
Global Spread of MCR-Producing Salmonella enterica Isolates



Abstract
:1. Introduction
2. Results
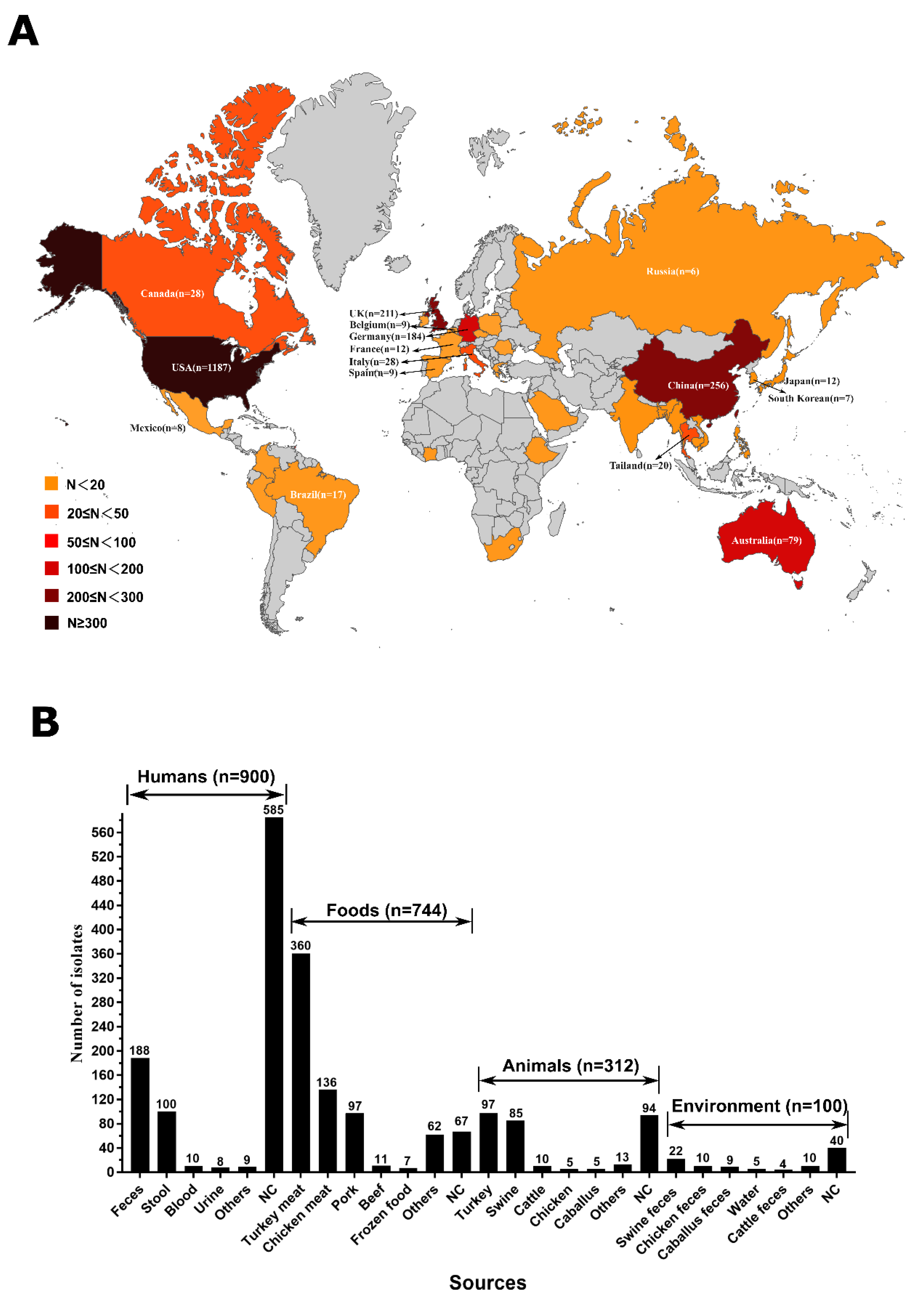
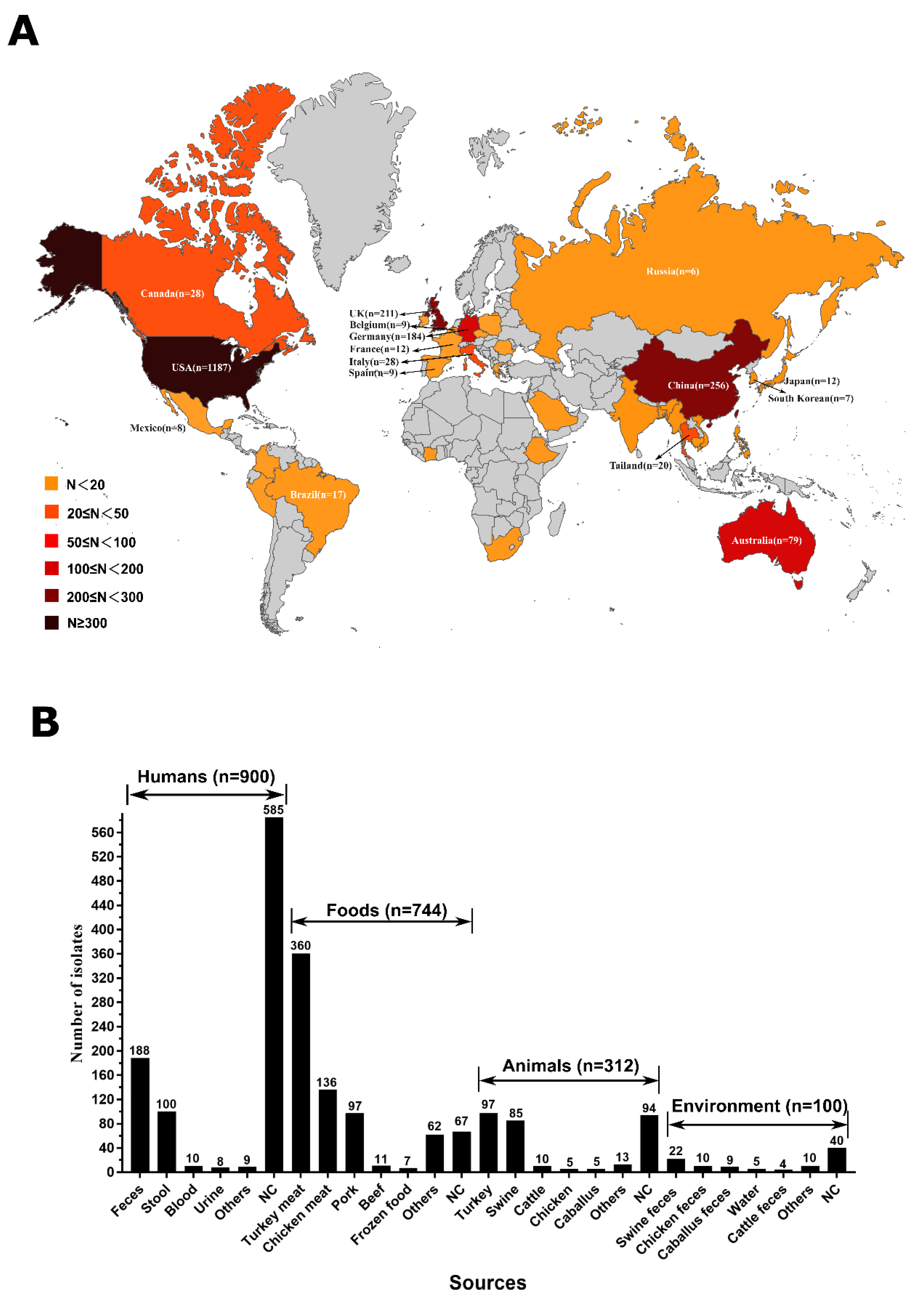
2.1. Widely Spread of mcr-Producing Salmonella Enterica in the Global and Sources Analysis
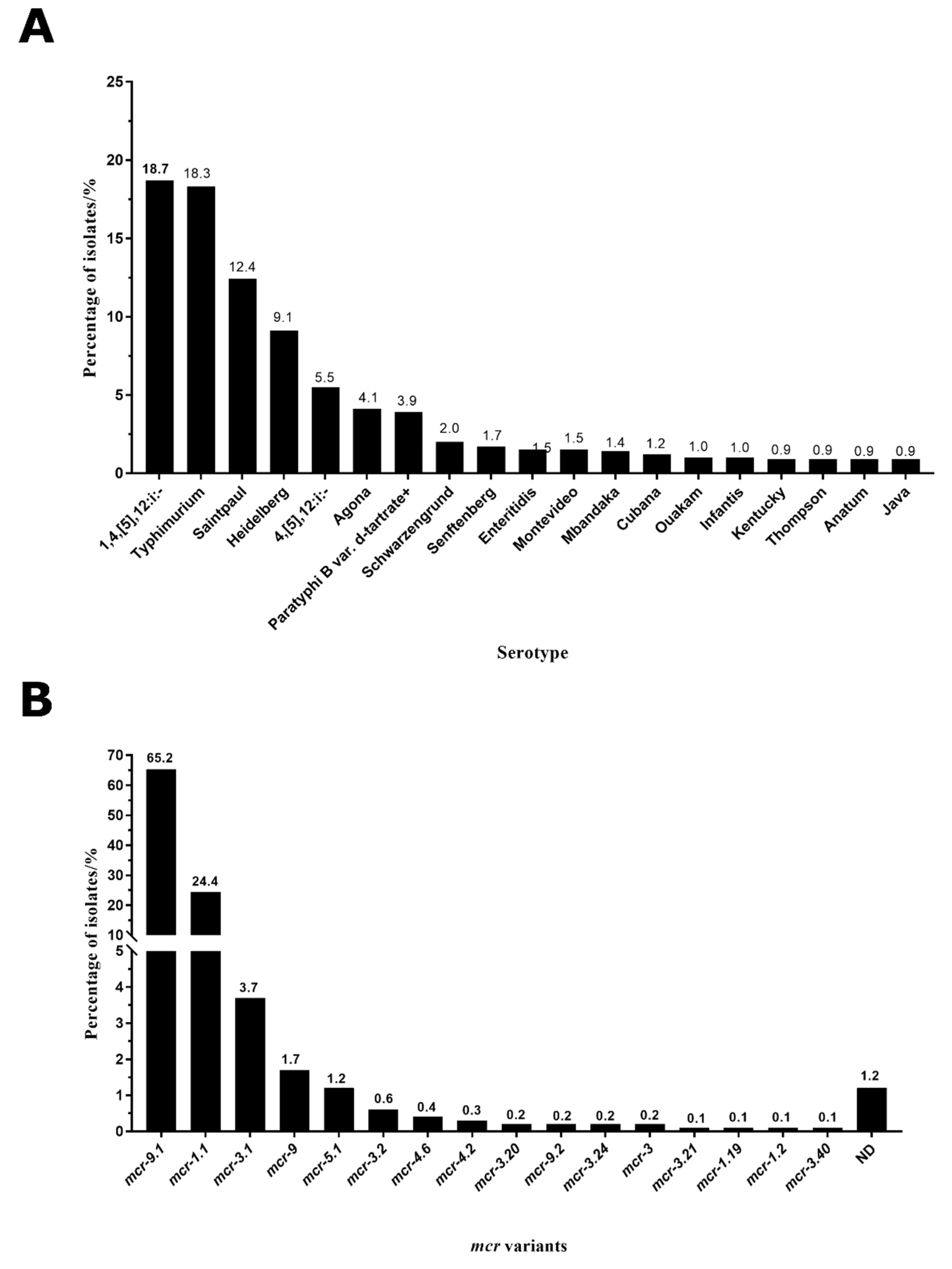
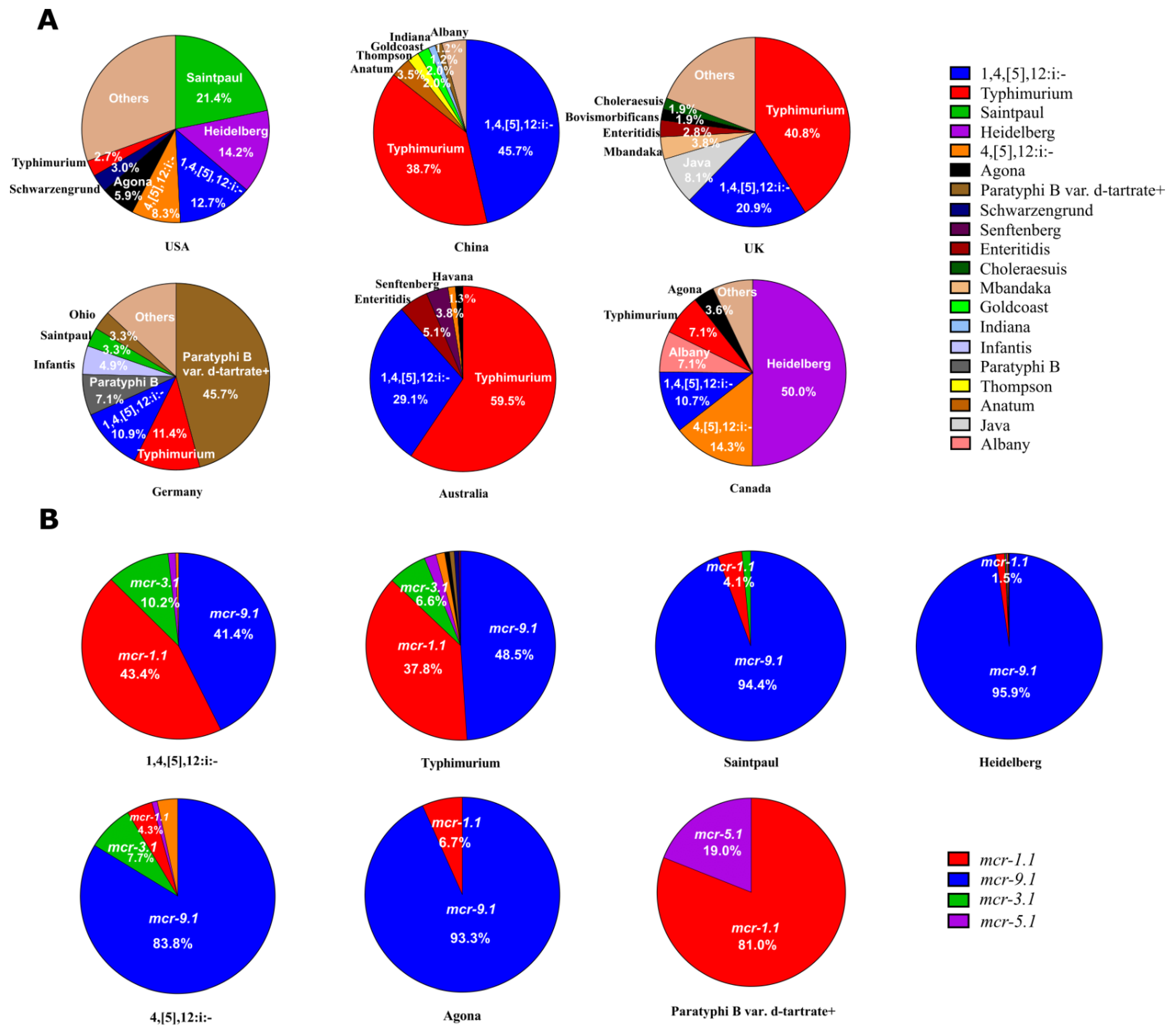
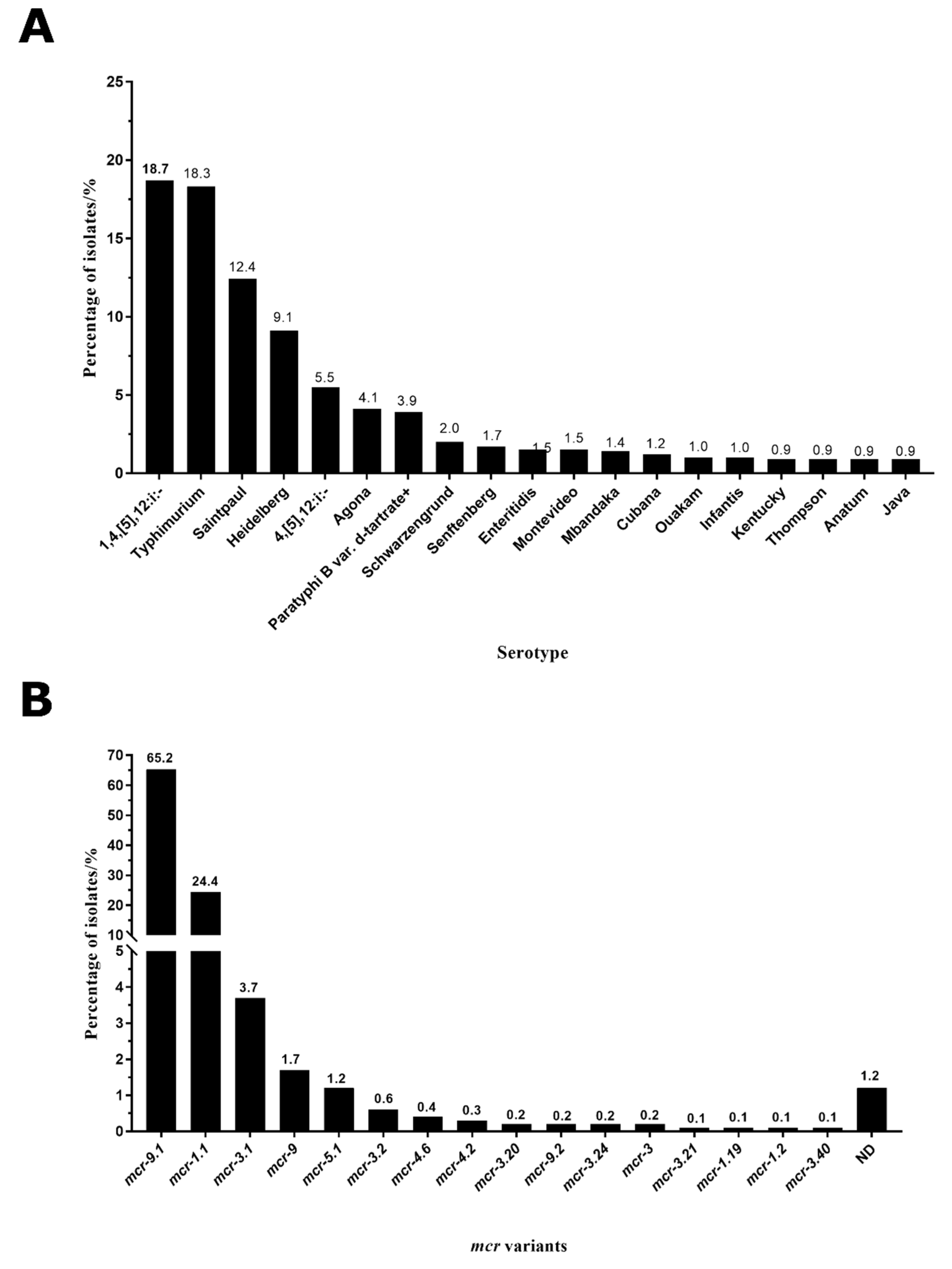
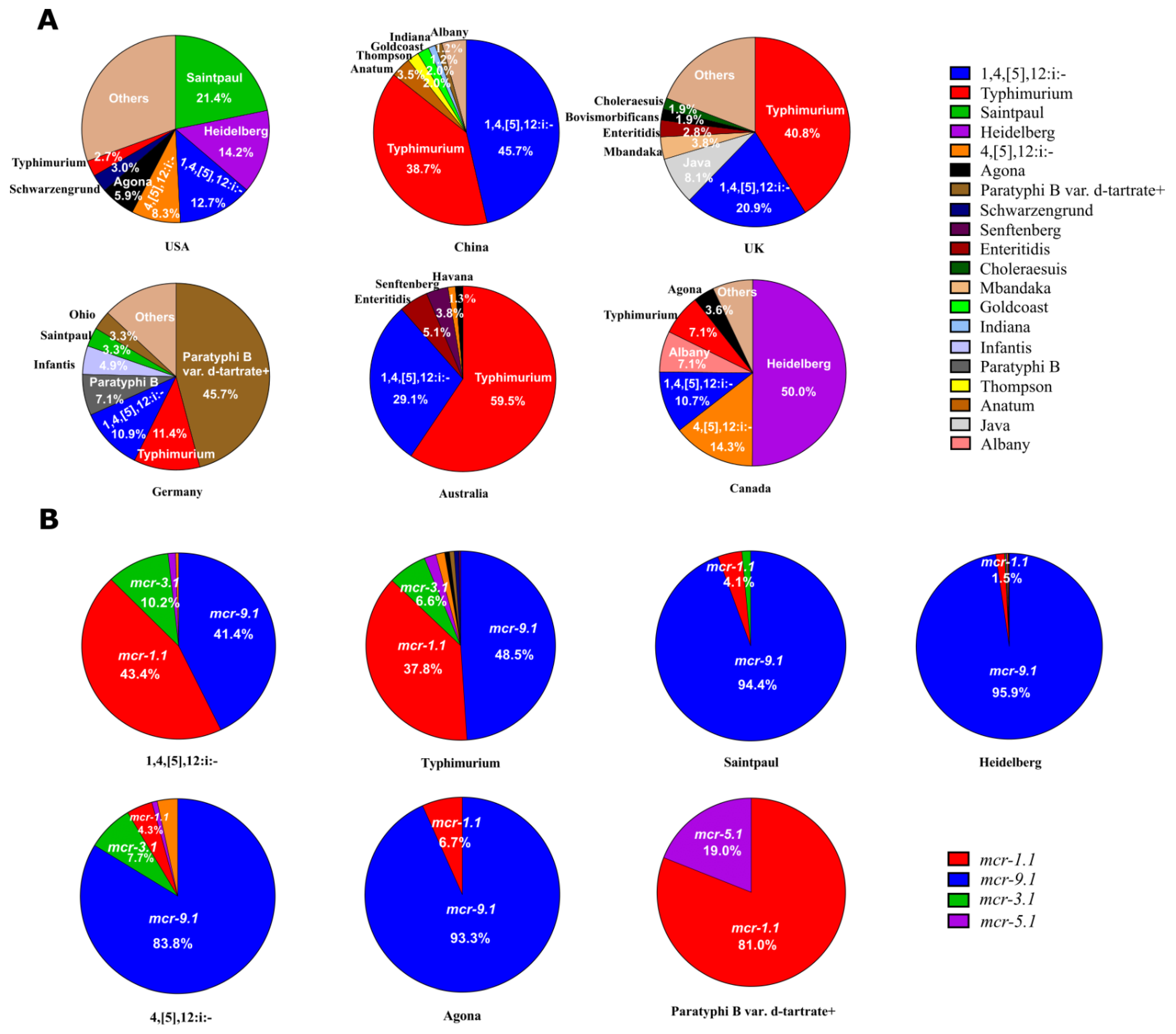
2.2. Typhimurium and 1,4,[5],12:i:- Were the Main Serotypes Carrying mcr Genes in Salmonella
2.3. mcr-9.1 and mcr-1.1 Were the Dominant Variants in Salmonella
2.4. Antimicrobial Resistance (AMR) Genotypes in mcr-Producing Salmonella Isolates
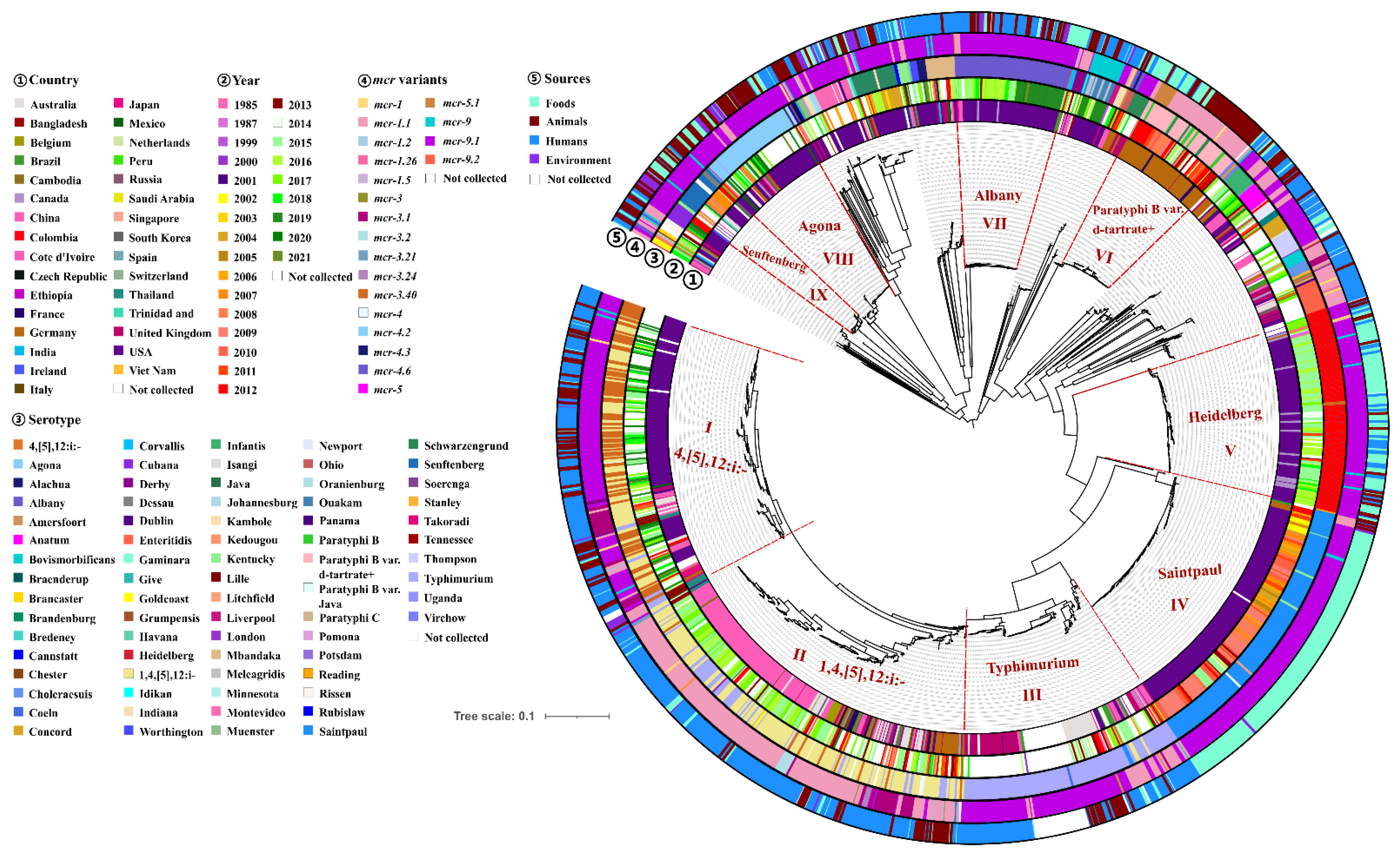
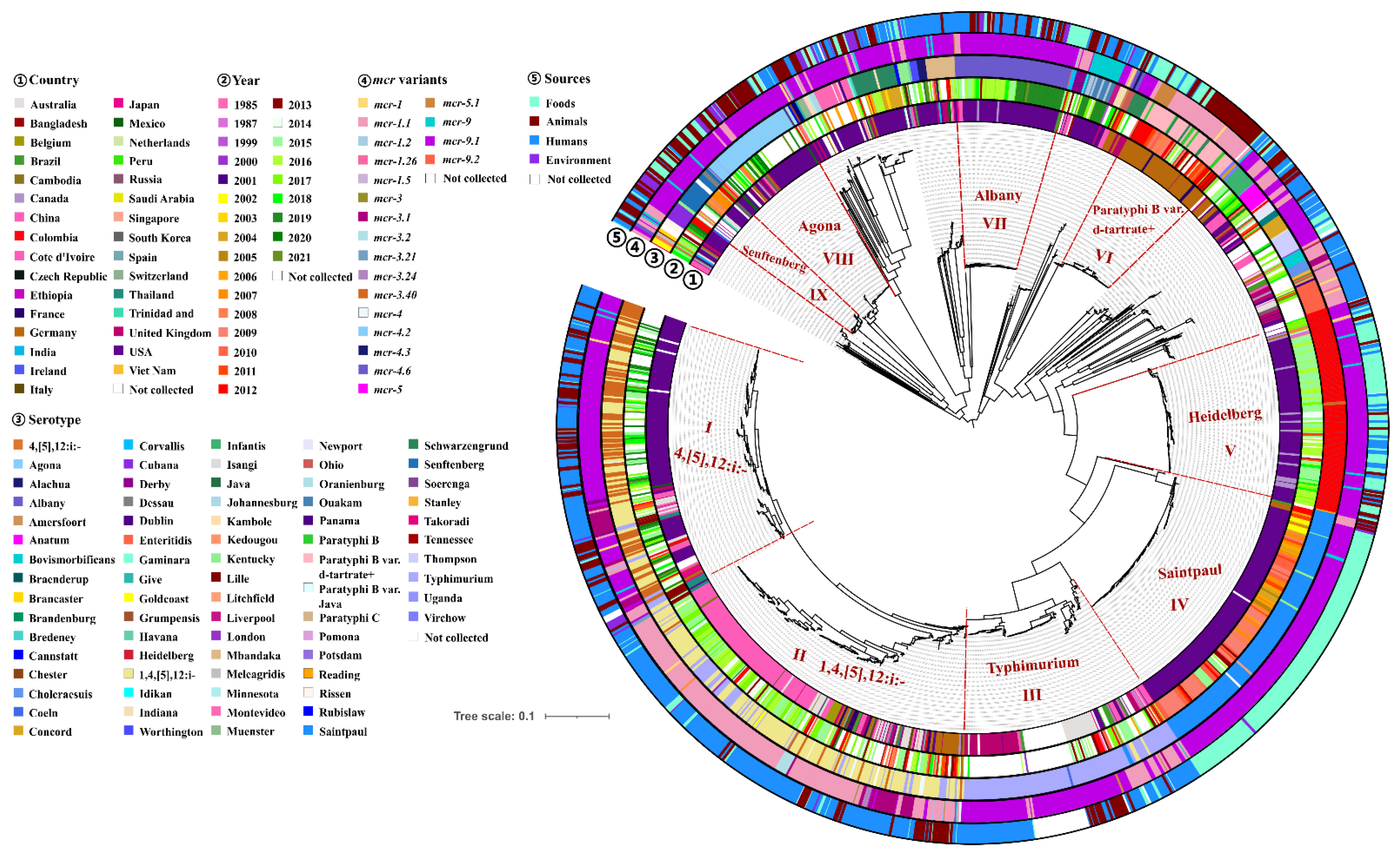
2.5. Phylogenomic Analysis of mcr-Producing Salmonella
3. Discussion
4. Materials and Methods
4.1. Salmonella Genomes Collected
4.2. The Identification of mcr Variants and Serotypes
4.3. Phylogenetic Analysis
