
Identification of Mobile Colistin Resistance Gene mcr-10 in Disinfectant and Antibiotic Resistant Escherichia coli from Disinfected Tableware


Abstract
1. Introduction
2. Results
2.1. Strain Identification and Antimicrobial Susceptibility
2.2. ARG Characterization of Multidrug-Resistant Positive E. coli
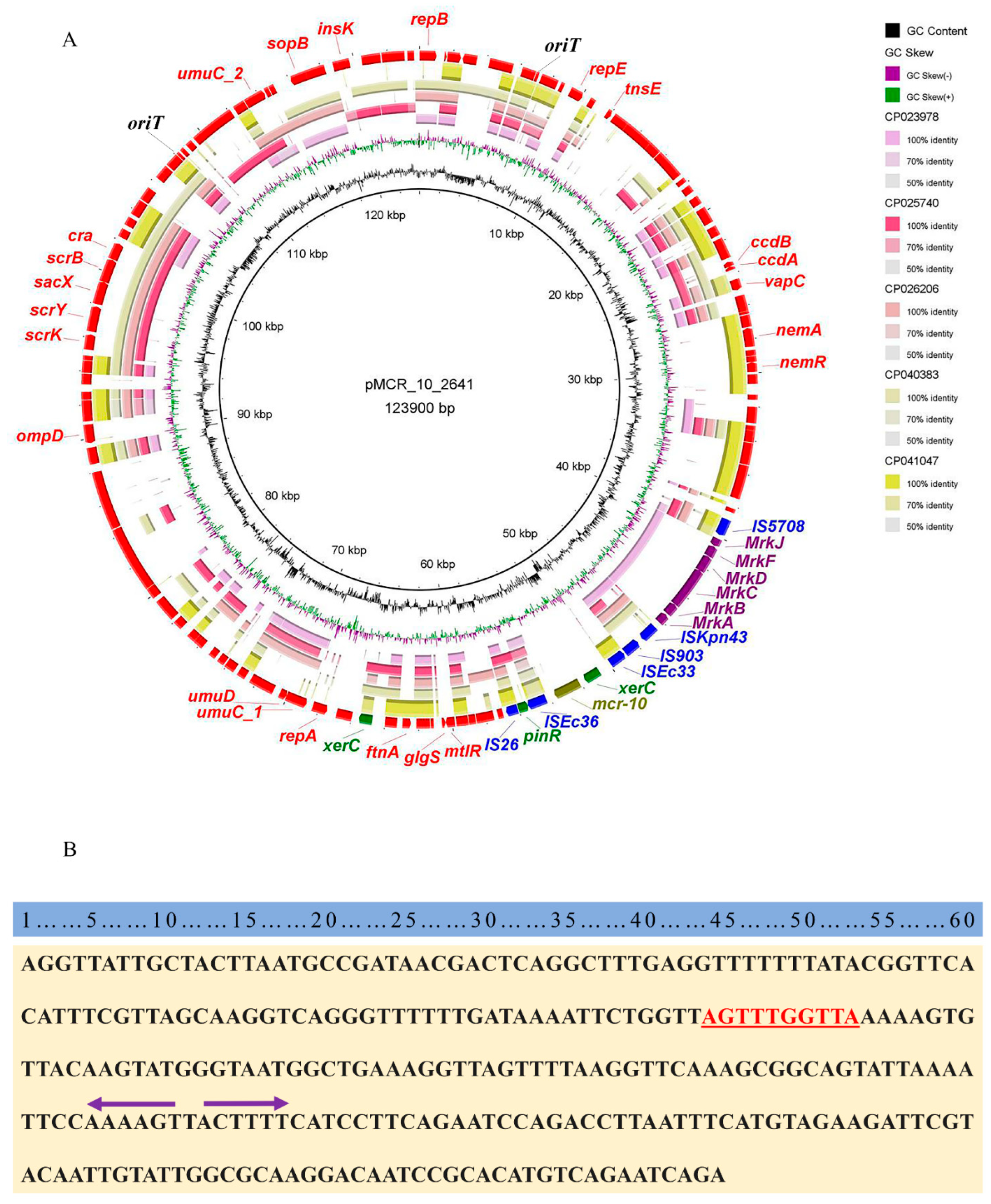
2.3. Genetic Characterization of Colistin-Resistant E. coli EC2641
2.4. Conjugation of mcr-10 among E. coli Isolate EC2641 under Laboratory Conditions
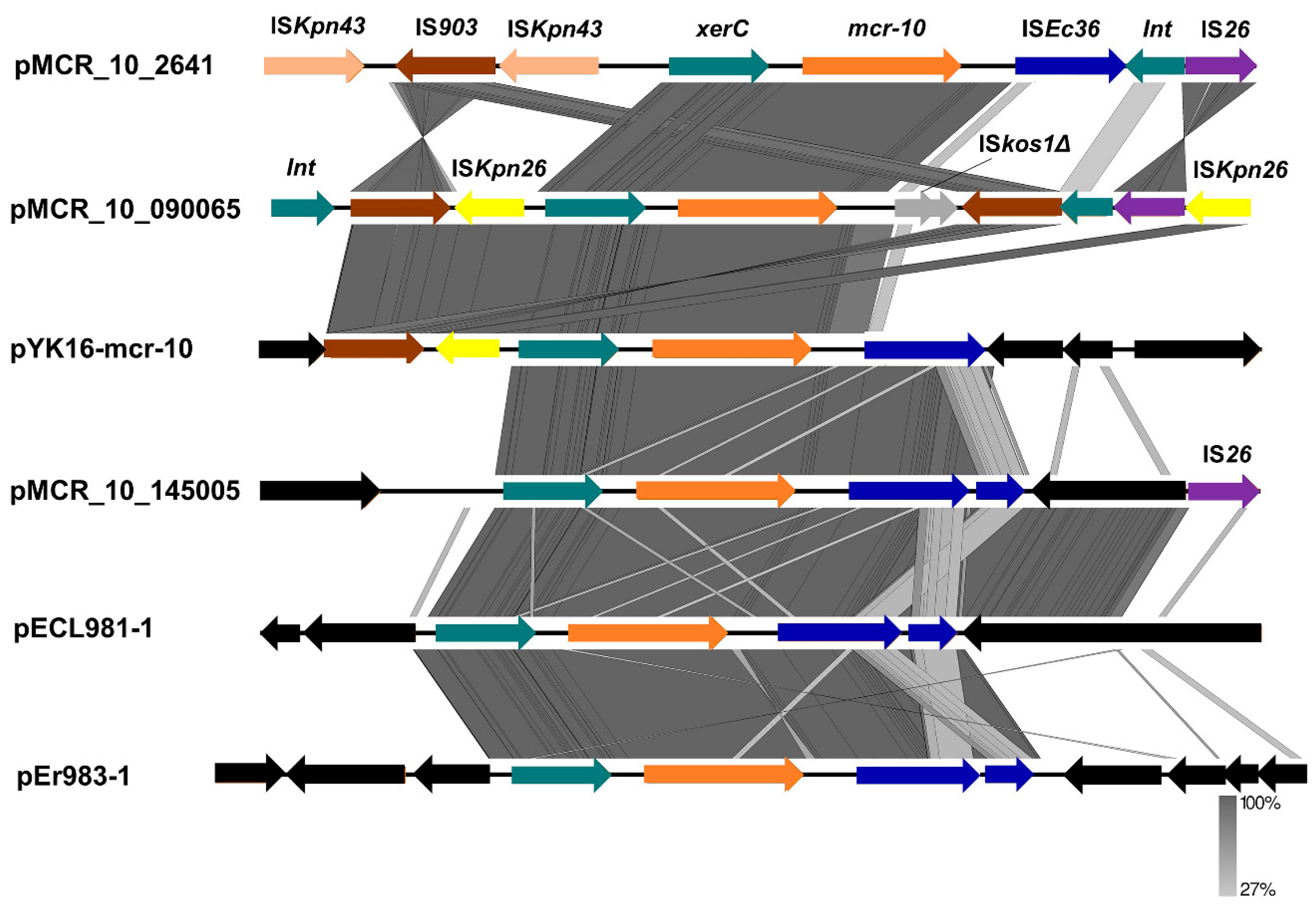
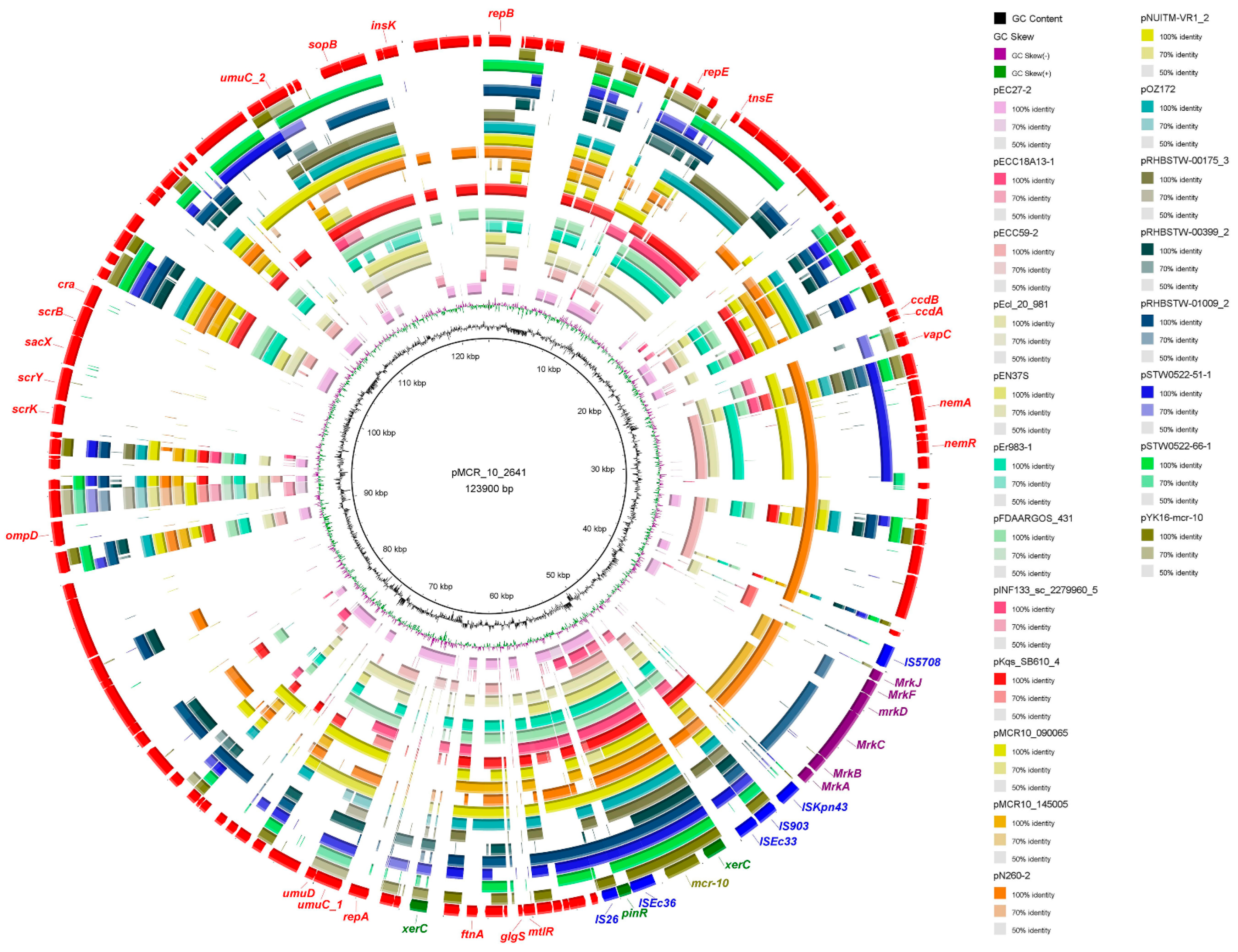
2.5. Genetic Characterization of mcr-10 Carrying Plasmids
2.6. Upregulation of mcr-10 Expression at Subinhibitory Concentrations of Colistin
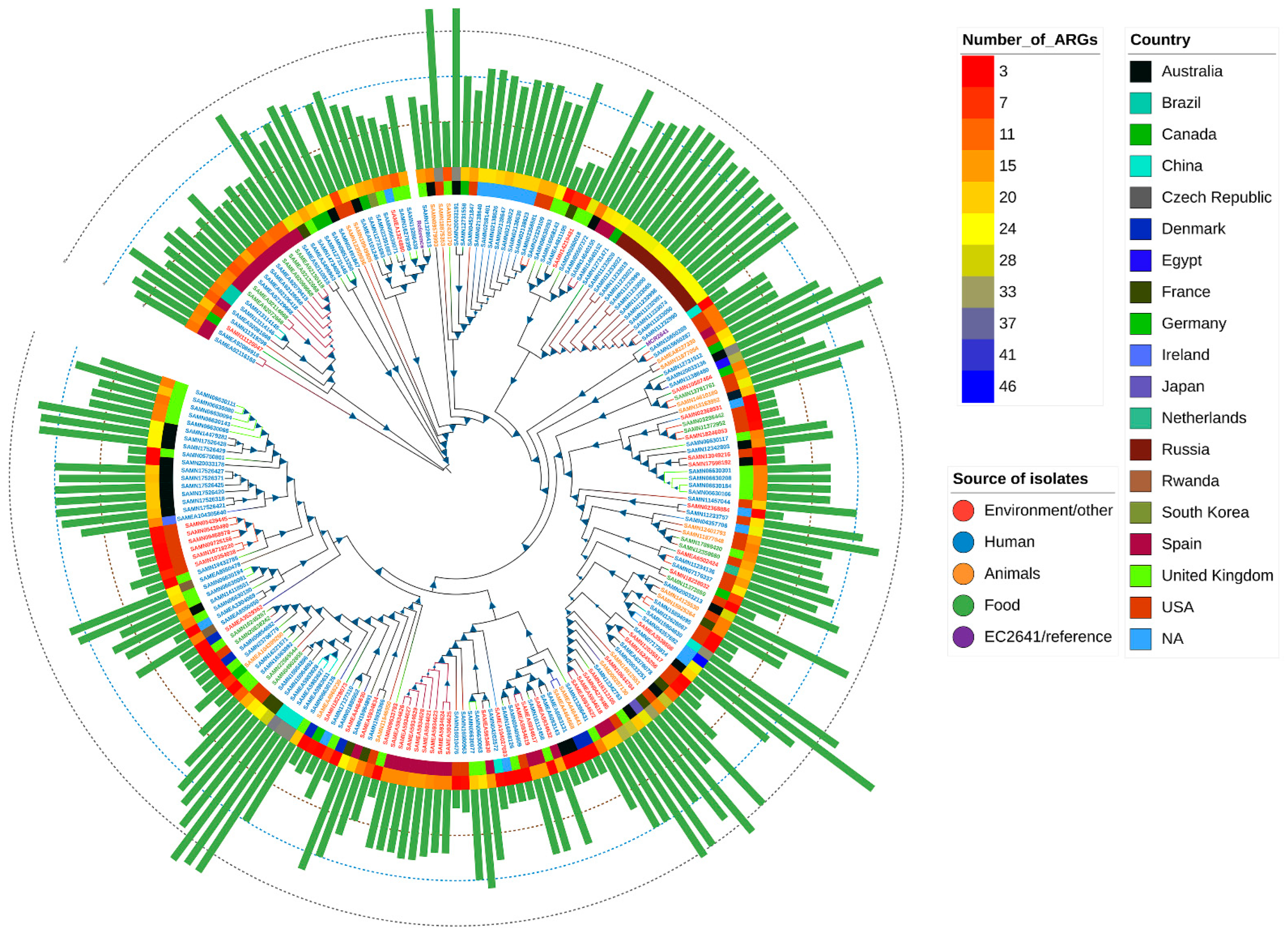
2.7. Global Phylogenetic Analysis of E. coli Carrying the mcr Variant
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolation and Species Identification
4.2. In Vitro Antimicrobial Susceptibility Testing
4.3. Transconjugation Assay
4.4. Induced Expression, RNA Extraction, Real-Time Reverse Transcription PCR (RT–PCR)
4.5. DNA Extraction, Whole-Genome Sequencing
4.6. Sequence Assembly, Annotation and Bioinformatic Analysis
4.7. Phylogenetic Analysis
4.8. Accession Numbers
