
Comparison of Oral Microbial Composition and Determinants Encoding Antimicrobial Resistance in Dogs and Their Owners



Abstract
:1. Introduction
2. Results
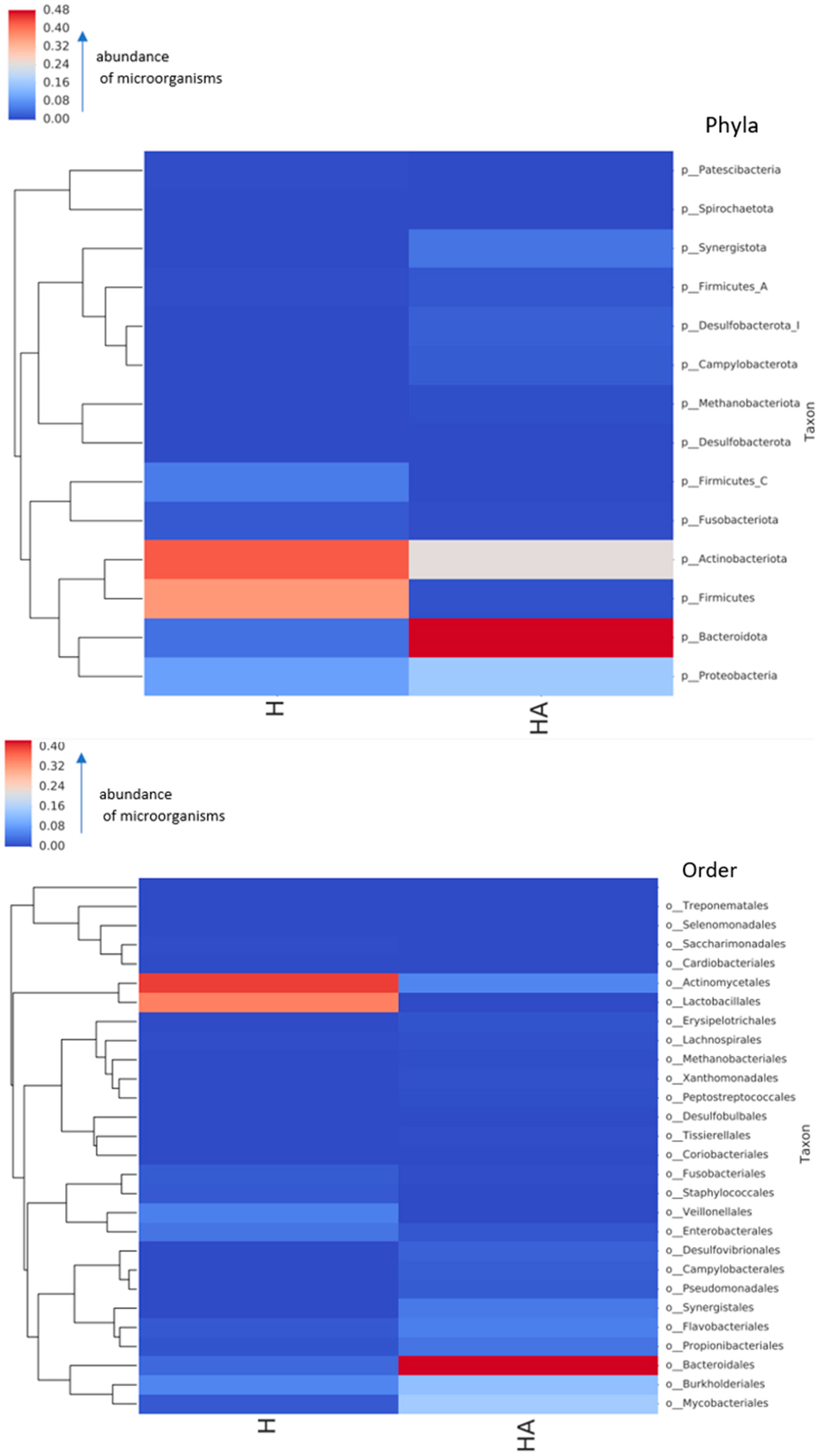
2.1. Microbial Composition of Oral Microbiomes of Pets and Their Owners
2.2. Prevalence of AMR Genes
3. Discussion
3.1. Human Microbiome
3.2. Dog Microbiome
3.3. Similarity of Human and Dog Microbiomes and the Influence of Keeping Dogs for Microbial Composition in Pet Owners
3.4. Zoonotic Potential of the AMR Genes
4. Materials and Methods
4.1. Animals and Humans Involved in the Study
4.2. Sampling Procedure
4.3. Microbial Profiling and Detection of AMR
4.4. Statistical and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rahman, M.T.; Sobur, M.A.; Islam, M.S.; Ievy, S.; Hossain, M.J.; El Zowalaty, M.E.; Rahman, A.T.; Ashour, H.M. Zoonotic diseases: Etiology, impact and control. Microorganisms 2020, 8, 1405. [Google Scholar] [CrossRef] [PubMed]
- Welburn, S.C.; Beange, I.; Ducrotoy, M.J.; Okello, A.L. The neglected zoonoses—The case for integrated control and advocacy. Clin. Microbiol. Infect. 2015, 21, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Cleaveland, S.; Sharp, J.; Abela-Ridder, B.; Allan, K.J.; Buza, J.; Crump, J.A.; Davis, A.; Del Rio Vilas, V.J.; de Glanville, W.A.; Kazwala, R.R.; et al. One Health contributions towards more effective and equitable approaches to health in low- and middle-income countries. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160168. [Google Scholar] [CrossRef] [PubMed]
- Ceric, O.; Tyson, G.H.; Goodman, L.B.; Mitchell, P.K.; Zhang, Y.; Prarat, M.; Cui, J.; Peak, L.; Scaria, J.; Antony, L.; et al. Enhancing the One Health initiative by using whole genome sequencing to monitor antimicrobial resistance of animal pathogens: Vet-LIRN collaborative project with veterinary diagnostic laboratories in United States and Canada. BMC Vet. Res. 2019, 15, 130. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Jeggo, M. The One Health Approach—Why is it so important? Trop. Med. Infect. Dis. 2019, 4, 88. [Google Scholar] [CrossRef]
- Nazir, M.A. Prevalence of periodontal disease, its association with systemic diseases and prevention. Int. J. Health Sci. 2017, 11, 72–80. [Google Scholar]
- Kačírová, J.; Maďari, A.; Mucha, R.; Fecskeova, L.K.; Mujakic, I.; Kobližek, M.; Nemcova, R.; Mad’ar, M. Study of microbiocenosis of canine dental biofilms. Sci. Rep. 2021, 11, 19776. [Google Scholar] [CrossRef]