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Portes, A.B.; Rodrigues, G.; Leitão, M.P.; Ferrari, R.; Conte Junior, C.A.; Panzenhagen, P. Global Distribution of Plasmid-Mediated Colistin Resistance Mcr Gene in Salmonella: A Systematic Review. J. Appl. Microbiol. 2022, 132, 872–889. [Google Scholar] [CrossRef]
- Wang, R.; van Dorp, L.; Shaw, L.P.; Bradley, P.; Wang, Q.; Wang, X.; Jin, L.; Zhang, Q.; Liu, Y.; Rieux, A.; et al. The Global Distribution and Spread of the Mobilized Colistin Resistance Gene Mcr-1. Nat. Commun. 2018, 9, 1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nang, S.C.; Li, J.; Velkov, T. The Rise and Spread of Mcr Plasmid-Mediated Polymyxin Resistance. Crit. Rev. Microbiol. 2019, 45, 131–161. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Mechanisms of Polymyxin Resistance: Acquired and Intrinsic Resistance in Bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Hussein, N.H.; AL-Kadmy, I.M.S.; Taha, B.M.; Hussein, J.D. Mobilized Colistin Resistance (Mcr) Genes from 1 to 10: A Comprehensive Review. Mol. Biol. Rep. 2021, 48, 2897–2907. [Google Scholar] [CrossRef]
