
Inhibition of Acinetobacter baumannii Biofilm Formation by Terpenes from Oregano (Lippia graveolens) Essential Oil

 ,
, 
 ,
,  and
and 
Abstract
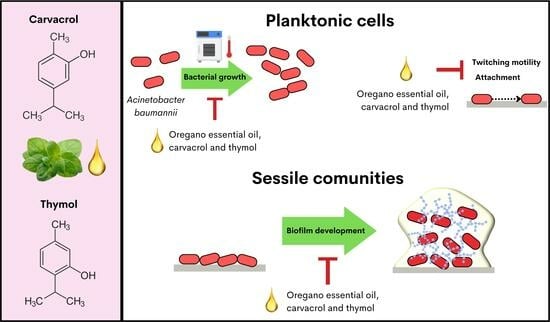
:
1. Introduction
2. Results and Discussion
2.1. Minimum Inhibitory and Bactericidal Concentration
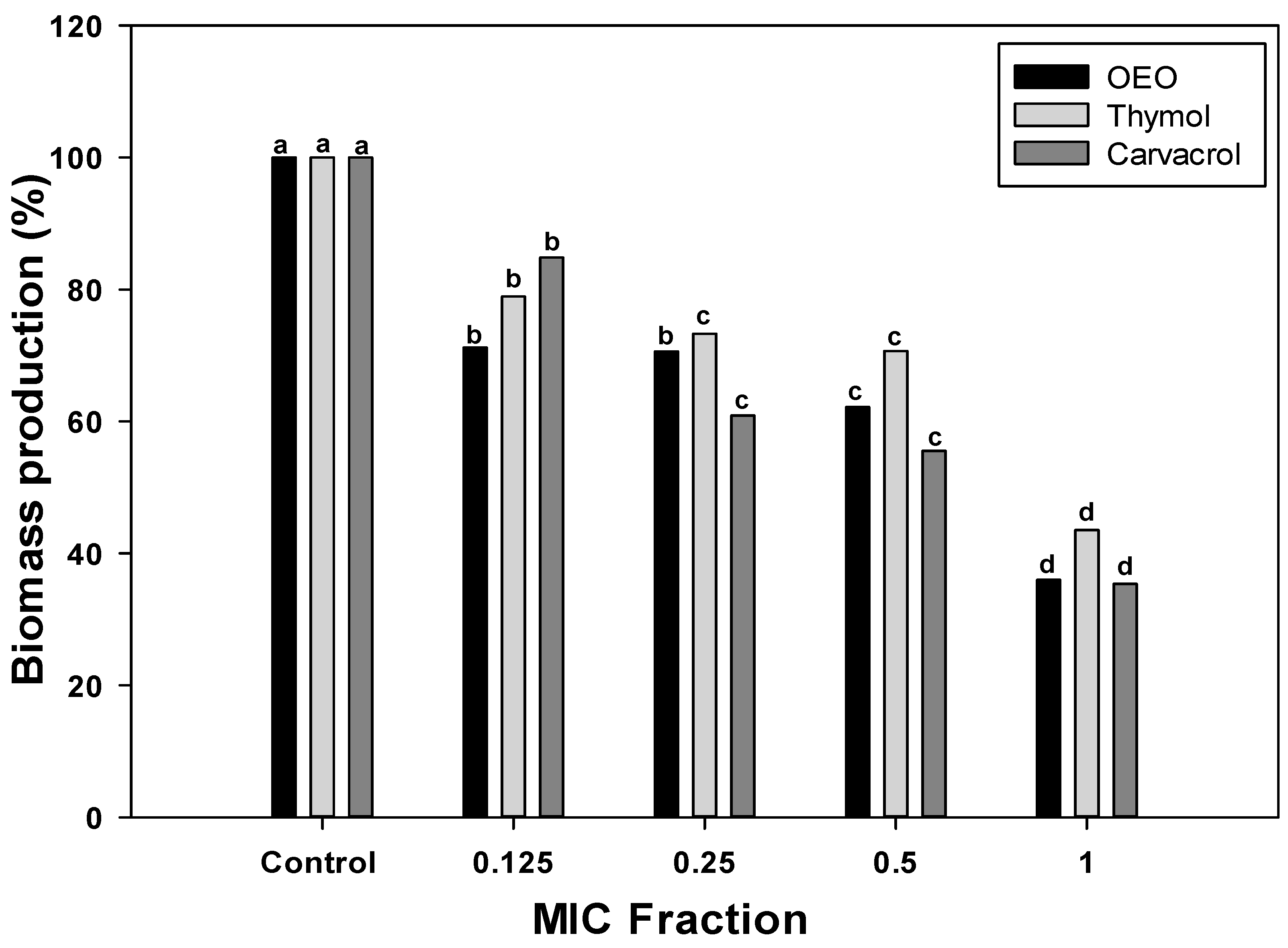
2.2. Changes in Biomass Production in A. baumannii Biofilms
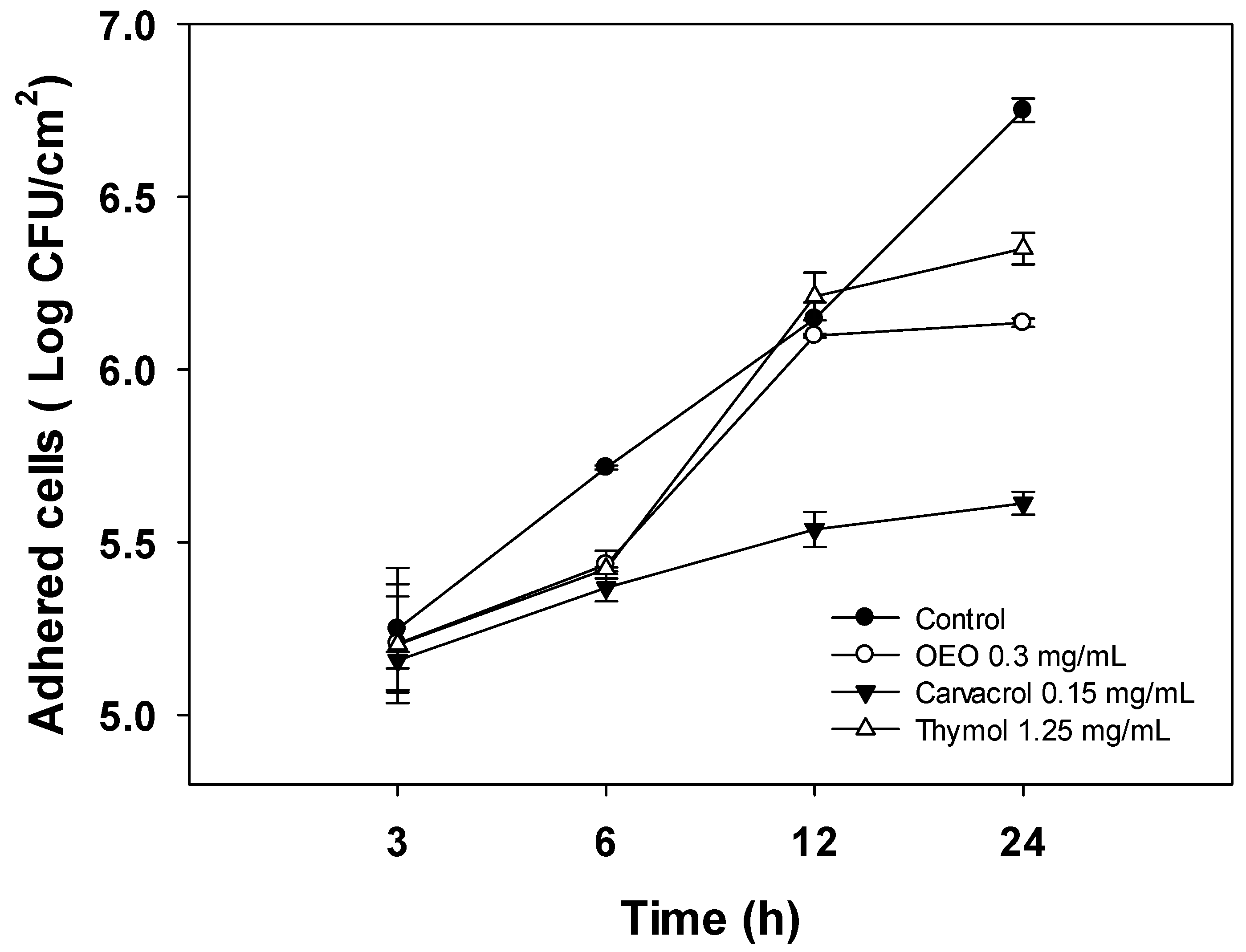
2.3. Changes in Cell Density of A. baumannii Biofilms Formed in the Presence of OEO, Carvacrol, and Thymol
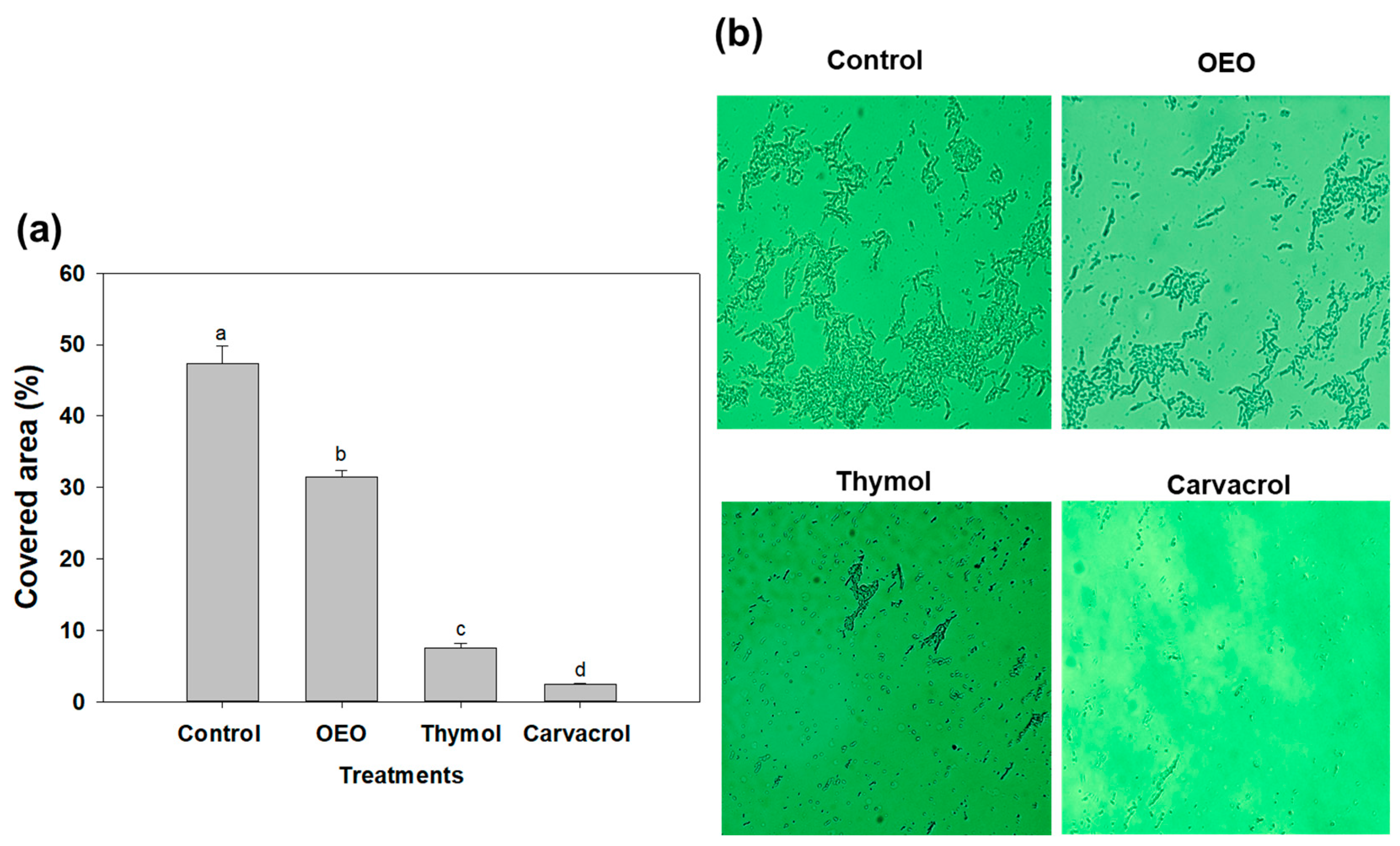
2.4. Fluorescence Microscopy of A. baumannii Biofilms Exposed to OEO, Carvacrol and Thymol
2.5. Inhibition of Twitching Motility
3. Materials and Methods
3.1. Bacteria Strains and Growth Conditions
3.2. Chemical Analysis of the Essential Oil
3.3. Minimal Inhibitory (MIC) and Bactericidal Concentrations (MBC) of OEO, Carvacrol and Thymol against A. baumannii
3.4. Biomass Production of A. baumannii Biofilms Exposed to OEO, Carvacrol, and Thymol
3.5. Effect of OEO, Carvacrol, and Thymol on Cell Density of A. baumannii Biofilms during Its Development on Stainless Steel Surfaces
3.6. Fluorescence Microscopy of A. baumannii Biofilms Exposed to OEO, Carvacrol and Thymol
3.7. Effect of OEO, Carvacrol, and Thymol on A. baumannii Twitching Motility
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hassannejad, N.; Bahador, A.; Rudbari, N.H.; Modarressi, M.H.; Parivar, K. In vivo antibacterial activity of Zataria multiflora Boiss extract and its components, carvacrol, and thymol, against colistin-resistant Acinetobacter baumannii in a pneumonic BALB/c mouse model. J. Cell. Biochem. 2019, 120, 18640–18649. [Google Scholar] [CrossRef] [PubMed]
