
Novel Megaplasmid Driving NDM-1-Mediated Carbapenem Resistance in Klebsiella pneumoniae ST1588 in South America

 ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paterson, D.L. Resistance in Gram-Negative Bacteria: Enterobacteriaceae. Am. J. Med. 2006, 119, S20–S28. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Chen, L.; Mathema, B.; Chavda, K.D.; DeLeo, F.R.; Bonomo, R.A.; Kreiswirth, B.N. Carbapenemase-Producing Klebsiella pneumoniae: Molecular and Genetic Decoding. Trends Microbiol. 2014, 22, 686–696. [Google Scholar] [CrossRef]
- Safavi, M.; Bostanshirin, N.; Hajikhani, B.; Yaslianifard, S.; van Belkum, A.; Goudarzi, M.; Hashemi, A.; Darban-Sarokhalil, D.; Dadashi, M. Global Genotype Distribution of Human Clinical Isolates of New Delhi Metallo-β-Lactamase-Producing Klebsiella pneumoniae; A Systematic Review. J. Glob. Antimicrob. Resist. 2020, 23, 420–429. [Google Scholar] [CrossRef]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; Mcnally, A.; Zong, Z. NDM Metallo-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef]
- Zhai, Y.; Lee, S.; Teng, L.; Ma, Z.; Hilliard, N.B.; May, R.J.; Brown, S.A.; Yu, F.; Desear, K.E.; Cherabuddi, K.; et al. Dissemination Mechanisms of NDM Genes in Hospitalized Patients. JAC-Antimicrob. Resist. 2021, 3, dlab032. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Anabalón, S.; Conceição Neto, C.O.; D’Alincourt Carvalho-Assef, A.P.; Lima, C.A.; Cifuentes, M.; Silva, F.; Barrera, B.; Domínguez, M.; González-Rocha, G.; Bello-Toledo, H. Introduction of NDM-1 and OXA-370 from Brazil into Chile in Strains of Klebsiella pneumoniae Isolated from a Single Patient. Int. J. Infect. Dis. 2019, 81, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.U.; Maryam, L.; Zarrilli, R. Structure, Genetics and Worldwide Spread of New Delhi Metallo-β-Lactamase (NDM): A Threat to Public Health. BMC Microbiol. 2017, 17, 101. [Google Scholar] [CrossRef]
- Yao, H.; Li, A.; Yu, R.; Schwarz, S.; Dong, H.; Du, X.D. Multiple Copies of Bla NDM-5 Located on Conjugative Megaplasmids from Porcine Escherichia coli Sequence Type 218 Isolates. Antimicrob. Agents Chemother. 2020, 64, e02134-19. [Google Scholar] [CrossRef]