- Poirel, L.; Kieffer, N.; Liassine, N.; Thanh, D.; Nordmann, P. Plasmid-Mediated Carbapenem and Colistin Resistance in a Clinical Isolate of Escherichia coli. Lancet Infect. Dis. 2016, 16, 281. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Chen, L.; Tang, Y.-W.; Kreiswirth, B.N. Emergence of the Mcr-1 Colistin Resistance Gene in Carbapenem-Resistant Enterobacteriaceae. Lancet Infect. Dis. 2016, 16, 287–288. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; Doi, Y.; Zeng, L.; Lv, L.; Liu, J.-H. Carbapenem-Resistant and Colistin-Resistant Escherichia coli Co-Producing NDM-9 and MCR-1. Lancet Infect. Dis. 2016, 16, 288–289. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Zhou, H.; Xu, J.; Wang, Y.; Zhang, Q.; Walsh, T.R.; Shao, B.; Wu, C.; Hu, Y.; Yang, L.; et al. Anthropogenic and Environmental Factors Associated with High Incidence of Mcr-1 Carriage in Humans across China. Nat. Microbiol. 2018, 3, 1054–1062. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, G.-B.; Zhang, R.; Shen, Y.; Tyrrell, J.M.; Huang, X.; Zhou, H.; Lei, L.; Li, H.-Y.; Doi, Y.; et al. Prevalence, Risk Factors, Outcomes, and Molecular Epidemiology of Mcr-1-Positive Enterobacteriaceae in Patients and Healthy Adults from China: An Epidemiological and Clinical Study. Lancet Infect. Dis. 2017, 17, 390–399. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, R.; Li, J.; Wu, Z.; Yin, W.; Schwarz, S.; Tyrrell, J.M.; Zheng, Y.; Wang, S.; Shen, Z.; et al. Comprehensive Resistome Analysis Reveals the Prevalence of NDM and MCR-1 in Chinese Poultry Production. Nat. Microbiol. 2017, 2, 16260. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Li, X.; Chen, Y.; Jiang, Y.; Zhou, Z.; Zhang, H.; Sun, L.; Ruan, Z.; Feng, Y.; Akova, M.; et al. Prevalence of Mcr-1 in Escherichia coli and Klebsiella Pneumoniae Recovered from Bloodstream Infections in China: A Multicentre Longitudinal Study. Lancet Infect. Dis. 2017, 17, 400–410. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Zhang, R.; Chen, Y.; Shen, Y.; Hu, F.; Liu, D.; Lu, J.; Guo, Y.; Xia, X.; et al. Changes in Colistin Resistance and Mcr-1 Abundance in Escherichia coli of Animal and Human Origins Following the Ban of Colistin-Positive Additives in China: An Epidemiological Comparative Study. Lancet Infect. Dis. 2020, 20, 1161–1171. [Google Scholar] [CrossRef]
- Shen, Z.; Wang, Y.; Shen, Y.; Shen, J.; Wu, C. Early Emergence of Mcr-1 in Escherichia coli from Food-Producing Animals. Lancet Infect. Dis. 2016, 16, 293. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Wan, Y.; Du, P.; Bai, L. The Epidemiology of Monophasic Salmonella Typhimurium. Foodborne Pathog. Dis. 2020, 17, 87–97. [Google Scholar] [CrossRef]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid Phylogenetic Analysis of Large Samples of Recombinant Bacterial Whole Genome Sequences Using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Resistance Determinants | Numbers | Ratio/% | |
|---|---|---|---|
| Carbapenems | blaNDM-1 | 1 | 0.0 |
| Tigecycline | tet(X4) | 1 | 0.0 |
| Aminoglycosides | dfrA12 | 232 | 10.2 |
| - | dfrA14 | 42 | 1.8 |
| - | dfrA17 | 21 | 0.9 |
| - | dfrA16 | 59 | 2.6 |
| - | dfrA19 | 382 | 16.8 |
| - | dfrA23 | 19 | 0.8 |
| - | dfrA5 | 35 | 1.5 |
| - | dfrA1 | 212 | 9.3 |
| - | aph(6)-Id | 1023 | 44.9 |
| - | aph(3″)-Ib | 1143 | 50.2 |
| - | aph(3′)-Ia | 685 | 30.1 |
| - | aph(4)-Ia | 246 | 10.8 |
| - | aadA2 | 931 | 40.9 |
| - | aadA1 | 1063 | 46.6 |
| - | aadA22 | 15 | 0.7 |
| - | aadA5 | 35 | 1.5 |
| - | aac(6′)-IIc | 357 | 15.7 |
| - | aac(6′)-Ib-cr5 | 85 | 3.7 |
| - | aac(3)-IId | 129 | 5.7 |
| - | aac(3)-IVa | 212 | 9.3 |
| - | aac(3)-IIe | 12 | 0.5 |
| - | aac(3)-IIg | 160 | 7.0 |
| β-Lactamase | blaCTX-M-14 | 136 | 6.0 |
| - | blaCTX-M-55 | 90 | 3.9 |
| - | blaCTX-M-65 | 14 | 0.6 |
| - | blaCTX-M-15 | 19 | 0.8 |
| - | blaCTX-M-3 | 13 | 0.6 |
| - | blaCTX-M-9 | 109 | 4.8 |
| - | blaCTX-M-130 | 3 | 0.1 |
| - | blaCTX-M-2 | 4 | 0.2 |
| - | blaCMY-2 | 46 | 2.0 |