- Appaneal, H.J.; O’Neill, E.; Lopes, V.V.; LaPlante, K.L.; Caffrey, A.R. National trends in hospital, long-term care and outpatient Acinetobacter baumannii resistance rates. J. Med. Microbiol. 2021, 70, 001473. [Google Scholar] [CrossRef]
- Gedefie, A.; Demsis, W.; Ashagrie, M.; Kassa, Y.; Tesfaye, M.; Tilahun, M.; Bisetegn, H.; Sahle, Z. Acinetobacter baumannii biofilm formation and its role in disease pathogenesis: A review. Infect. Drug Resist. 2021, 14, 3711–3719. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Villoria, A.M.; Valverde-Garduno, V. Antibiotic-resistant Acinetobacter baumannii increasing success remains a challenge as a nosocomial pathogen. J. Pathog. 2016, 2016, 7318075. [Google Scholar]
- Vo, N.; Sidner, B.S.; Yu, Y.; Piepenbrink, K.H. Type IV pilus-mediated inhibition of Acinetobacter baumannii biofilm formation by phenothiazine compounds. Microbiol. Spectr. 2023, 11, e0102323. [Google Scholar] [CrossRef] [PubMed]
- Shahed-Al-Mahmud, M.; Roy, R.; Sugiokto, F.G.; Islam, M.N.; Lin, M.-D.; Lin, L.-C.; Lin, N.-T. Phage φAB6-borne depolymerase combats Acinetobacter baumannii biofilm formation and infection. Antibiotics 2021, 10, 279. [Google Scholar] [CrossRef] [PubMed]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef]
- Kaye, K.S.; Pogue, J.M. Infections caused by resistant gram-negative bacteria: Epidemiology and management. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2015, 35, 949–962. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.-J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.M.; Tracy, E.N.; Carruthers, M.D.; Rather, P.N.; Actis, L.A.; Munson, R.S., Jr. Acinetobacter baumannii strain M2 produces type IV pili which play a role in natural transformation and twitching motility but not surface-associated motility. MBio 2013, 4, e00360-13. [Google Scholar] [CrossRef] [PubMed]