- Shearer, P. Periodontal Literature Review. BARK. 2010, pp. 1–8. Available online: https://www.banfield.com/getmedia/fca67363-7362-4bb0-9824-50905f7ca817/48fdbdb1-8ff0-40b6-8d7f-3e8fac47de3a-pdf0 (accessed on 1 June 2023).
- Yamasaki, Y.; Nomura, R.; Nakano, K.; Naka, S.; Motsumoto-Nakano, M.; Asai, F.; Ooshima, T. Distribution of periodontopathic bacterial species in dogs and their owners. Arch. Oral Biol. 2012, 13, 1183–1188. [Google Scholar] [CrossRef]
- Bai, Y.; Song, P.; Shen, Z.; Shi, H.; Jiang, Z.; Lin, J.; Jin, Y. Porphyromonas gulae infection in canines, pet owners and veterinarians in China: An epidemiological study and risk factor analysis. One Health Adv. 2023, 1, 9. [Google Scholar] [CrossRef]
- Nises, J.; Rosander, A.; Pettersson, A.; Backhans, A. The occurrence of Treponema spp. in gingival plaque from dogs with varying degree of periodontal disease. PLoS ONE 2018, 13, e0201888. [Google Scholar] [CrossRef]
- Maruyama, N.; Mori, A.; Shono, S.; Oda, H.; Sako, T. Evaluation of changes in periodontal bacteria in healthy dogs over 6 months using quantitative real-time PCR. Pol. J. Vet. Sci. 2018, 21, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Ghasemzadeh, I.; Namazi, S.H. Review of bacterial and viral zoonotic infections transmitted by dogs. J. Med. Life 2015, 8, 1–5. [Google Scholar] [PubMed]
- Dewhirst, F.E.; Klein, E.A.; Thompson, E.C.; Blanton, J.M.; Chen, T.; Milella, L.; Buckley, C.M.; Davis, I.J.; Bennett, M.L.; Marshall-Jones, Z.V. The canine oral microbiome. PLoS ONE 2012, 7, e36067. [Google Scholar] [CrossRef]
- Buchan, B.W.; Ledeboer, N.A. Emerging technologies for the clinical microbiology laboratory. Clin. Microbiol. Rev. 2014, 27, 783–822. [Google Scholar] [CrossRef] [PubMed]
- Rosselli, R.; Romoli, O.; Vitulo, N.; Vezzi, A.; Campanaro, S.; de Pascele, F.; Schiavon, R.; Tiarca, M.; Poletto, F.; Concheri, G.; et al. Direct 16S rRNA-seq from bacterial communities: A PCR-independent approach to simultaneously assess microbial diversity and functional activity potential of each taxon. Sci. Rep. 2016, 6, 32165. [Google Scholar] [CrossRef]
- Hassler, H.B.; Probert, B.; Moore, C.; Lawson, E.; Jackson, R.W.; Russell, B.T.; Richards, V.P. Phylogenies of the 16S rRNA gene and its hypervariable regions lack concordance with core genome phylogenies. Microbiome 2022, 10, 104. [Google Scholar] [CrossRef]
- Matsuo, Y.; Komiya, S.; Yasumizu, Y.; Yasuoka, Y.; Mizushima, L.; Takagi, T.; Kryukov, K.; Fukuda, A.; Morimoto, Y.; Naito, Y.; et al. Full-length 16S rRNA gene amplicon analysis of human gut microbiota using MinION™ nanopore sequencing confers species-level resolution. BMC Microbiol. 2021, 21, 35. [Google Scholar] [CrossRef]
