
Multidrug-Resistant ESBL-Producing E. coli in Clinical Samples from the UK

 , and
, and
Abstract
1. Introduction
2. Results
2.1. Strains
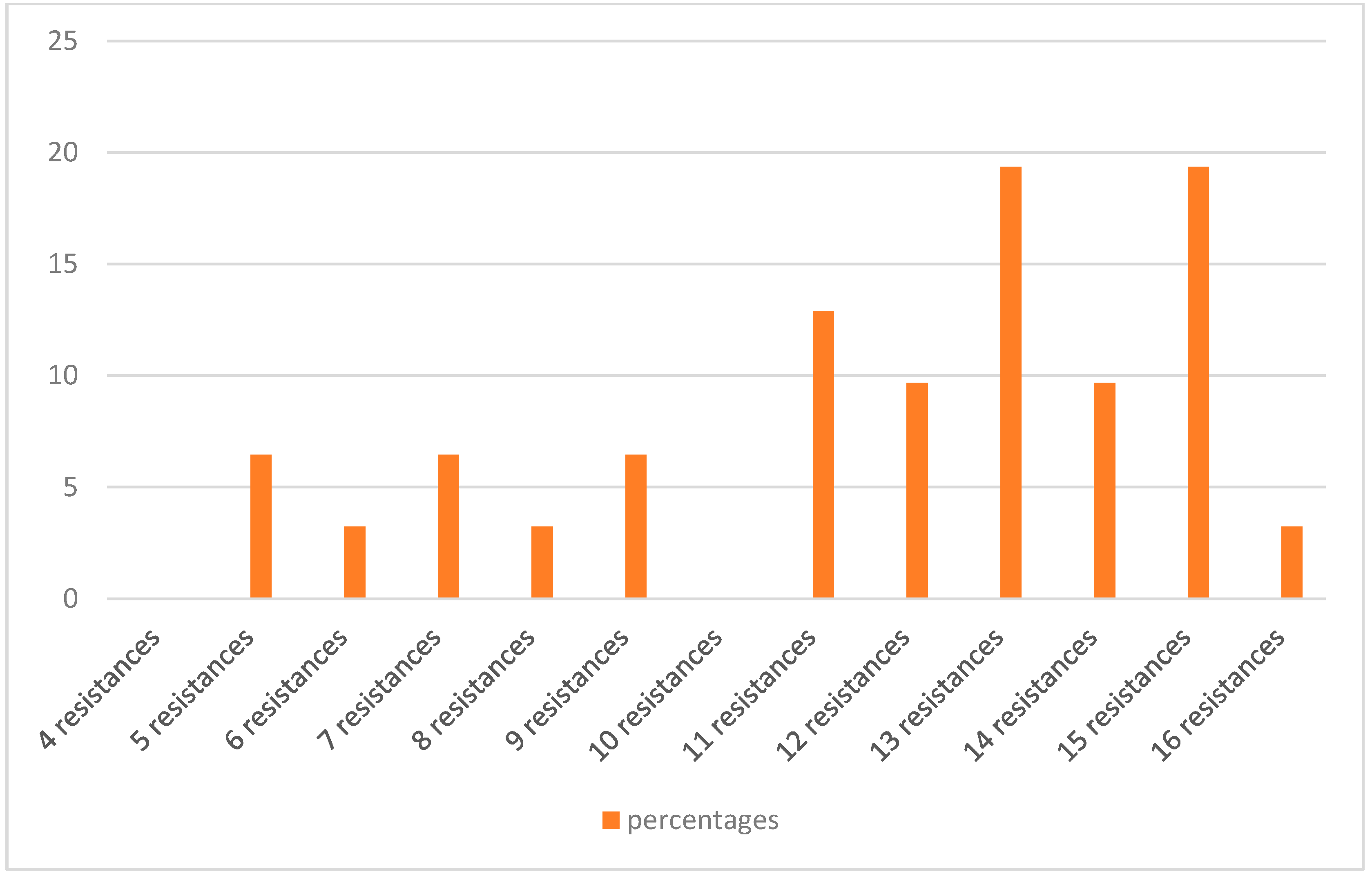
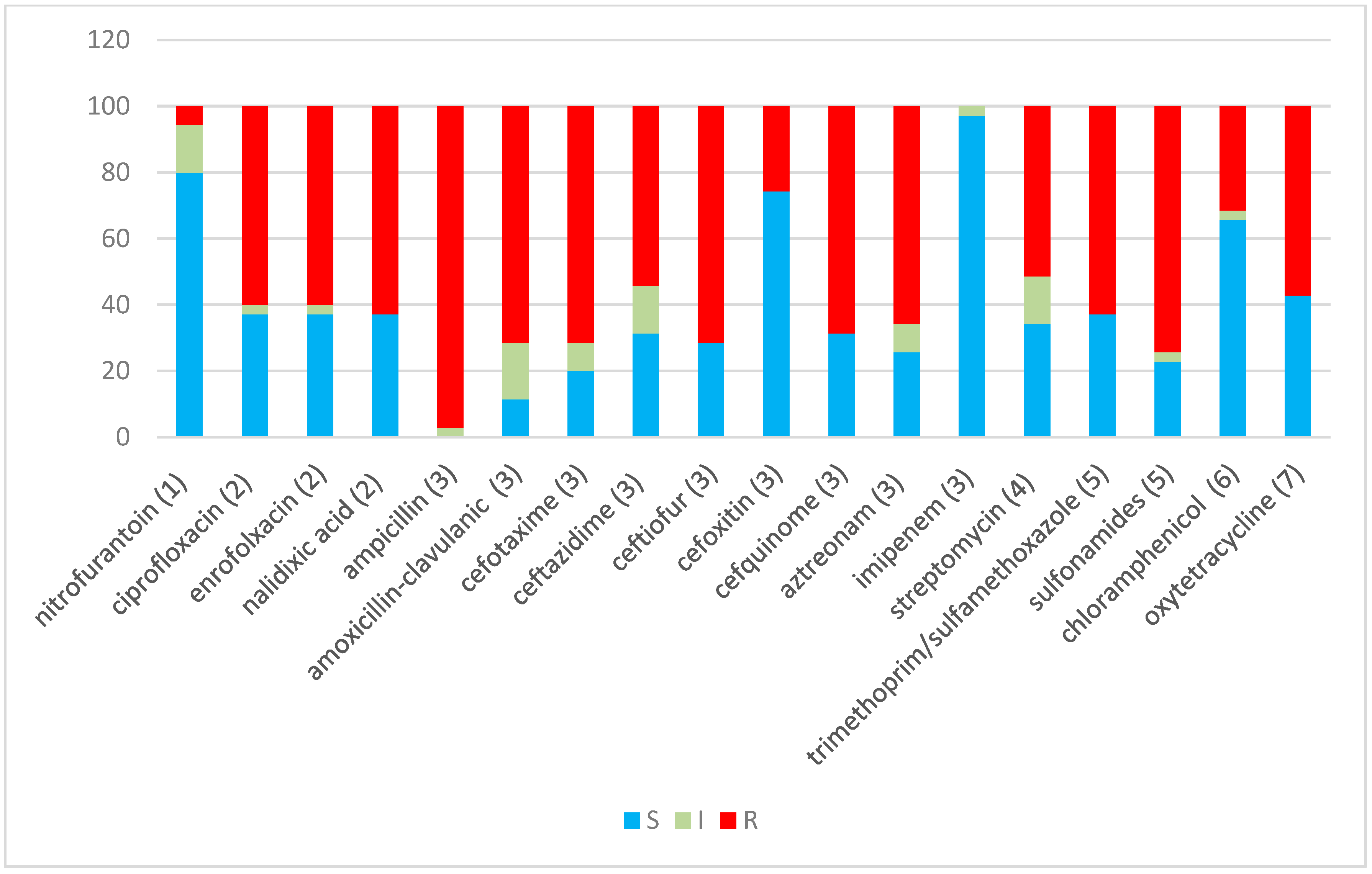
2.2. Antibiotic Resistance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate ID | β-Lactamases | Gene Sequence Type | Intl1/merA and merC | ESBL Screening Test | AmpC Screening Test | Antimicrobial Resistance Profile |
|---|---|---|---|---|---|---|
| H1B | CTX-M/OXA | CTX-M-55/OXA-1 | Intl1 | + * | * STX, AMC, OT, CAZ, NA, EFT, S300, CFQ, ATM, AMP, ENR, CTX, CIP | |
| H2B | CTX-M/OXA | CTX-M-285/OXA-1 | Intl1 | + | STX, AMC, CAZ, NA, EFT, S300, CFQ, ATM, AMP, ENR, CTX, CIP | |
| H3B | CTX-M/OXA | CTX-M-15/OXA-1 | + | AMC, OT, CAZ, NA, EFT, CFQ, ATM, AMP, (C), ENR, CTX, CIP | ||
| H4B | CTX-M/TEM | ND/TEM-1 | + | - | AMC, OT, S10, CAZ, NA, EFT, S300, CFQ, ATM, AMP, C, ENR, CTX, CIP, FOX | |
| H5B | CTX-M/TEM/ | CTX-M-15/ TEM-1 | + | AMC, OT, S10, CAZ, NA, EFT, S300, CFQ, ATM, AMP, C, ENR, CTX, CIP | ||
| H6B | TEM | − | - | AMC, (ATM), AMP, (CTX), FOX | ||
| H7B | CTX-M/OXA | ND/OXA-1 | Intl1 | + | STX, OT, (S10), (CAZ), NA, EFT, S300, CFQ, ATM, AMP, ENR, CTX, CIP | |
| H8B | CTX-M/OXA | CTX-M-254/OXA-1 | Intl1 | + | STX, (AMC), OT, CAZ, NA, EFT, S300, CFQ, ATM, AMP, ENR, CTX, CIP | |
| H9B | CTX-M/OXA | ND/OXA-1 | Intl1 | + | STX, AMC, OT, (S10), (CAZ), NA, EFT, S300, CFQ, ATM, AMP, ENR, CTX, CIP | |
| H10B | CTX-M/SHV/TEM/OXA/plasmid AmpC | ND/ND/ND/OXA-1/CIT/FOX | Intl1 | ND | ND | STX, AMC, OT, S10, EFT, S300, CFQ, ATM, AMP, CTX, (CIP) |
| H11B | CTX-M/TEM | ND/ND | + | - | AMC, OT, S10, CAZ, NA, EFT, S300, CFQ, ATM, AMP, C, ENR, CTX, CIP, FOX | |
| H12B | CTX-M | ND | + | - | AMC, (F), OT, S10, CAZ, NA, EFT, S300, CFQ, ATM, AMP, C, ENR, CTX, CIP, FOX | |
| H13B | CTX-M | ND | + | STX, S10, OT, CAZ, NA, EFT, S300, CFQ, ATM, AMP, C, ENR, CTX, CIP | ||
| H14B | CTX-M | ND | Intl1 | + | STX, (S10), CAZ, NA, EFT, S300, CFQ, ATM, AMP, C, ENR, CTX, CIP | |
| H15B | CTX-M/OXA | CTX-M-15/OXA-1 | Intl1 | + | STX, AMC, (S10), CAZ, NA, EFT, S300, CFQ, ATM, AMP, C, CTX, CIP | |