- Govindan, R.; Chackaravarthi, G.; Ramachandran, G.; Chelliah, C.K.; Muthuchamy, M.; Quero, F.; Mothana, R.A.; Noman, O.M.; Siddiqui, N.A.; Li, W.-J. Effective removal of biofilm formation in Acinetobacter baumannii using chitosan nanoparticles loaded plant essential oils. J. King Saud Univ. Sci. 2022, 34, 101845. [Google Scholar] [CrossRef]
- Bouyahya, A.; Dakka, N.; Et-Touys, A.; Abrini, J.; Bakri, Y. Medicinal plant products targeting quorum sensing for combating bacterial infections. Asian Pac. J. Trop. Med. 2017, 10, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Hernández, I.; Aguilar, C.N.; Martínez-Ávila, G.C.; Torres-León, C.; Ilina, A.; Flores-Gallegos, A.C.; Kumar Verma, D.; Chávez-González, M.L. Mexican Oregano (Lippia graveolens Kunth) as source of bioactive compounds: A review. Molecules 2021, 26, 5156. [Google Scholar] [CrossRef]
- Kachur, K.; Suntres, Z. The antibacterial properties of phenolic isomers, carvacrol and thymol. Crit. Rev. Food Sci. Nutr. 2020, 60, 3042–3053. [Google Scholar] [CrossRef] [PubMed]
- Krisilia, V.; Deli, G.; Koutsaviti, A.; Tzakou, O. Thymbra L. and Satureja L. essential oils as rich sources of carvacrol, a food additive with health-promoting effects. Am. J. Essent. Oils Nat. Prod. 2021, 9, 12–23. [Google Scholar]
- Rathod, N.B.; Kulawik, P.; Ozogul, F.; Regenstein, J.M.; Ozogul, Y. Biological activity of plant-based carvacrol and thymol and their impact on human health and food quality. Trends Food Sci. Technol. 2021, 116, 733–748. [Google Scholar] [CrossRef]