- Noyes, N.R.; Weinroth, M.E.; Parker, J.K.; Dean, C.J.; Lakin, S.M.; Raymond, R.A.; Rovira, P.; Doster, E.; Abdo, Z.; Martin, J.N.; et al. Enrichment allows identification of diverse, rare elements in metagenomic resistome-virulome sequencing. Microbiome 2017, 5, 142. [Google Scholar] [CrossRef]
- Deurenberg, R.H.; Bathoorn, E.; Chlebowicz, M.A.; Couto, N.; Ferdous, M.; García-Cobos, S.; Kooistra-Smid, A.M.D.; Raangs, E.C.; Rosema, S.; Veloo, A.C.M.; et al. Application of next generation sequencing in clinical microbiology and infection prevention. J. Biotechnol. 2017, 243, 16–24. [Google Scholar] [CrossRef]
- Tóth, A.G.; Tóth, I.; Rózsa, B.; Dubecz, A.; Patai, Á.V.; Németh, T.; Kaplan, S.; Kovács, E.G.; Makrai, L.; Solymosi, N. Canine saliva as a possible source of antimicrobial resistance genes. Antibiotics 2022, 27, 1490. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Cong, Y. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell. Mol. Immunol. 2021, 18, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Riggio, M.P.; Lennon, A.; Taylor, D.J.; Bennett, D. Molecular identification of bacteria associated with canine periodontal disease. Vet. Microbiol. 2011, 150, 394–400. [Google Scholar] [CrossRef]
- Gadê-Neto, C.R.; Rodrigues, R.R.; Louzada, L.M.; Arruda-Vasconcelos, R.; Teixeira, F.B.; Viana Casarin, R.C.; Gomes, B.P. Microbiota of periodontal pockets and root canals in induced experimental periodontal disease in dogs. J. Investig. Clin. Dent. 2019, 10, e12439. [Google Scholar] [CrossRef]
- Özavci, V.; Erbas, G.; Parin, U.; Yüksel, H.T.; Kirkan, Ş. Molecular detection of feline and canine periodontal pathogens. Vet. Anim. Sci. 2019, 8, 100069. [Google Scholar] [CrossRef] [PubMed]
- Santibáñez, R.; Rodríguez-Salas, C.; Flores-Yáñez, C.; Garrido, D.; Thomson, P. Assessment of changes in the oral microbiome that occur in dogs with periodontal disease. Vet. Sci. 2021, 8, 291. [Google Scholar] [CrossRef]
- Ruparell, A.; Inui, T.; Staunton, R.; Wallis, C.; Deusch, S.; Holcombe, L.J. The canine oral microbiome: Variation in bacterial populations across different niches. BMC Microbiol. 2020, 20, 42. [Google Scholar] [CrossRef]
- Kačírová, J.; Sondorová, M.; Maďari, A.; Styková, E.; Mucha, R.; Nemcová, R.; Marečáková, N.; Farbáková, J.; Maďar, M. Detection of periodontal pathogens from dental plaques of dogs with and without periodontal disease. Pathogens 2022, 11, 480. [Google Scholar] [CrossRef]
- Elliott, D.R.; Wilson, M.; Buckley, C.M.; Spratt, D.A. Cultivable oral microbiota of domestic dogs. J. Clin. Microbiol. 2005, 43, 5470–5476. [Google Scholar] [CrossRef]