| H16B | CTX-M | ND | Intl1 | + | STX, AMC, CAZ, NA, EFT, S300, CFQ, ATM, AMP, ENR, CTX, CIP | |
| H17B | + | - | AMC, OT, S10, CAZ, NA, EFT, S300, CFQ, ATM, AMP, C, ENR, CTX, CIP, FOX | |||
| H18B | + | - | AMC, OT, S10, CAZ, NA, EFT, S300, CFQ, ATM, AMP, C, ENR, CTX, CIP, FOX | |||
| H19U | Intl1 | − | - | STX, AMC, (F), OT, S10, (CAZ), NA, S300, AMP, ENR, (CTX), CIP, FOX | ||
| H20U | SHV/TEM/plasmid AmpC | ND/TEM-1/CIT/FOX | Intl1/merA | ND | ND | SXT, AMC, F, OT, (IMP), S10, S300, AMP |
| H21U | CTX-M/TEM | ND/ND | merA/merC | + | STX, AMC, S10, (CAZ), EFT, S300, CFQ, (ATM), AMP, ENR, CTX | |
| H22U | CTX-M/TEM | CTX-M-14/TEM-1 | + | STX, (AMC), S10, EFT, S300, CFQ, (ATM), AMP, CTX | ||
| H23U | TEM | Intl1 | ND | STX, AMC, NA, S300, AMP, (ENR) | ||
| H24U | TEM | TEM-30 | ND | STX, AMC, S10, AMP | ||
| H25R | TEM | TEM-32 | Intl1 | ND | AMC, F, OT, S10, AMP | |
| H26B | CTX-M/TEM | CTX-M-15/TEM-1 | Intl1/merA/ merC | + | - | STX, AMC, S10, CAZ, EFT, S300, CFQ, ATM, AMP, C, CTX |
| H27B | CTX-M | CTX-M-55 | + | AMC, CAZ, EFT, ATM, AMP, CTX | ||
| H28E | CTX-M/OXA | ND/OXA-1 | Intl1 | + | - | STX, AMC, OT, CAZ, NA, EFT, S300, CFQ, ATM, AMP, C, ENR, CTX, CIP, FOX |
| H29F | − | (AMP) | ||||
| H30R | CTX-M/OXA | ND/OXA-1 | + | AMC, OT, CAZ, NA, EFT, CFQ, ATM, AMP, ENR, CTX, CIP | ||
| H31B | CTX-M/OXA | ND/OXA-1 | Intl1 | + | STX, AMC, (S10), CAZ, NA, EFT, S300, CFQ, ATM, AMP, ENR, CTX, CIP | |
| H32F | CTX-M/OXA | CTX-M-254/OXA-1 | Intl1 | + | STX, AMC, (F), OT, S10, CAZ, NA, EFT, S300, CFQ, ATM, AMP, ENR, CTX, CIP | |
| H33F | TEM | TEM-1 | − | + | AMC, AMP, (CTX), FOX | |
| H34U | ND | ND | STX, AMC, (F), OT, S10, S300, AMP | |||
| H35U | ND | ND | STX, AMC, (F), OT, S10, S300, AMP |
| Antibiotics/Abbreviation | Antimicrobial Class | Disc Content |
|---|---|---|
| Ampicillin (AMP) | Β-Lactam/Penicillin | 10 μg |
| Amoxicillin–clavulanic acid (AMC) | Β-Lactam/Penicillin Combination with Beta-Lactamase Inhibitor | 20/10 μg |
| Aztreonam (ATM) | Β-Lactam/Monobactam | 30 μg |
| Cefotaxime (CTX) | Β-Lactam/Third-Generation Cephalosporin | 30 μg |