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hora, P.I.; Pati, S.G.; McNamara, P.J.; Arnold, W.A. Increased Use of Quaternary Ammonium Compounds during the SARS-CoV-2 Pandemic and Beyond: Consideration of Environmental Implications. Environ. Sci. Technol. Lett. 2020, 7, 622–631. [Google Scholar] [CrossRef]
- Zheng, G.; Filippelli, G.M.; Salamova, A. Increased Indoor Exposure to Commonly Used Disinfectants during the COVID-19 Pandemic. Environ. Sci. Technol. Lett. 2020, 7, 760–765. [Google Scholar] [CrossRef]
- Butucel, E.; Balta, I.; Ahmadi, M.; Dumitrescu, G.; Morariu, F.; Pet, I.; Stef, L.; Corcionivoschi, N. Biocides as Biomedicines against Foodborne Pathogenic Bacteria. Biomedicines 2022, 10, 379. [Google Scholar] [CrossRef] [PubMed]
- Vereshchagin, A.N.; Frolov, N.A.; Egorova, K.S.; Seitkalieva, M.M.; Ananikov, V.P. Quaternary Ammonium Compounds (QACs) and Ionic Liquids (ILs) as Biocides: From Simple Antiseptics to Tunable Antimicrobials. Int. J. Mol. Sci. 2021, 22, 6793. [Google Scholar] [CrossRef]
- Marple, B.; Roland, P.; Benninger, M. Safety review of benzalkonium chloride used as a preservative in intranasal solutions: An overview of conflicting data and opinions. Otolaryngol. Head Neck Surg. 2004, 130, 131–141. [Google Scholar] [CrossRef]
- Mao, X.; Auer, D.L.; Buchalla, W.; Hiller, K.-A.; Maisch, T.; Hellwig, E.; Al-Ahmad, A.; Cieplik, F. Cetylpyridinium Chloride: Mechanism of Action, Antimicrobial Efficacy in Biofilms, and Potential Risks of Resistance. Antimicrob. Agents Chemother. 2020, 64, e00576-20. [Google Scholar] [CrossRef]