- Bell, S.E.; Nash, A.K.; Zanghi, B.M.; Otto, C.M.; Perry, E.B. An assessment of the stability of the canine oral microbiota after probiotic administration in healthy dogs over time. Front. Vet. Sci. 2020, 7, 616. [Google Scholar] [CrossRef]
- Oba, P.M.; Carroll, M.Q.; Alexander, C.; Somrak, A.J.; Keating, S.C.J.; Sage, A.M.; Swanson, K.S. Dental chews positively shift the oral microbiota of adult dogs. J. Anim. Sci. 2021, 99, 100. [Google Scholar] [CrossRef] [PubMed]
- The Human Microbiome Project Consortium. A framework for human microbiome research. Nature 2012, 486, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Sampaio-Maia, B.; Monteiro-Silva, F. Acquisition and maturation of oral microbiome throughout childhood: An update. Dent. Res. J. 2014, 11, 291–301. [Google Scholar]
- Deo, P.N.; Deshmukh, R. Oral microbiome: Unveiling the fundamentals. J. Oral Maxillofac. Pathol. 2019, 23, 122–128. [Google Scholar] [CrossRef]
- Marsh, P.D. Role of the oral microflora in health. Microb. Ecol. Health Dis. 2009, 12, 130–137. [Google Scholar]
- Garrigues, Q.; Apper, E.; Chastant, S.; Mila, H. Gut microbiota development in the growing dog: A dynamic process influenced by maternal, environmental and host factors. Front. Vet. Sci. 2022, 9, 964649. [Google Scholar] [CrossRef]
- Dunn, A.B.; Jordan, S.; Baker, B.J.; Carlson, N.S. The maternal infant microbiome: Considerations for labor and birth. MCN Am. J. Matern. Child Nurs. 2017, 42, 318–325. [Google Scholar] [CrossRef]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef]
- Marsh, P. Contemporary perspective on plaque control. Br. Dent. J. 2012, 212, 601–606. [Google Scholar] [CrossRef]
- Willis, J.R.; Gabaldón, T. The human oral microbiome in health and disease: From sequences to ecosystems. Microorganisms 2020, 8, 308. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Yang, X.; Li, C.; Song, Z. The oral microbiota: Community composition, influencing factors, pathogenesis, and interventions. Front. Microbiol. 2022, 13, 895537. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Luo, J.; Zhang, P.; Howell, K. The salivary microbiome shows a high prevalence of core bacterial members yet variability across human populations. Npj Biofilms Microbiomes 2022, 8, 85. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M. The Role of Streptococci in Oral Periodontitis: Growth and Disruption of Yellow Complex Bacteria. Master’s Thesis, Wilfrid Laurier University, Waterloo, ON, Canada, 2020. Available online: https://scholars.wlu.ca/etd/2336 (accessed on 1 August 2023).