| Ceftazidime (CAZ) | Β-Lactam/Third-Generation Cephalosporin | 30 μg |
| Cefquinome (CFQ) | β-Lactam/Fourth-Generation Cephalosporin | 30 μg |
| Ceftiofur (EFT) | β-Lactam/Third-Generation Cephalosporin | 30 μg |
| Cefoxitin (FOX) | β-Lactam/Second-Generation Cephalosporin | 30 μg |
| Imipenem (IMP) | β-Lactam/Carbapenem | 10 μg |
| Streptomycin (S10) | Aminoglycoside | 10μg |
| Ciprofloxacin (CIP) | Quinolone/Fluoroquinolone | 5 μg |
| Enrofloxacin (ENR) | Quinolone/Fluoroquinolone | 5 μg |
| Nalidixic acid (NA) | Quinolone | 30 μg |
| Nitrofurantoin (F) | Nitrofuran derivative | 300 μg |
| Chloramphenicol (C) | Phenicol | 30 μg |
| Trimethoprim–sulfamethoxazole (SXT) | Sulphonamide/complex | 1.25/23.75 μg |
| Sulphonamide (S300) | Sulphonamide | 300 μg |
| Oxytetracycline (OT) | Tetracycline | 30 μg |
2.3. Phenotyping of ESBL/AmpC and PCR Typing of ESBL Genes
2.4. Intl1, merA and merC Detection
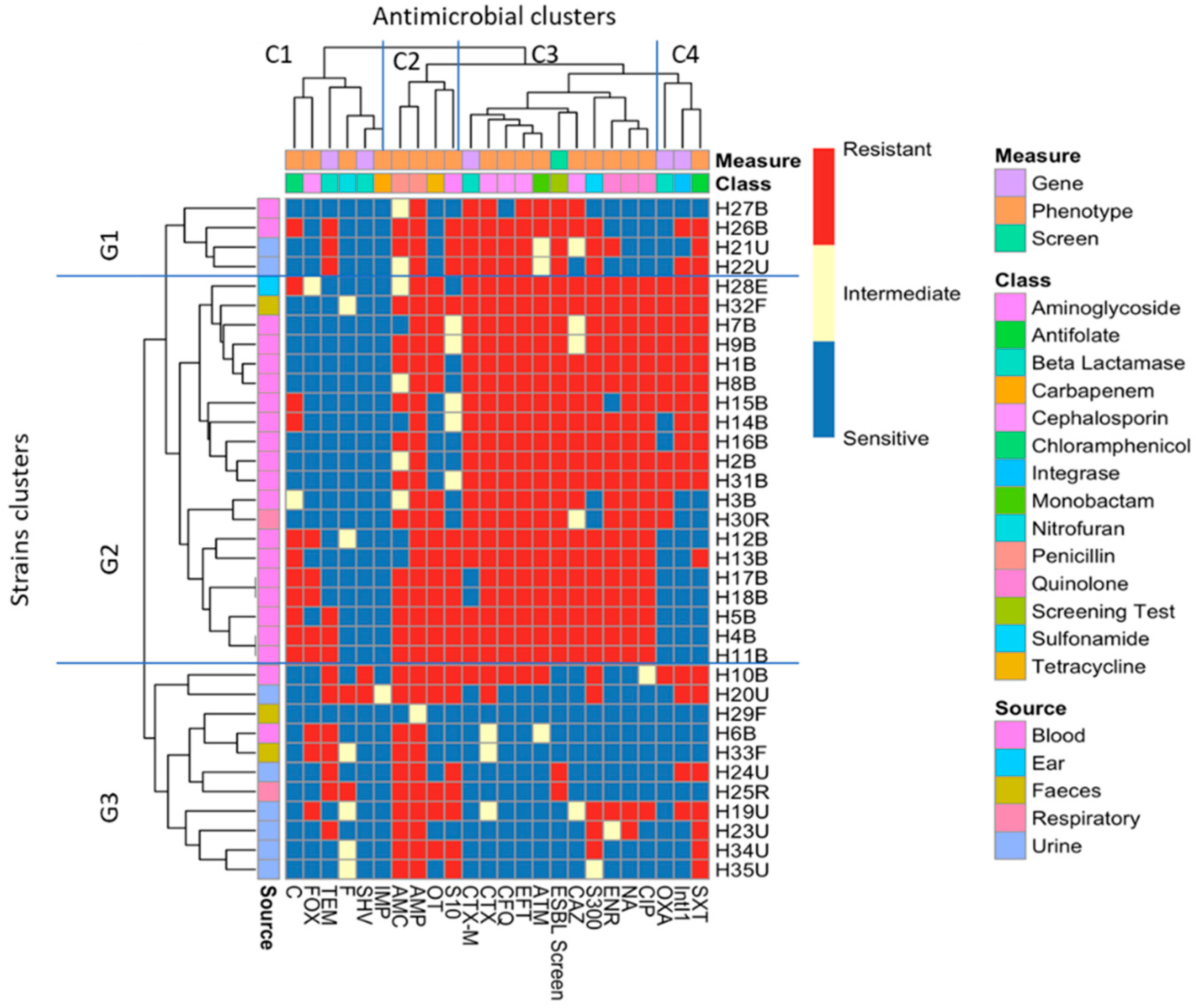
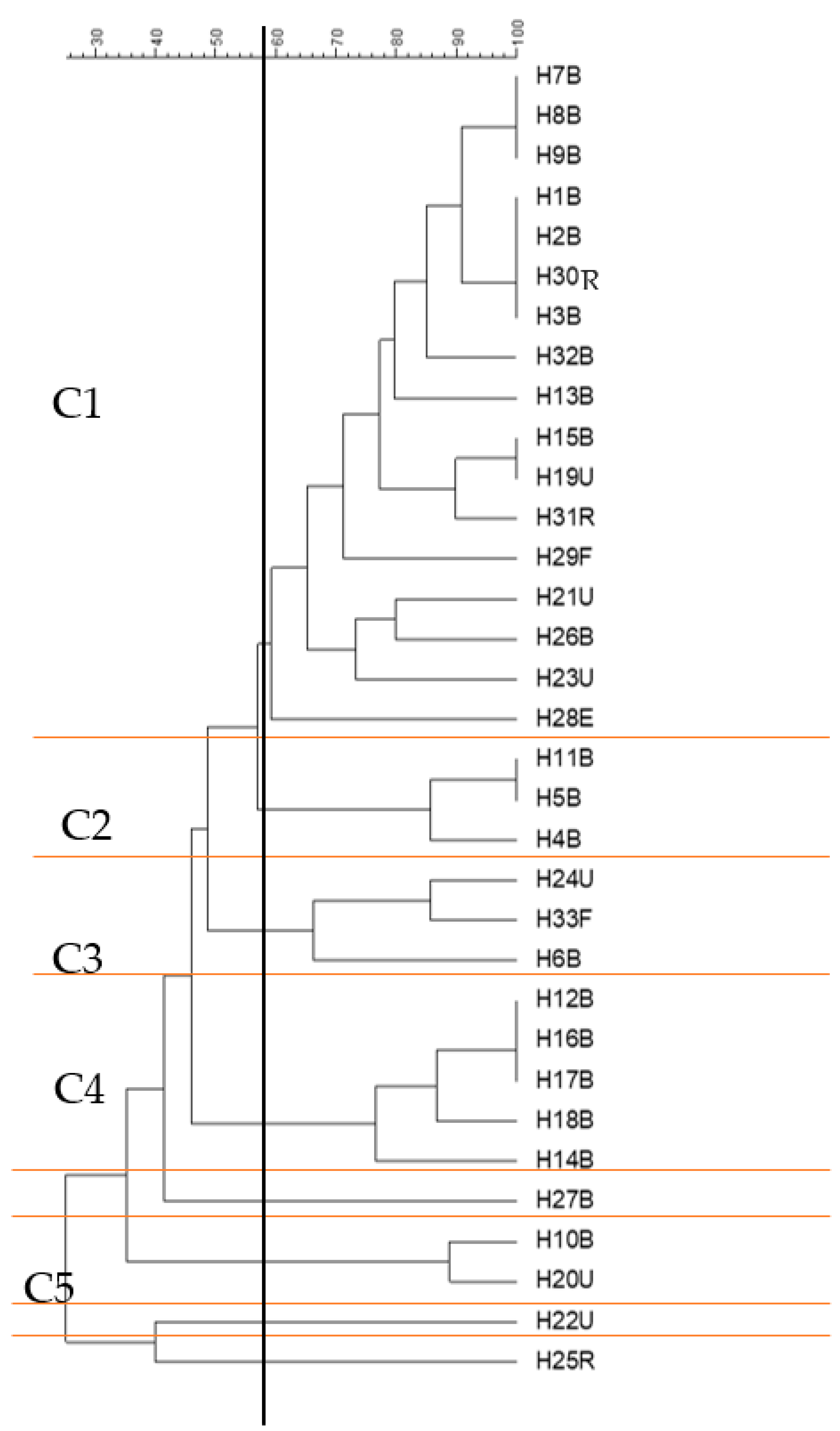
2.5. Cluster Analysis of the Antibiotic Sensitivity Results with Different Genes and Screening Tests
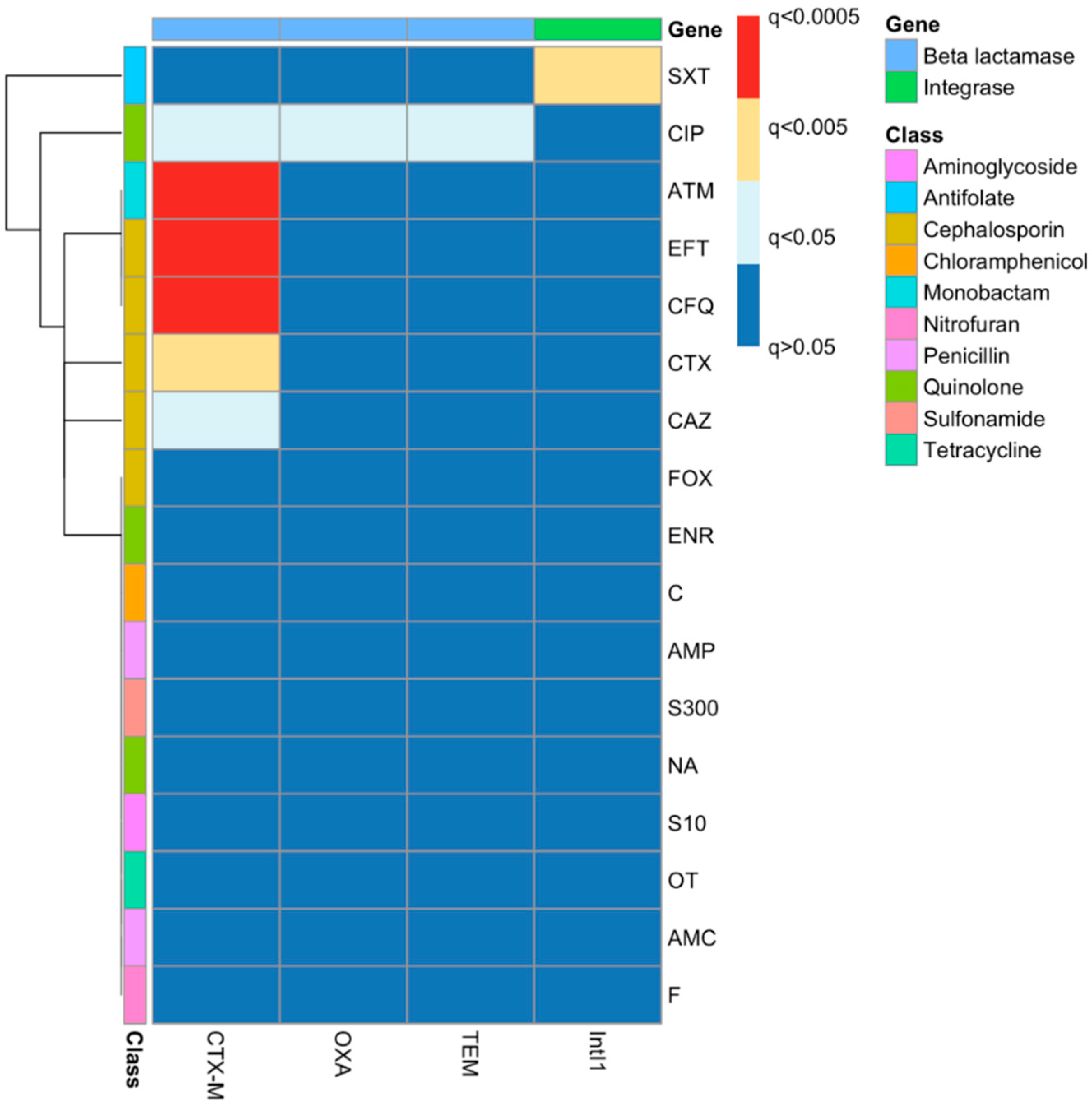
2.6. Phenotypic–Genotypic Association Using Chi-Squared Tests
2.7. Genotyping of E. coli Strains Using ERIC-PCR
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Antibiotic Sensitivity Tests
4.3. Phenotypic Confirmation of ESBL/AmpC-Producing E. coli
4.4. Genotyping of Isolates
4.4.1. DNA Extraction
4.4.2. ERIC-PCR
4.5. PCR Detection of Β-lactamase Genes
4.6. PCR Detection of Plasmid ampC
| Oligonucleotide Name | Sequence | Product Size (bp) | Reference |
|---|---|---|---|
| CTX-M-F | ATGTGCAGYACCAGTAARGTKATGGC | 529 a | [73] |
| CTX-M-R | TGGGTRAARTARGTSACCAGAAYSAGCGG | ||
| TEM-F | GCGGAACCCCTATTTG | 964 | |
| TEM-R | ACCAATGCTTAATCAGTGAG | ||
| SHV-F- | TTATCTCCCTGTTAGCCACC | 796 | |
| SHV-R- | GATTTGCTGATTTCGCTCGG | ||
| OXA-1-F- | ATGAAAAACACAATACATATCAACTTCGC | 820 | |
| OXA-1-R- | GTGTGTTTAGAATGGTGATCGCATT | ||
| OXA-2-F- | ACGATAGTTGTGGCAGACGAAC | 601 | |
| OXA-2-R- | ATYCTGTTTGGCGTATCRATATTC | ||
