * |
| ISSep2 | 1 * | 1 * | 1 * | 1 * |
| ISSep3 | 0 | 0 | 0 | 0 |
| ISSha1 | 0 | 0 | 0 | 0 |
| Genomic Location | Type | D1 | D2 | D23C9 | D2P11 |
|---|---|---|---|---|---|
| mecR1 | Disrupting gene | + | + | + | + |
| SA0142 (hypothetical protein) | Disrupting gene | + | + | + | + |
| SA0084 (hypothetical protein)—SA0085 (tRNA dihidrouridine sintase) | Intergenic | + | + | + | + |
| SA0516 (tRNA specific adenosine deaminase)—SA0517 (ABC superfamily ATP-binding cassette transporter) | Intergenic | + | + | + | + |
| vraG (ABC transporter permease)—SA0618 (pitR, putative phosphate uptake regulator) ** | Intergenic | + | + | + * | + * |
| SA0621 (integral membrane protein) **—rbf (AraC type transcription regulator) ** | Intergenic | + | + | - | - |
| sarX (staphylococcal accessory regulator protein X) **—SA0624 (putative transcriptional regulatory protein) | Intergenic | + | + | + * | + * |
| SA0625 (hypothetical protein)—SA0626 (hypothetical protein) | Intergenic | - | - | + | - |
| SA0742 (clfA, clumping factor A) | Disrupting gene | - | - | + | - |
| SA0954 (hypothetical protein)—SA0955 (hypothetical protein) | Intergenic | + | + | + | + |
| SA0185 (putative membrane protein YfhO)—rbgA (GTP-binding protein) | Intergenic | + | + | + | + |
| SA1176 (hypothetical protein)—tkt (transketolase) | Intergenic | + | + | + | + |
| SA1232 (lysA, diaminopimelate decarboxylase)—SA1233 (msaC, modulator of sarA) *** | Disrupting gene | + | + | + | + |
| SA1639 (hypothetical protein) | Disrupting gene | + | + | + | + |
| SA1648 (enterotoxin seO)—tRNAser | Intergenic | + | + | + | + |
| agrB (accesory gene regulator B) | Disrupting gene | - | - | + | - |
| SA2019 (hypothetical protein)—SA2010 (hypothetical protein) | Intergenic | + | + | + | + |
| SA2414 (hypothetical protein)—SA2415 (braE, ABC superfamily ATP-binding cassette transporter membrane protein) | Intergenic | - | - | + | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Gregorio, S.; Haim, M.S.; Famiglietti, Á.M.R.; Di Conza, J.; Mollerach, M. Comparative Genomics Identifies Novel Genetic Changes Associated with Oxacillin, Vancomycin and Daptomycin Susceptibility in ST100 Methicillin-Resistant Staphylococcus aureus. Antibiotics 2023, 12, 372. https://doi.org/10.3390/antibiotics12020372
Di Gregorio S, Haim MS, Famiglietti ÁMR, Di Conza J, Mollerach M. Comparative Genomics Identifies Novel Genetic Changes Associated with Oxacillin, Vancomycin and Daptomycin Susceptibility in ST100 Methicillin-Resistant Staphylococcus aureus. Antibiotics. 2023; 12(2):372. https://doi.org/10.3390/antibiotics12020372
Chicago/Turabian StyleDi Gregorio, Sabrina, María Sol Haim, Ángela María Rosa Famiglietti, José Di Conza, and Marta Mollerach. 2023. "Comparative Genomics Identifies Novel Genetic Changes Associated with Oxacillin, Vancomycin and Daptomycin Susceptibility in ST100 Methicillin-Resistant Staphylococcus aureus" Antibiotics 12, no. 2: 372. https://doi.org/10.3390/antibiotics12020372
APA StyleDi Gregorio, S., Haim, M. S., Famiglietti, Á. M. R., Di Conza, J., & Mollerach, M. (2023). Comparative Genomics Identifies Novel Genetic Changes Associated with Oxacillin, Vancomycin and Daptomycin Susceptibility in ST100 Methicillin-Resistant Staphylococcus aureus. Antibiotics, 12(2), 372. https://doi.org/10.3390/antibiotics12020372