- Matera, G.; Muto, V.; Vinci, M.; Zicca, E.; Abdollahi-Roodsaz, S.; van de Veerdonk, F.L.; Kullberg, B.-J.; Liberto, M.C.; van der Meer, J.W.; Foca, A. Receptor recognition of and immune intracellular pathways for Veillonella parvula lipopolysaccharide. Clin. Vaccine Immunol. 2009, 16, 1804–1809. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef]
- Fisher, R.G.; Denison, M.R. Veillonella parvula bacteremia without an underlying source. J. Clin. Microbiol. 1996, 34, 3235–3236. [Google Scholar] [CrossRef] [PubMed]
- Oba, P.M.; Sieja, K.M.; Kreating, S.C.; Hristova, T.; Somrak, A.J.; Swanson, K.S. Oral microbiota populations of adult dog consuming wet or dry foods. J. Anim. Sci. 2022, 100, 200. [Google Scholar] [CrossRef]
- Kačírová, J.; Maďar, M.; Štrkolcová, G.; Maďari, A.; Nemcová, R. Dental Biofilm as Etiological Agent of Canine Periodontal Disease. In Bacterial Biofilms; Dincer, S., Ozdenefe, M.S., Arkut, A., Eds.; IntechOpen: Rijeka, Croatia, 2020. [Google Scholar] [CrossRef]
- O’Neill, D.G.; Mitchell, C.E.; Humphrey, J.; Church, D.B.; Brodbelt, D.C.; Pegram, C. Epidemiology of periodontal disease in dogs in the UK primary-care veterinary setting. J. Small Anim. Pract. 2021, 62, 1051–1061. [Google Scholar] [CrossRef]
- Wallis, C.; Saito, E.K.; Salt, C.; Holcombe, L.J.; Desforges, N.G. Association of periodontal disease with breed size, breed, weight, and age in pure-bred client-owned dogs in the United States. Vet. J. 2021, 275, 105717. [Google Scholar] [CrossRef]
- Dos Santos, P.J.D.; Cunha, E.; Nunes, T.; Tavares, L.; Oliveira, M. Relation between periodontal disease and systemic diseases in dogs. Res. Vet. Sci. 2019, 125, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Oba, P.M.; Carroll, M.Q.; Alexander, C.; Valentine, H.; Somrak, A.J.; Keating, S.C.J.; Sage, A.M.; Swanson, K.S. Microbiota populations in supragingival plaque, subgingival plaque, and saliva habitats of adult dogs. Anim. Microbiome 2021, 7, 38. [Google Scholar] [CrossRef]
- Oh, C.; Lee, K.; Cheong, Y.; Lee, S.W.; Park, S.Y.; Song, C.S.; Choi, I.S.; Lee, J.B. Comparison of the oral microbiomes of canines and their owners using next-generation sequencing. PLoS ONE 2015, 10, e0131468. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Cui, Z.; Fan, P.; Du, G. Effects of dog ownership on the gut microbiota of elderly owners. PLoS ONE 2022, 17, e0278105. [Google Scholar] [CrossRef] [PubMed]
- Holcombe, L.J.; Patel, N.; Colyer, A.; Deusch, O.; O’Flynn, C.; Harris, S. Early canine plaque biofilms: Characterization of key bacterial interactions involved in initial colonization of enamel. PLoS ONE 2014, 9, e113744. [Google Scholar] [CrossRef] [PubMed]
- Wipler, J.; Čermáková, Z.; Hanzálek, T.; Horáková, H.; Žemličková, H. Sharing bacterial microbiota between owners and their pets (dogs, cats). Klinicka Mikrobiologie a Infekcni Lekarstvi 2017, 23, 48–57. [Google Scholar]
- Enlund, K.B.; Brunius, C.; Hanson, J.; Hagman, R.; Höglund, O.V.; Gustås, P.; Pettersson, A. Dental home care in dogs-a questionnaire study among Swedish dog owners, veterinarians and veterinary nurses. BMC Vet. Res. 2020, 16, 90. [Google Scholar] [CrossRef]
- Iwahara, K.; Kuriyama, T.; Shimura, S.; Williams, D.W.; Yanagisawa, M.; Nakagawa, K.; Karasawa, T. Detection of cfxA and cfxA2, the beta-lactamase genes of Prevotella spp., in clinical samples from dentoalveolar infection by real-time PCR. J. Clin. Microbiol. 2006, 44, 172–176. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef]