- Li, M.; Guan, C.; Song, G.; Gao, X.; Yang, W.; Wang, T.; Zhang, Y. Characterization of a Conjugative Multidrug Resistance IncP-2 Megaplasmid, PPAG5, from a Clinical Pseudomonas aeruginosa Isolate. Microbiol. Spectr. 2022, 10, e0199221. [Google Scholar] [CrossRef]
- Zou, H.; Berglund, B.; Xu, H.; Chi, X.; Zhao, Q.; Zhou, Z.; Xia, H.; Li, X.; Zheng, B. Genetic Characterization and Virulence of a Carbapenem-Resistant Raoultella ornithinolytica Isolated from Well Water Carrying a Novel Megaplasmid Containing bla NDM-1. Environ. Pollut. 2020, 260, 114041. [Google Scholar] [CrossRef] [PubMed]
- Cazares, A.; Moore, M.P.; Hall, J.P.J.; Wright, L.L.; Grimes, M.; Emond-Rhéault, J.G.; Pongchaikul, P.; Santanirand, P.; Levesque, R.C.; Fothergill, J.L.; et al. A Megaplasmid Family Driving Dissemination of Multidrug Resistance in Pseudomonas. Nat. Commun. 2020, 11, 1370. [Google Scholar] [CrossRef] [PubMed]
- Ghaly, T.M.; Paulsen, I.T.; Sajjad, A.; Tetu, S.G.; Gillings, M.R. A Novel Family of Acinetobacter Mega-Plasmids Are Disseminating Multi-Drug Resistance Across the Globe While Acquiring Location-Specific Accessory Genes. Front. Microbiol. 2020, 11, 3058. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 31st ed.; CLSI: Wayne, PA, USA, 2021; ISBN 9781684401048. [Google Scholar]
- Lee, K.; Chong, Y.; Shin, H.B.; Kim, Y.A.; Yong, D.; Yum, J.H. Modified Hodge and EDTA-Disk Synergy Tests to Screen Metallo-Beta-Lactamase-Producing Strains of Pseudomonas and Acinetobacter Species. Clin. Microbiol. Infect. 2001, 7, 88–91. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for Detection of Acquired Carbapenemase Genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Antonopoulos, D.A.; Assaf, R.; Aziz, R.K.; Brettin, T.; Bun, C.; Conrad, N.; Davis, J.J.; Dietrich, E.M.; Disz, T.; Gerdes, S.; et al. PATRIC as a Unique Resource for Studying Antimicrobial Resistance. Brief. Bioinform. 2019, 20, 1094–1102. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Schmartz, G.P.; Hartung, A.; Hirsch, P.; Kern, F.; Fehlmann, T.; Müller, R.; Keller, A. PLSDB: Advancing a Comprehensive Database of Bacterial Plasmids. Nucleic Acids Res. 2022, 50, D273–D278. [Google Scholar] [CrossRef] [PubMed]