| CMY-2-F- | ATGATGAAAAAATCGTTATGCTGC | 1138 | |
| CMY-2-R | GCTTTTCAAGAATGCGCCAGG | ||
| MultiACC-F- | CACCTCCAGCGACTTGTTAC | 346 | [76] |
| MultiACC-R- | GTTAGCCAGCATCACGATCC | ||
| MultiMOX-F- | GCAACAACGACAATCCATCCT | 895 | |
| MultiMOX-R- | GGGATAGGCGTAACTCTCCCAA | ||
| MultiDHA-F- | TGATGGCACAGCAGGATATTC | 997 | |
| MultiDHA-R- | GCTTTGACTCTTTCGGTATTCG | ||
| MultiCIT-F- | CGAAGAGGCAATGACCAGAC | 538 | |
| MultiCIT-R- | ACGGACAGGGTTAGGATAGY | ||
| MultiEBC-F- | CGGTAAAGCCGATGTTGCG | 683 | |
| MultiEBC-R- | AGCCTAACCCCTGATACA | ||
| MultiFOX-F- | CTACAGTGCGGGTGGTTT | 162 | |
| MultiFOX-R- | CTATTTGCGGCCAGGTGA | ||
| ERIC-F- | ATGTAAGCTCCTGGGGATTCAC | variable | [70] |
| ERIC-R- | AAGTAAGTGACTGGGGTGAGCG |
4.7. 16S rDNA V3 Region PCR
4.8. Sequencing of PCR Products
4.9. Statistical Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Rev. Antimicrob. Resist. 2016. Available online: https://amr-review.org/sites/default/files/160518_Final%20paper_with%20cover.pdf (accessed on 1 September 2022).
- O’Neill, J. Infection Prevention, Control and Surveillance: Limiting the Development and Spread of Drug Resistance. Rev. Antimicrob. Resist. 2016. Available online: https://amr-review.org/sites/default/files/Health%20infrastructure%20and%20surveillance%20final%20version_LR_NO%20CROPS.pdf (accessed on 1 September 2022).
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- PHE. English Surveillance Programme for Antimicrobial Utilisation and Resistance (ESPAUR): Report 2020 to 2021; GOV.UK: 2021. Available online: https://www.gov.uk/government/publications/english-surveillance-programme-antimicrobial-utilisation-and-resistance-espaur-report (accessed on 1 September 2022).
- Bradford, P.A. Extended-Spectrum β-Lactamases in the 21st Century: Characterization, Epidemiology, and Detection of This Important Resistance Threat. Clin. Microbiol. Rev. 2001, 14, 933–951. [Google Scholar] [CrossRef]