- Melville, C.M.; Scott, K.P.; Mercer, D.K.; Flint, H.J. Novel tetracycline resistance gene, tet(32), in the Clostridium-related human colonic anaerobe K10 and its transmission in vitro to the rumen anaerobe Butyrivibrio fibrisolvens. Antimicrob. Agents Chemother. 2001, 45, 3246–3249. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, I.; Sakellari, D.; Spala, A.; Arsenakis, M.; Konstantinidis, A. Prevalence of tetM, tetQ, nim and blaTEM genes in the oral cavities of Greek subjects: A pilot study. J. Clin. Periodontol. 2009, 36, 569–574. [Google Scholar] [CrossRef]
- Domínguez-Pérez, R.A.; De la Torre-Luna, R.; Ahumada-Cantillano, M.; Vázquez-Garcidueñas, M.S.; Pérez-Serrano, R.M.; Martínez-Martínez, R.E.; Guillen-Nepita, A.N. Detection of the antibiotic resistance genes blaTem-1, cfxA, tetQ, tetM, tetW, and ermC in endodontic infections of a Mexican population. J. Glob. Antimicrob. Resist. 2018, 15, 20–24. [Google Scholar] [CrossRef]
- Pérez-Serrano, R.M.; Domínguez-Pérez, R.A.; Ayala-Herrera, J.L.; Luna-Jaramillo, A.E.; de Larrea, G.Z.; Solís-Sainz, J.C.; García-Solís, P.; Loyola-Rodríguez, J.P. Dental plaque microbiota of pet owners and their dogs as a shared source and reservoir of antimicrobial resistance genes. J. Glob. Antimicrob. Res. 2020, 21, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Hao, J.; Yang, J.; Tong, C.; Xie, L.; Xiao, D.; Zeng, Z.; Xiong, W. The Co-occurrence of Antibiotic Resistance Genes between Dogs and Their Owners in Families. iMeta 2022, 1, e21. [Google Scholar] [CrossRef]
- Pomba, C.; Rantala, M.; Greko, C.; Baptiste, K.E.; Catry, B.; van Duijkeren, E.; Mateus, A.; Moreno, M.A.; Pyörälä, S.; Ružauskas, M.; et al. Public health risk of antimicrobial resistance transfer from companion animals. J. Antimicrob. Chemother. 2017, 72, 957–968. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Brown, C.T.; Irber, L. sourmash: A library for MinHash sketching of DNA. J. Open Source Softw. 2016, 1, 27. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 73, 35–336. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Z Score Calculator for 2 Population Proportions. Social Science Statistics. Available online: https://www.socscistatistics.com/tests/ztest/default.aspx (accessed on 20 August 2023).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Class | Resistance Determinants * | Number of Detected Genes in Samples | |
|---|---|---|---|
| Dogs | Owners | ||
| β-lactams | CfxA family class A broad-spectrum beta-lactamase | 35 | 145 |
| class A extended-spectrum beta-lactamase CfxA4 | 15 | 0 | |
| class A broad-spectrum beta-lactamase CfxA5 | 10 | 0 | |
| CSP family class A extended-spectrum beta-lactamase SPU-1 | 0 | 23 | |
| oxacillin-hydrolyzing class D beta-lactamase OXA-85 | 0 | 19 | |
| class A extended-spectrum beta-lactamase CfxA2 | 0 | 9 | |
| OXA-2 family class D extended-spectrum beta-lactamase OXA-539 | 7 | 0 | |
| class A beta-lactamase BRO-1 | 5 | 0 | |
| Aminoglycosides | ANT(3″)-Ia family aminoglycoside nucleotidyltransferases | 15 | 0 |
| aminoglycoside O-phosphotransferase APH(3″)-Ib | 8 | 0 | |
| aminoglycoside O-phosphotransferase APH(6)-Id | 17 | 0 | |
| Tetracyclines | tetracycline efflux ABC transporter TetAB subunit A | 19 | 0 |
| tetracycline efflux ABC transporter Tet(46) subunit B | 4 | 38 | |
| tetracycline efflux Na+/H+ antiporter family transporter Tet(35) | 6 | 0 | |