- Mahizan, N.A.; Yang, S.-K.; Moo, C.-L.; Song, A.A.-L.; Chong, C.-M.; Chong, C.-W.; Abushelaibi, A.; Lim, S.-H.E.; Lai, K.-S. Terpene derivatives as a potential agent against antimicrobial resistance (AMR) pathogens. Molecules 2019, 24, 2631. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Rodriguez, M.R.; Hernandez-Mendoza, A.; Gonzalez-Aguilar, G.A.; Martinez-Tellez, M.A.; Martins, C.M.; Ayala-Zavala, J.F. Carvacrol as potential quorum sensing inhibitor of Pseudomonas aeruginosa and biofilm production on stainless steel surfaces. Food Control 2017, 75, 255–261. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; De Feo, V.J.P. Essential oils and antifungal activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Fimbres-García, J.O.; Flores-Sauceda, M.; Othon-Díaz, E.D.; García-Galaz, A.; Tapia-Rodríguez, M.R.; Silva-Espinoza, B.A.; Ayala-Zavala, J.F. Facing resistant bacteria with plant essential oils: Reviewing the oregano case. Antibiotics 2022, 11, 1777. [Google Scholar] [CrossRef]
- Choudhary, M.; Shrivastava, R.; Vashistt, J. Eugenol and geraniol impede Csu-pilus assembly and evades multidrug-resistant Acinetobacter baumannii biofilms: In-vitro and in-silico evidence. Biochem. Biophys. Res. Commun. 2022, 636, 10–17. [Google Scholar] [CrossRef]
- Alves, S.; Duarte, A.; Sousa, S.; Domingues, F.C. Study of the major essential oil compounds of Coriandrum sativum against Acinetobacter baumannii and the effect of linalool on adhesion, biofilms and quorum sensing. Biofouling 2016, 32, 155–165. [Google Scholar] [CrossRef]
- Mirghani, R.; Saba, T.; Khaliq, H.; Mitchell, J.; Do, L.; Chambi, L.; Diaz, K.; Kennedy, T.; Alkassab, K.; Huynh, T. Biofilms: Formation, drug resistance and alternatives to conventional approaches. AIMS Microbiol. 2022, 8, 239. [Google Scholar] [CrossRef] [PubMed]