- Bush, K.; Macielag, M.J. New β-Lactam Antibiotics and β-Lactamase Inhibitors. Expert Opin. Ther. Pat. 2010, 20, 1277–1293. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Simner, P.J.; Bradford, P.A. Extended-Spectrum β -Lactamases: An Update on Their Characteristics, Epidemiology and Detection. JAC-Antimicrob. Resist. 2021, 3, dlab092. [Google Scholar] [CrossRef] [PubMed]
- Nossair, M.A.; Alkhedaide, A.; Soliman, M.M.; Ramadan, H. Prevalence and Molecular Characterization of Cattle, and Poultry. Pathogens 2022, 2022.11, 852. [Google Scholar] [CrossRef] [PubMed]
- Miltgen, G.; Martak, D.; Valot, B.; Kamus, L.; Garrigos, T.; Verchere, G.; Gbaguidi-Haore, H.; Ben Cimon, C.; Ramiandrisoa, M.; Picot, S.; et al. One Health Compartmental Analysis of ESBL-Producing Escherichia coli on Reunion Island Reveals Partitioning between Humans and Livestock. J. Antimicrob. Chemother. 2022, 77, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Subramanya, S.H.; Bairy, I.; Metok, Y.; Baral, B.P.; Gautam, D.; Nayak, N. Detection and Characterization of ESBL-Producing Enterobacteriaceae from the Gut of Subsistence Farmers, Their Livestock, and the Surrounding Environment in Rural Nepal. Sci. Rep. 2021, 11, 2091. [Google Scholar] [CrossRef]
- Benavides, J.A.; Salgado-Caxito, M.; Opazo-Capurro, A.; González Muñoz, P.; Piñeiro, A.; Otto Medina, M.; Rivas, L.; Munita, J.; Millán, J. ESBL-Producing Escherichia coli Carrying CTX-M Genes Circulating among Livestock, Dogs, and Wild Mammals in Small-Scale Farms of Central Chile. Antibiotics 2021, 10, 510. [Google Scholar] [CrossRef]
- Day, M.J.; Hopkins, K.L.; Wareham, D.W.; Toleman, M.A.; Elviss, N.; Randall, L.; Teale, C.; Cleary, P.; Wiuff, C.; Doumith, M.; et al. Extended-Spectrum β-Lactamase-Producing Escherichia coli in Human-Derived and Foodchain-Derived Samples from England, Wales, and Scotland: An Epidemiological Surveillance and Typing Study. Lancet Infect. Dis. 2019, 19, 1325–1335. [Google Scholar] [CrossRef]
- UKHSA. Escherichia coli (E. coli) Bacteraemia: Financial Year Counts and Rates by Acute Trust and CCG, up to Financial Year 2021 to 2022 (Open Source). 2021. Available online: https://www.gov.uk/government/statistics/escherichia-coli-e-coli-bacteraemia-annual-data (accessed on 10 August 2021).