- Potron, A.; Poirel, L.; Rondinaud, E.; Nordmann, P. Intercontinental Spread of OXA-48 Beta-Lactamase-Producing Enterobacteriaceae over a 11-Year Period, 2001 to 2011. Euro Surveill. 2013, 18, 20549. [Google Scholar] [CrossRef]
- Rumi, M.V.; Mas, J.; Elena, A.; Cerdeira, L.; Muñoz, M.E.; Lincopan, N.; Gentilini, É.R.; di Conza, J.; Gutkind, G. Co-Occurrence of Clinically Relevant β-Lactamases and MCR-1 Encoding Genes in Escherichia coli from Companion Animals in Argentina. Vet. Microbiol. 2019, 230, 228–234. [Google Scholar] [CrossRef]
- Aydin, F.; Gümüşsoy, K.S.; Atabay, H.I.; Iça, T.; Abay, S. Prevalence and Distribution of Arcobacter Species in Various Sources in Turkey and Molecular Analysis of Isolated Strains by ERIC-PCR. J. Appl. Microbiol. 2007, 103, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Aires, C.A.M.; Pereira, P.S.; de Araujo, C.F.M.; Chagas, T.P.G.; Oliveira, J.C.R.; Buonora, S.N.; Albano, R.M.; Carvalho-Assef, A.P.D.A.; Asensi, M.D. Multiclonal Expansion of Klebsiella pneumoniae Isolates Producing NDM-1 in Rio de Janeiro, Brazil. Antimicrob. Agents Chemother. 2017, 61, e01048-16. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Yang, X.; Chan, E.W.C.; Zhang, R.; Chen, S. Klebsiella Species: Taxonomy, Hypervirulence and Multidrug Resistance. EBioMedicine 2022, 79, 103998. [Google Scholar] [CrossRef]
- Schandel, K.A.; Muller, M.M.; Webster, R.E. Localization of TraC, a Protein Involved in Assembly of the F Conjugative Pilus. J. Bacteriol. 1992, 174, 3800–3806. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Baomo, L.; Lili, S.; Moran, R.A.; van Schaik, W.; Chao, Z. Temperature-Regulated IncX3 Plasmid Characteristics and the Role of Plasmid-Encoded H-NS in Thermoregulation. Front. Microbiol. 2022, 1, 765492. [Google Scholar] [CrossRef]
- Bragagnolo, N.; Rodriguez, C.; Samari-Kermani, N.; Fours, A.; Korouzhdehi, M.; Lysenko, R.; Audette, G.F. Protein Dynamics in F-like Bacterial Conjugation. Biomedicines 2020, 8, 362. [Google Scholar] [CrossRef] [PubMed]