| - | blaCMY-16 | 1 | 0.0 |
| - | blaTEM | 1315 | 57.7 |
| - | blaSHV-12 | 307 | 13.5 |
| - | blaDHA-1 | 33 | 1.4 |
| - | blaOXA-1 | 115 | 5.0 |
| - | blaOXA-10 | 3 | 0.1 |
| Rifampicin | arr-3 | 86 | 3.8 |
| Chloramphenicol | cmlA1 | 292 | 12.8 |
| - | catB3 | 76 | 3.3 |
| - | catA2 | 296 | 13.0 |
| Macrolides | mph(A) | 113 | 5.0 |
| - | mef(B) | 7 | 0.3 |
| - | erm(B) | 10 | 0.4 |
| - | erm(42) | 5 | 0.2 |
| Fosfomycin | fosA7 | 326 | 14.3 |
| - | fosA3 | 137 | 6.0 |
| - | fosA4 | 1 | 0.0 |
| Florfenicol | floR | 480 | 21.1 |
| Lincomycin | lnu(F) | 64 | 2.8 |
| - | lnu(G) | 8 | 0.4 |
| Sulfonamide | sul3 | 334 | 14.7 |
| - | sul2 | 1220 | 53.5 |
| - | sul1 | 1184 | 52.0 |
| Tetracyclines | tet(A) | 578 | 25.4 |
| - | tet(M) | 71 | 3.1 |
| - | tet(B) | 1099 | 48.2 |
| - | tet(C) | 34 | 1.5 |
| - | tet(D) | 368 | 16.1 |
| Quinolones | qnrS1 | 160 | 7.0 |
| - | oqxA | 180 | 7.9 |
| - | oqxB | 181 | 7.9 |
| - | qnrS2 | 10 | 0.4 |
| - | qnrB4 | 32 | 1.4 |
| - | qnrB19 | 60 | 2.6 |