- Milani, E.S.; Hasani, A.; Varschochi, M.; Sadeghi, J.; Memar, M.Y. Biocide resistance in Acinetobacter baumannii: Appraising the mechanisms. J. Hosp. Infect. 2021, 117, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Sivaranjani, M.; Srinivasan, R.; Aravindraja, C.; Karutha Pandian, S.; Veera Ravi, A. Inhibitory effect of α-mangostin on Acinetobacter baumannii biofilms—An in vitro study. Biofouling 2018, 34, 579–593. [Google Scholar] [CrossRef]
- Nabawy, A.; Makabenta, J.M.; Li, C.-H.; Park, J.; Chattopadhyay, A.N.; Schmidt-Malan, S.; Gupta, A.; Patel, R.; Rotello, V.M. Activity of biodegradable polymeric nanosponges against dual-species bacterial biofilms. ACS Biomater. Sci. Eng. 2020, 7, 1780–1786. [Google Scholar] [CrossRef]
- Duménil, G. Type IV pili as a therapeutic target. Trends Microbiol. 2019, 27, 658–661. [Google Scholar] [CrossRef] [PubMed]
- Bukvički, D.; Cirić, A.; Soković, M.; Vannini, L.; Nissen, L.; Novaković, M.; Vujisić, L.; Asakawa, Y.; Marin, P.D. Micromeria thymifolia essential oil suppresses quorum-sensing signaling in Pseudomonas aeruginosa. Nat. Prod. Commun. 2016, 11, 1903–1906. [Google Scholar] [CrossRef] [PubMed]
- Chadha, J.; Harjai, K.; Chhibber, S. Repurposing phytochemicals as anti-virulent agents to attenuate quorum sensing-regulated virulence factors and biofilm formation in Pseudomonas aeruginosa. Microb. Biotechnol. 2022, 15, 1695–1718. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, I.; Cruz-Valenzuela, M.R.; Silva-Espinoza, B.A.; Gonzalez-Aguilar, G.A.; Moctezuma, E.; Gutierrez-Pacheco, M.M.; Tapia-Rodriguez, M.R.; Ortega-Ramirez, L.A.; Ayala-Zavala, J.F. Oregano (Lippia graveolens) essential oil added within pectin edible coatings prevents fungal decay and increases the antioxidant capacity of treated tomatoes. J. Sci. Food Agric. 2016, 96, 3772–3778. [Google Scholar] [CrossRef]