- Torres, A.G.; Arenas-Hernández, M.M.; Martínez-Laguna, Y. Overview of E. coli. In Pathogenic Escherichia coli in Latin America; Bentham Science Publishers: Sharjah, United Arab Emirates, 2010; pp. 1–7. [Google Scholar]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Finlay, B.B. Molecular Mechanisms of Escherichia coli Pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef]
- Riley, L.W. Distinguishing Pathovars from Nonpathovars: Escherichia coli. Microbiol. Spectr. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Kayser, F.H.; Bienz, K.A.; Eckert, J.; Zinkernagel, R.M. Medical Microbiology; Thieme: New York, NY, USA, 2005. [Google Scholar]
- Reinthaler, F.F.; Posch, J.; Feierl, G.; Wüst, G.; Haas, D.; Ruckenbauer, G.; Mascher, F.; Marth, E. Antibiotic Resistance of E. coli in Sewage and Sludge. Water Res. 2003, 37, 1685–1690. [Google Scholar] [CrossRef]
- Pitout, J.D.D.; Sanders, C.C.; Sanders, W.E., Jr. Antimicrobial Resistance with Focus on β-Lactam Resistance in Gram-Negative Bacilli. Am. J. Med. 1997, 103, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Segreti, J. Overview of the Epidemiological Profile and Laboratory Detection of Extended-Spectrum β-Lactamases. Clin. Infect. Dis. 2006, 42 (Suppl. S4), 153–163. [Google Scholar] [CrossRef] [PubMed]
- Wellington, E.M.H.; Boxall, A.B.A.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The Role of the Natural Environment in the Emergence of Antibiotic Resistance in Gram-Negative Bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef]
- Ibrahim, D.R. Phenotypic and Genotypic Study of Multidrug Resistant, Extended Spectrum β-Lactamase (ESBL)-Producing Escherichia coli Isolated from a Dairy Farm. Ph.D. Dissertation, University of Nottingham, Nottingham, UK, 2017. Available online: https://eprints.nottingham.ac.uk/id/eprint/46613 (accessed on 1 November 2022).
- Sultan, I.; Siddiqui, M.T.; Gogry, F.A.; Haq, Q.M.R. Molecular Characterization of Resistance Determinants and Mobile Genetic Elements of ESBL Producing Multidrug-Resistant Bacteria from Freshwater Lakes in Kashmir, India. Sci. Total Environ. 2022, 827, 154221. [Google Scholar] [CrossRef]
- Qiao, J.; Zhang, Q.; Alali, W.Q.; Wang, J.; Meng, L.; Xiao, Y.; Yang, H.; Chen, S.; Cui, S.; Yang, B. Characterization of Extended-Spectrum β-Lactamases (ESBLs)-Producing Salmonella in Retail Raw Chicken Carcasses. Int. J. Food Microbiol. 2017, 248, 72–81. [Google Scholar] [CrossRef]
- Liebert, C.A.; Hall, R.M.; Summers, A.O. Transposon Tn 21, Flagship of the Floating Genome. Microbiol. Mol. Biol. Rev. 1999, 63, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Liebert, C.A.; Wireman, J.; Smith, T.; Summers, A.O. Phylogeny of mercury Resistance (Mer) Operons of Gram-Negative Bacteria Isolated from the Fecal Flora of Primates. Appl. Environ. Microbiol. 1997, 63, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R. Analysis of Antibiotic Resistance Regions in Gram-Negative Bacteria. FEMS Microbiol. Rev. 2011, 35, 820–855. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated Functional Classification of β-Lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.G.; Barlow, M. Revised Ambler Classification of β-Lactamases. J. Antimicrob. Chemother. 2005, 55, 1050–1051. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L.; Bonomo, R.A. Extended-Spectrum β-Lactamases: A Clinical Update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [PubMed]
- CLSI. (M31-A4) Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Clinical and Laboratory Standards Institute: Malvern, PA, USA, 2013. [Google Scholar]
- Philippon, A.; Arlet, G.; Jacoby, G.A. Plasmid-Determined AmpC-Type-β-Lactamases. Antimicrob. Agents Chemother. 2002, 46, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Livermore, D.M. Defining an Extended-Spectrum b -Lactamase. Clin. Microbiol. Infect. 2008, 14, 3–10. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Brenwald, N.P.; Jevons, G.; Andrews, J.; Ang, L.; Fraise, A.P. Disc Methods for Detecting AmpC β-Lactamase-Producing Clinical Isolates of Escherichia coli and Klebsiella pneumoniae. J. Antimicrob. Chemother. 2005, 56, 600–601. [Google Scholar] [CrossRef]
- WHO. Global Antimicrobial Resistance Surveillance System (GLASS) Report: Early Implementation 2020; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- Tadesse, D.A.; Zhao, S.; Tong, E.; Ayers, S.; Singh, A.; Bartholomew, M.J.; McDermott, P.F. Antimicrobial Drug Resistance in Escherichia coli from Humans and Food Animals, United States, 1950–2002. Emerg. Infect. Dis. 2012, 18, 741. [Google Scholar] [CrossRef] [PubMed]
- Pandit, R.; Awal, B.; Shrestha, S.S.; Joshi, G.; Rijal, B.P.; Parajuli, N.P. Extended-Spectrum β-Lactamase (ESBL) Genotypes among Multidrug-Resistant Uropathogenic Escherichia coli Clinical Isolates from a Teaching Hospital of Nepal. Interdiscip. Perspect. Infect. Dis. 2020, 2020, 6525826. [Google Scholar] [CrossRef]
- Wu, D.; Ding, Y.; Yao, K.; Gao, W.; Wang, Y. Antimicrobial Resistance Analysis of Clinical Escherichia coli Isolates in Neonatal Ward. Front. Pediatr. 2021, 9, 670470. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.-H. Escherichia coli Contamination of Pork Carcasses in UK Slaughterhouses. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 2013. Available online: https://eprints.nottingham.ac.uk/id/eprint/13477 (accessed on 23 September 2021).