- Newire, E.A.; Ahmed, S.F.; House, B.; Valiente, E.; Pimentel, G. Detection of New SHV-12, SHV-5 and SHV-2a Variants of Extended Spectrum Beta-Lactamase in Klebsiella pneumoniae in Egypt. Ann. Clin. Microbiol. Antimicrob. 2013, 12, 16. [Google Scholar] [CrossRef]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a New Antibiotic Resistance Mechanism in India, Pakistan, and the UK: A Molecular, Biological, and Epidemiological Study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of NDM-1 Positive Bacteria in the New Delhi Environment and Its Implications for Human Health: An Environmental Point Prevalence Study. Lancet Infect. Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Elgorriaga-Islas, E.; Guggiana-Nilo, P.; Domínguez-Yévenes, M.; González-Rocha, G.; Mella-Montecinos, S.; Labarca-Labarca, J.; García-Cañete, P.; Bello-Toledo, H. Prevalence of Plasmid-Mediated Quinolone Resistance Determinant Aac(6′)-Ib-Cr among ESBL Producing Enterobacteria Isolates from Chilean Hospitals. Enferm. Infecc. Microbiol. Clin. 2012, 30, 466–468. [Google Scholar] [CrossRef] [PubMed]
- Potron, A.; Poirel, L.; Nordmann, P. Plasmid-Mediated Transfer of the Bla(NDM-1) Gene in Gram-Negative Rods. FEMS Microbiol. Lett. 2011, 324, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Poirel, L.; Walsh, T.R.; Livermore, D.M. The Emerging NDM Carbapenemases. Trends Microbiol. 2011, 19, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Gunton, J.E.; Gilmour, M.W.; Alonso, G.; Taylor, D.E. Subcellular Localization and Functional Domains of the Coupling Protein, TraG, from IncHI1 Plasmid R27. Microbiology 2005, 151, 3549–3561. [Google Scholar] [CrossRef] [PubMed]
- Gibert, M.; Paytubi, S. Temperature Dependent Control of the R27 Conjugative Plasmid Genes. Front. Mol. Biosci. 2020, 7, 124. [Google Scholar] [CrossRef]
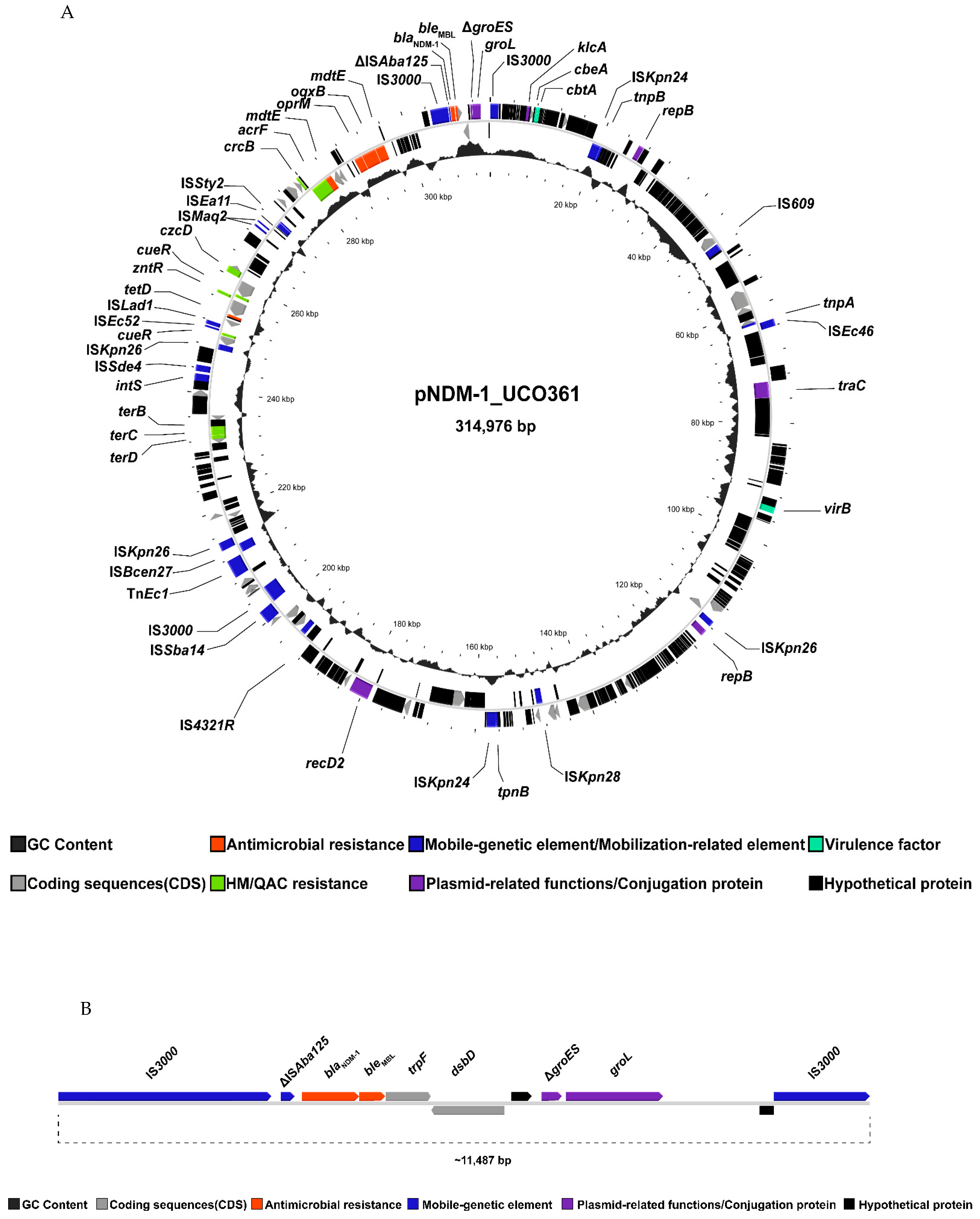

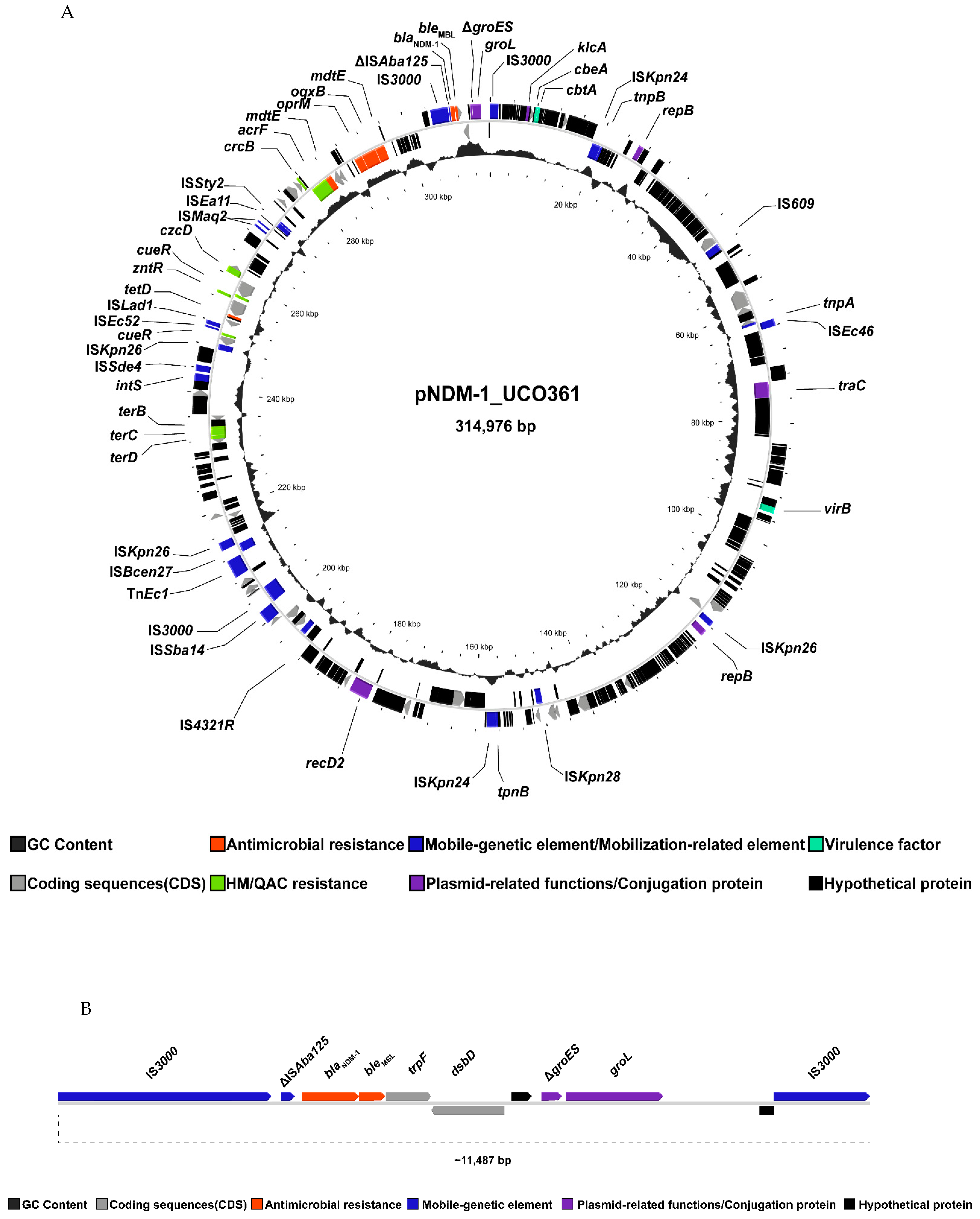{kind=link}
| Strains | MIC (µg/mL) | Resistome | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IPM | MEM | ETP | ATM | CTX | AMK | CIP | β-lactams | aminoglycosides | quinolones | SXT | |