- Merchel Piovesan Pereira, B.; Tagkopoulos, I. Benzalkonium Chlorides: Uses, Regulatory Status, and Microbial Resistance. Appl. Environ. Microbiol. 2019, 85, e00377-19. [Google Scholar] [CrossRef]
- Buffet-Bataillon, S.; Branger, B.; Cormier, M.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Effect of higher minimum inhibitory concentrations of quaternary ammonium compounds in clinical E. coli isolates on antibiotic susceptibilities and clinical outcomes. J. Hosp. Infect. 2011, 79, 141–146. [Google Scholar] [CrossRef]
- Kim, M.; Weigand, M.R.; Oh, S.; Hatt, J.K.; Krishnan, R.; Tezel, U.; Pavlostathis, S.G.; Konstantinidis, K.T.; Dozois, C.M. Widely Used Benzalkonium Chloride Disinfectants Can Promote Antibiotic Resistance. Appl. Environ. Microbiol. 2018, 84, e01201-18. [Google Scholar] [CrossRef]
- Russell, A.D. Biocide use and antibiotic resistance: The relevance of laboratory findings to clinical and environmental situations. Lancet Infect. Dis. 2003, 3, 794–803. [Google Scholar] [CrossRef]
- Zhang, A.; He, X.; Meng, Y.; Guo, L.; Long, M.; Yu, H.; Li, B.; Fan, L.; Liu, S.; Wang, H.; et al. Antibiotic and Disinfectant Resistance of Escherichia coli Isolated from Retail Meats in Sichuan, China. Microb. Drug Resist. 2016, 22, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Brunel, J.M.; Dubus, J.C.; Reynaud-Gaubert, M.; Rolain, J.M. Colistin: An update on the antibiotic of the 21st century. Expert Rev. Anti. Infect. Ther. 2012, 10, 917–934. [Google Scholar] [CrossRef] [PubMed]