- Ibrahim, D.R.; Dodd, C.E.R.; Stekel, D.J.; Ramsden, S.J.; Hobman, J.L. Multidrug Resistant, Extended Spectrum β-Lactamase (ESBL)-Producing Escherichia coli Isolated from a Dairy Farm. FEMS Microbiol. Ecol. 2016, 92, fiw013. [Google Scholar] [CrossRef] [PubMed]
- Papich, M.G. Saunders Handbook of Veterinary Drugs; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Sprayberry, K.A.; Robinson, N.E. Robinson’s Current Therapy in Equine Medicine; Elsevier Health Sciences: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Hornish, R.E.; Kotarski, S.F. Cephalosporins in Veterinary Medicine-Ceftiofur Use in Food Animals. Curr. Top. Med. Chem. 2002, 2, 717–731. [Google Scholar] [CrossRef]
- Grabowski, Ł.; Gaffke, L.; Pierzynowska, K.; Cyske, Z.; Choszcz, M.; Węgrzyn, G.; Węgrzyn, A. Enrofloxacin-The Ruthless Killer of Eukaryotic Cells or the Last Hope in the Fight against Bacterial Infections? Int. J. Mol. Sci. 2022, 23, 3648. [Google Scholar] [CrossRef]
- Estaleva, C.E.L.; Zimba, T.F.; Sekyere, J.O.; Govinden, U.; Chenia, H.Y.; Simonsen, G.S.; Haldorsen, B.; Essack, S.Y.; Sundsfjord, A. High Prevalence of Multidrug Resistant ESBL- and Plasmid Mediated AmpC-Producing Clinical Isolates of Escherichia coli at Maputo Central Hospital, Mozambique. BMC Infect. Dis. 2021, 21, 16. [Google Scholar] [CrossRef] [PubMed]
- Cartelle, M.; del Mar Tomas, M.; Molina, F.; Moure, R.; Villanueva, R.; Bou, G. High-Level Resistance to Ceftazidime Conferred by a Novel Enzyme, CTX-M-32, Derived from CTX-M-1 through a Single Asp240-Gly Substitution. Antimicrob. Agents Chemother. 2004, 48, 2308–2313. [Google Scholar] [CrossRef]
- Marshall, B.M.; Levy, S.B. Food Animals and Antimicrobials: Impacts on Human Health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Ahrens, P.; Madsen, M.; Pallesen, L.V.; Poulsen, R.L.; Westh, H. Glycopeptide Susceptibility among Danish Enterococcus Faecium and Enterococcus Faecalis Isolates of Animal and Human Origin and PCR Identification of Genes within the VanA Cluster. Antimicrob. Agents Chemother. 1996, 40, 1938–1940. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Seyfarth, A.M.; Emborg, H.-D.; Pedersen, K.; Hendriksen, R.S.; Bager, F. Effect of Abolishment of the Use of Antimicrobial Agents for Growth Promotion on Occurrence of Antimicrobial Resistance in Fecal Enterococci from Food Animals in Denmark. Antimicrob. Agents Chemother. 2001, 45, 2054–2059. [Google Scholar] [CrossRef] [PubMed]
- Bager, F.; Madsen, M.; Christensen, J.; Aarestrup, F.M. Avoparcin Used as a Growth Promoter Is Associated with the Occurrence of Vancomycin-Resistant Enterococcus faecium on Danish Poultry and Pig Farms. Prev. Vet. Med. 1997, 31, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Ralte, V.S.C.; Loganathan, A.; Manohar, P.; Sailo, C.V.; Sanga, Z.; Ralte, L.; Zothanzama, J.; Leptihn, S.; Nachimuthu, R.; Kumar, N.S. The Emergence of Carbapenem-Resistant Gram-Negative Bacteria in Mizoram, Northeast India. Microbiol. Res. 2022, 13, 342–349. [Google Scholar] [CrossRef]
- Hasan, S.M.; Ibrahim, K.S. Molecular Characterization of Extended Spectrum β -Lactamase (ESBL) and Virulence Gene-Factors in Uropathogenic Escherichia coli (UPEC) in Children in Duhok City, Kurdistan. Antibiotics 2022, 11, 1246. [Google Scholar] [CrossRef] [PubMed]
- Rupp, M.E.; Fey, P.D. Extended Spectrum β-Lactamase (ESBL)-Producing Enterobacteriaceae. Drugs 2003, 63, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Zeng, Z.; Xiao, X.; Ding, Y.; Deng, J.; Wei, Y.; Liu, J. Genomic and Phenotypic Characterization of a Colistin-Resistant Escherichia coli Isolate Co-Harboring blaNDM-5, blaOXA-1, and blaCTX-M-55 Isolated from Urine. Infect. Drug Resist. 2022, 15, 1329–1343. [Google Scholar] [CrossRef]
- Carvalho, I.; Carvalho, J.A.; Martínez-Álvarez, S.; Sadi, M.; Capita, R.; Alonso-Calleja, C.; Rabbi, F.; Dapkevicius, M.L.N.E.; Igrejas, G.; Torres, C.; et al. Characterization of ESBL-Producing Escherichia coli and Klebsiella pneumoniae Isolated from Clinical Samples in a Northern Portuguese Hospital: Predominance of CTX-M-15 and High Genetic Diversity. Microorganisms 2021, 9, 1914. [Google Scholar] [CrossRef]
- Ojdana, D.; Sacha, P.; Wieczorek, P.; Czaban, S.; Michalska, A.; Jaworowska, J.; Jurczak, A.; Poniatowski, B.; Tryniszewska, E. The Occurrence of blaCTX-M, blaSHV, and blaTEM Genes in Extended-Spectrum β -Lactamase-Positive Strains of Klebsiella pneumoniae, Escherichia coli, and Proteus mirabilis in Poland. Int. J. Antibiot. 2014, 2014, 935842. [Google Scholar] [CrossRef]
- Ghenea, A.E.; Zlatian, O.M.; Cristea, O.M.; Ungureanu, A.; Mititelu, R.R.; Balasoiu, A.T.; Vasile, C.M.; Salan, A.I.; Iliuta, D.; Popescu, M.; et al. TEM,CTX-M,SHV Genes in ESBL-Producing Escherichia coli and Klebsiella pneumoniae Isolated from Clinical Samples in a County Clinical Emergency Hospital Romania-Predominance of CTX-M-15. Antibiotics 2022, 11, 503. [Google Scholar] [CrossRef]
- Pimenta, A.C.; Fernandes, R.; Moreira, I.S. Evolution of Drug Resistance: Insight on TEM β-Lactamases Structure and Activity and β-Lactam Antibiotics. Mini-Rev. Med. Chem. 2014, 14, 111–122. [Google Scholar] [CrossRef]