| - | qnrB2 | 214 | 9.4 |
| AMR mutations | gyrA_D87G | 128 | 5.6 |
| - | gyrA_S83Y | 58 | 2.5 |
| - | gyrA_D87N | 98 | 4.3 |
| - | gyrA_G81C | 1 | 0.0 |
| - | gyrA_S83F | 57 | 2.5 |
| - | parC_S80I | 12 | 0.5 |
| - | parC_S80R | 5 | 0.2 |
| - | parE_S458P | 1 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Tian, X.; Shi, C. Global Spread of MCR-Producing Salmonella enterica Isolates. Antibiotics 2022, 11, 998. https://doi.org/10.3390/antibiotics11080998
Zhang Z, Tian X, Shi C. Global Spread of MCR-Producing Salmonella enterica Isolates. Antibiotics. 2022; 11(8):998. https://doi.org/10.3390/antibiotics11080998
Chicago/Turabian StyleZhang, Zengfeng, Xiaorong Tian, and Chunlei Shi. 2022. "Global Spread of MCR-Producing Salmonella enterica Isolates" Antibiotics 11, no. 8: 998. https://doi.org/10.3390/antibiotics11080998
APA StyleZhang, Z., Tian, X., & Shi, C. (2022). Global Spread of MCR-Producing Salmonella enterica Isolates. Antibiotics, 11(8), 998. https://doi.org/10.3390/antibiotics11080998