- Sheu, C.C.; Chang, Y.T.; Lin, S.Y.; Chen, Y.H.; Hsueh, P.R. Infections Caused by Carbapenem-Resistant Enterobacteriaceae: An Update on Therapeutic Options. Front. Microbiol. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Nation, R.L.; Turnidge, J.D.; Milne, R.W.; Coulthard, K.; Rayner, C.R.; Paterson, D.L. Colistin: The re-emerging antibiotic for multidrug-resistant Gram-negative bacterial infections. Lancet Infect. Dis. 2006, 6, 589–601. [Google Scholar] [CrossRef]
- Sun, H.; Wan, Y.; Du, P.; Liu, D.; Li, R.; Zhang, P.; Wu, Y.; Fanning, S.; Wang, Y.; Bai, L. Investigation of tigecycline resistant Escherichia coli from raw meat reveals potential transmission among food-producing animals. Food Control 2021, 121, 107633. [Google Scholar] [CrossRef]
- Tartor, Y.H.; Abd El-Aziz, N.K.; Gharieb, R.M.A.; El Damaty, H.M.; Enany, S.; Soliman, E.A.; Abdellatif, S.S.; Attia, A.S.A.; Bahnass, M.M.; El-Shazly, Y.A.; et al. Whole-Genome Sequencing of Gram-Negative Bacteria Isolated From Bovine Mastitis and Raw Milk: The First Emergence of Colistin mcr-10 and Fosfomycin fosA5 Resistance Genes in Klebsiella pneumoniae in Middle East. Front. Microbiol. 2021, 12, 770813. [Google Scholar] [CrossRef]