- Ahmed, D.; Ud-Din, A.I.M.; Wahid, S.U.H.; Mazumder, R.; Nahar, K.; Hossain, A. Emergence of blaTEM Type Extended-Spectrum β-Lactamase Producing Salmonella Spp. in the Urban Area of Bangladesh. Int. Sch. Res. Not. 2014, 2014, 715310. [Google Scholar] [CrossRef]
- Pishtiwan, A.H.; Khadija, K.M. Prevalence of blaTEM, blaSHV, and blaCTX-M Genes among ESBL-Producing Klebsiella pneumoniae and Escherichia coli Isolated from Thalassemia Patients in Erbil, Iraq. Mediterr. J. Hematol. Infect. Dis. 2019, 11, e2019041. [Google Scholar] [CrossRef] [PubMed]
- Grinsted, J.; De La Cruz, F. The Tn21 Subgroup of Bacterial Transposable Elements. Plasmid 1990, 24, 163–189. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef]
- Malek, M.M.; Amer, F.A.; Allam, A.A.; El-Sokkary, R.H.; Gheith, T.; Arafa, M.A. Occurrence of Classes I and II Integrons in Enterobacteriaceae Collected from Zagazig University Hospitals, Egypt. Front. Microbiol. 2015, 6, 601. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xia, R.; Han, N.; Xu, H. Genetic Diversity Analyses of Class 1 Integrons and Their Associated Antimicrobial Resistance Genes in Enterobacteriaceae Strains Recovered from Aquatic Habitats in China. Lett. Appl. Microbiol. 2011, 52, 667–675. [Google Scholar] [CrossRef]
- Zinner, S.H.; Mayer, K.H. 33-Sulfonamides and Trimethoprim. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 8th ed.; Bennett, J.E., Dolin, R., Blaser, M.J., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2015. [Google Scholar] [CrossRef]
- Hansen, W.; Yourassowsky, E. Detection of beta-glucuronidase in lactose-fermenting members of the family Enterobacteriaceae and its presence in bacterial urine cultures. J. Clin. Microbiol. 1984, 20, 1177–1179. [Google Scholar] [CrossRef]
- Prescott, L.M.; Harley, J.P. Harley Prescott: Laboratory Exercises in Microbiology, 5th ed.; The McGraw-Hill: New York, NY, USA, 2002. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; twenty-fifth informational supplement; CLSI Document M100-S25; Clinical and Laboratory Standards Institute: Malvern, PA, USA, 2015; Volume 32. [Google Scholar]
- Versalovic, J.; Koeuth, T.; Lupski, R. Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.; Quattro, J. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Dierikx, C.M.; van Duijkeren, E.; Schoormans, A.H.; van Essen-Zandbergen, A.; Veldman, K.; Kant, A.; Huijsdens, X.W.; van der Zwaluw, K.; Wagenaar, J.A.; Mevius, D.J. Occurrence and characteristics of extended-spectrum-β-lactamase-and AmpC-producing clinical isolates derived from companion animals and horses. J. Antimicrob. Chemother. 2012, 67, 1368–1374. [Google Scholar] [CrossRef]
- Dierikx, C.; van Essen-Zandbergen, A.; Veldman, K.; Smith, H.; Mevius, D. Increased detection of extended spectrum beta-lactamase producing Salmonella enterica and Escherichia coli isolates from poultry. Vet. Microbiol. 2010, 145, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef]
- Ercolini, D.; Hill, P.; Dodd, C.E.R. Bacterial community structure and location in Stilton cheese. Appl. Environ. Microbiol. 2003, 69, 3540–3548. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]





| Isolates ID | Gene | Variant Type | Accession Number |
|---|---|---|---|
| H1B | blaCTX-M | blaCTX-M-55 | OP589230 |
| H2B | blaCTX-M | blaCTX-M-258 | OP679875 |
| H3B | blaCTX-M | blaCTX-M-15 | OP689692 |
| H4B | blaTEM | blaTEM-1 | OP703167 |
| H5B | blaCTX-M/blaTEM | blaCTX-M-15/blaTEM-1 | OP620950/OP703168 |
| H8B | blaCTX-M | blaCTX-M-254 | OP649440 |
| H10B | 16S rDNA | - | OP627530 |
| H15B | blaCTX-M | blaCTX-M-15 | OP620949 |
| H20U | blaTEM | blaTEM-1 | OP723109 |
| H22U | blaCTX-M/blaTEM | blaCTX-M-14/blaTEM-1 | OP679876/OP723108 |
| H24U | blaTEM | blaTEM-30 | OP703169 |
| H25R | blaTEM | blaTEM-32 | OP620947 |
| H26B | blaCTX-M/blaTEM | blaCTX-M-15/blaTEM-1 | OP689690/OP703166 |
| H27B | blaCTX-M | blaCTX-M-55 | OP689691 |
| H32F | blaCTX-M | blaCTX-M-245 | OP620948 |
| H33F | blaTEM | blaTEM-1 | OP649441 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, D.R.; Dodd, C.E.R.; Stekel, D.J.; Meshioye, R.T.; Diggle, M.; Lister, M.; Hobman, J.L. Multidrug-Resistant ESBL-Producing E. coli in Clinical Samples from the UK. Antibiotics 2023, 12, 169. https://doi.org/10.3390/antibiotics12010169
Ibrahim DR, Dodd CER, Stekel DJ, Meshioye RT, Diggle M, Lister M, Hobman JL. Multidrug-Resistant ESBL-Producing E. coli in Clinical Samples from the UK. Antibiotics. 2023; 12(1):169. https://doi.org/10.3390/antibiotics12010169
Chicago/Turabian StyleIbrahim, Delveen R., Christine E. R. Dodd, Dov J. Stekel, Remilekun T. Meshioye, Mathew Diggle, Michelle Lister, and Jon L. Hobman. 2023. "Multidrug-Resistant ESBL-Producing E. coli in Clinical Samples from the UK" Antibiotics 12, no. 1: 169. https://doi.org/10.3390/antibiotics12010169
APA StyleIbrahim, D. R., Dodd, C. E. R., Stekel, D. J., Meshioye, R. T., Diggle, M., Lister, M., & Hobman, J. L. (2023). Multidrug-Resistant ESBL-Producing E. coli in Clinical Samples from the UK. Antibiotics, 12(1), 169. https://doi.org/10.3390/antibiotics12010169