- Rajapaksha, D.C.; Edirisinghe, S.L.; Nikapitiya, C.; Whang, I.; De Zoysa, M. The Antimicrobial Peptide octopromycin suppresses biofilm formation and quorum sensing in Acinetobacter baumannii. Antibiotics 2023, 12, 623. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. baumannii | MIC (mg/mL) | MBC (mg/mL) |
|---|---|---|
| OEO | 0.6 | 1.2 |
| Carvacrol | 0.3 | 0.6 |
| Thymol | 2.5 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tapia-Rodriguez, M.R.; Cantu-Soto, E.U.; Vazquez-Armenta, F.J.; Bernal-Mercado, A.T.; Ayala-Zavala, J.F. Inhibition of Acinetobacter baumannii Biofilm Formation by Terpenes from Oregano (Lippia graveolens) Essential Oil. Antibiotics 2023, 12, 1539. https://doi.org/10.3390/antibiotics12101539
Tapia-Rodriguez MR, Cantu-Soto EU, Vazquez-Armenta FJ, Bernal-Mercado AT, Ayala-Zavala JF. Inhibition of Acinetobacter baumannii Biofilm Formation by Terpenes from Oregano (Lippia graveolens) Essential Oil. Antibiotics. 2023; 12(10):1539. https://doi.org/10.3390/antibiotics12101539
Chicago/Turabian StyleTapia-Rodriguez, Melvin Roberto, Ernesto Uriel Cantu-Soto, Francisco Javier Vazquez-Armenta, Ariadna Thalia Bernal-Mercado, and Jesus Fernando Ayala-Zavala. 2023. "Inhibition of Acinetobacter baumannii Biofilm Formation by Terpenes from Oregano (Lippia graveolens) Essential Oil" Antibiotics 12, no. 10: 1539. https://doi.org/10.3390/antibiotics12101539
APA StyleTapia-Rodriguez, M. R., Cantu-Soto, E. U., Vazquez-Armenta, F. J., Bernal-Mercado, A. T., & Ayala-Zavala, J. F. (2023). Inhibition of Acinetobacter baumannii Biofilm Formation by Terpenes from Oregano (Lippia graveolens) Essential Oil. Antibiotics, 12(10), 1539. https://doi.org/10.3390/antibiotics12101539