| tetracycline efflux ABC transporter Tet(46) | 0 | 44 | |
| tetracycline resistance ribosomal protection protein Tet(W) | 45 | 305 | |
| tetracycline resistance ribosomal protection protein Tet(M) | 0 | 366 | |
| tetracycline resistance ribosomal protection protein Tet(32) | 131 | 30 | |
| tetracycline efflux MFS transporter Tet(33) | 21 | 0 | |
| tetracycline resistance ribosomal protection protein Tet(Q) | 36 | 132 | |
| tetracycline resistance ribosomal protection protein Tet(O) | 12 | 8 | |
| tetracycline-inactivating monooxygenase Tet(X) | 6 | 0 | |
| tetracycline efflux MFS transporter Tet(Z) | 0 | 6 | |
| tetracycline resistance NADPH-dependent oxidoreductase Tet(37) | 0 | 6 | |
| Amphenicols | type A-3 chloramphenicol O-acetyltransferase CatIII | 9 | 0 |
| chloramphenicol efflux MFS transporter Cmx | 8 | 0 | |
| Macrolides | ABC-F type ribosomal protection protein Msr(D) | 0 | 1118 |
| 23S rRNA (adenine(2058)-N(6))-methyltransferase Erm(X) | 10 | 150 | |
| 23S rRNA (adenine(2058)-N(6))-methyltransferase Erm(39) | 0 | 11 | |
| 23S rRNA (adenine(2058)-N(6))-methyltransferase Erm(40) | 0 | 19 | |
| 23S rRNA (adenine(2058)-N(6))-methyltransferase Erm(F) | 0 | 25 | |
| macrolide efflux MFS transporter Mef(A) | 0 | 1089 | |
| Streptogramins, pleuromutilins and lincomycin | ABC-F type ribosomal protection protein Lsa(C) | 68 | 121 |
| ABC-F type ribosomal protection protein Lsa(B) | 6 | 0 | |
| Sulfonamides | sulfonamide-resistant dihydropteroate synthase Sul1 | 21 | 0 |
| sulfonamide-resistant dihydropteroate synthase Sul2 | 12 | 0 | |
| Other | quaternary ammonium compound efflux SMR transporter QacE delta 1 | 8 | 0 |
| Dogs | Owners | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Serial Number of Dogs | Age (Years) | Weight, kg | Breed | Diet | Living Place | Neutered | Age | Sex | Years of Keeping Dog(s) | Sleeps with Dog(s) |
| 1. | 0.5 | 26 | Afghan Hound | dry | City | no | 33 | female | 1 | yes |
| 2. | 15 | 37 | mixed | dry | City | no | 31 | male | 14 | no |
| 3. | 7 | 8 | mixed | mix | urban | yes | 26 | female | 7 | yes |
| 4. | 12 | 28 | mixed | mix | City | yes | 33 | female | 10 | yes |
| 5. | 4 | 25 | Retriever | dry | City | yes | 32 | female | 4 | no |
| 6. | 4 | 45 | German Shepherd | mix | urban | no | 27 | male | 4 | yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šakarnytė, L.; Šiugždinienė, R.; Žymantienė, J.; Ruzauskas, M. Comparison of Oral Microbial Composition and Determinants Encoding Antimicrobial Resistance in Dogs and Their Owners. Antibiotics 2023, 12, 1554. https://doi.org/10.3390/antibiotics12101554
Šakarnytė L, Šiugždinienė R, Žymantienė J, Ruzauskas M. Comparison of Oral Microbial Composition and Determinants Encoding Antimicrobial Resistance in Dogs and Their Owners. Antibiotics. 2023; 12(10):1554. https://doi.org/10.3390/antibiotics12101554
Chicago/Turabian StyleŠakarnytė, Laura, Rita Šiugždinienė, Judita Žymantienė, and Modestas Ruzauskas. 2023. "Comparison of Oral Microbial Composition and Determinants Encoding Antimicrobial Resistance in Dogs and Their Owners" Antibiotics 12, no. 10: 1554. https://doi.org/10.3390/antibiotics12101554
APA StyleŠakarnytė, L., Šiugždinienė, R., Žymantienė, J., & Ruzauskas, M. (2023). Comparison of Oral Microbial Composition and Determinants Encoding Antimicrobial Resistance in Dogs and Their Owners. Antibiotics, 12(10), 1554. https://doi.org/10.3390/antibiotics12101554