| K. pneumoniae UCO-361 | 16 | >32 | 32 | >32 | >512 | 8 | 2 | blaNDM-1;blaSHV-106; blaCTX-M-15; blaOXA-1; blaTEM-1B | aac(3)-IIa; aac(6′)-Ib-cr; aph(6)-Id; aph(3″)-Ib | qnrB1; oqxA; oqxB; aac(6′)-Ib-cr | dfrA14; sul2 |
| E. coli Tc-01 | 16 | 32 | 16 | 16 | 512 | 2 | 2 | blaNDM-1; blaCTX-M-15; blaOXA-1; blaTEM-1B | aac(6′)-Ib-cr aph(6)-Id; aph(3″)-Ib | qnrB1; aac(6′)-Ib-cr | |
| E. coli J53 | 0.25 | ≤0.06 | 0.016 | ≤0.06 | 0.063 | 1 | 0.002 | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quezada-Aguiluz, M.; Opazo-Capurro, A.; Lincopan, N.; Esposito, F.; Fuga, B.; Mella-Montecino, S.; Riedel, G.; Lima, C.A.; Bello-Toledo, H.; Cifuentes, M.; et al. Novel Megaplasmid Driving NDM-1-Mediated Carbapenem Resistance in Klebsiella pneumoniae ST1588 in South America. Antibiotics 2022, 11, 1207. https://doi.org/10.3390/antibiotics11091207
Quezada-Aguiluz M, Opazo-Capurro A, Lincopan N, Esposito F, Fuga B, Mella-Montecino S, Riedel G, Lima CA, Bello-Toledo H, Cifuentes M, et al. Novel Megaplasmid Driving NDM-1-Mediated Carbapenem Resistance in Klebsiella pneumoniae ST1588 in South America. Antibiotics. 2022; 11(9):1207. https://doi.org/10.3390/antibiotics11091207
Chicago/Turabian StyleQuezada-Aguiluz, Mario, Andrés Opazo-Capurro, Nilton Lincopan, Fernanda Esposito, Bruna Fuga, Sergio Mella-Montecino, Gisela Riedel, Celia A. Lima, Helia Bello-Toledo, Marcela Cifuentes, and et al. 2022. "Novel Megaplasmid Driving NDM-1-Mediated Carbapenem Resistance in Klebsiella pneumoniae ST1588 in South America" Antibiotics 11, no. 9: 1207. https://doi.org/10.3390/antibiotics11091207
APA StyleQuezada-Aguiluz, M., Opazo-Capurro, A., Lincopan, N., Esposito, F., Fuga, B., Mella-Montecino, S., Riedel, G., Lima, C. A., Bello-Toledo, H., Cifuentes, M., Silva-Ojeda, F., Barrera, B., Hormazábal, J. C., & González-Rocha, G. (2022). Novel Megaplasmid Driving NDM-1-Mediated Carbapenem Resistance in Klebsiella pneumoniae ST1588 in South America. Antibiotics, 11(9), 1207. https://doi.org/10.3390/antibiotics11091207