- Ling, Z.; Yin, W.; Shen, Z.; Wang, Y.; Shen, J.; Walsh, T.R. Epidemiology of mobile colistin resistance genes mcr-1 to mcr-9. J. Antimicrob. Chemother. 2020, 75, 3087–3095. [Google Scholar] [CrossRef]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of novel mobile colistin resistance gene mcr-10. Emerg. Microbes Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef]
- Xu, T.; Zhang, C.; Ji, Y.; Song, J.; Liu, Y.; Guo, Y.; Zhou, K. Identification of mcr-10 carried by self-transmissible plasmids and chromosome in Enterobacter roggenkampii strains isolated from hospital sewage water. Environ. Pollut. 2021, 268, 115706. [Google Scholar] [CrossRef]
- Edgar, R.; Bibi, E. MdfA, an Escherichia coli multidrug resistance protein with an extraordinarily broad spectrum of drug recognition. J. Bacteriol. 1997, 179, 2274–2280. [Google Scholar] [CrossRef]
- Kümmerle, N.; Feucht, H.H.; Kaulfers, P.M. Plasmid-mediated formaldehyde resistance in Escherichia coli: Characterization of resistance gene. Antimicrob. Agents Chemother. 1996, 40, 2276–2279. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, L.; Feng, Y.; He, D.; Wang, C.; Zong, Z. Potential Mobilization of mcr-10 by an Integrative Mobile Element via Site-Specific Recombination in Cronobacter sakazakii. Antimicrob. Agents Chemother. 2021, 65, e01717-20. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.W.; Zhang, Y.; Wang, Y.T.; Wang, H.N. Detection of Mobile Colistin Resistance Gene mcr-10.1 in a Conjugative Plasmid from Enterobacter roggenkampii of Chicken Origin in China. Antimicrob. Agents Chemother. 2020, 64, e01191-20. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Hua, X.; Xu, Q.; Yang, Y.; Zhang, L.; He, J.; Mu, X.; Hu, L.; Leptihn, S.; Yu, Y. Mechanism of eravacycline resistance in Acinetobacter baumannii mediated by a deletion mutation in the sensor kinase adeS, leading to elevated expression of the efflux pump AdeABC. Infect. Genet. Evol. 2020, 80, 104185. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Hua, X.; He, J.; Zhang, D.; Chen, Q.; Zhang, L.; Loh, B.; Leptihn, S.; Wen, Y.; Higgins, P.G.; et al. The distribution of mutations and hotspots in transcription regulators of resistance-nodulation-cell division efflux pumps in tigecycline non-susceptible Acinetobacter baumannii in China. Int. J. Med. Microbiol. 2020, 310, 151464. [Google Scholar] [CrossRef]
- Gerson, S.; Nowak, J.; Zander, E.; Ertel, J.; Wen, Y.; Krut, O.; Seifert, H.; Higgins, P.G. Diversity of mutations in regulatory genes of resistance-nodulation-cell division efflux pumps in association with tigecycline resistance in Acinetobacter baumannii. J. Antimicrob. Chemother. 2018, 73, 1501–1508. [Google Scholar] [CrossRef]
- Tong, C.; Hu, H.; Chen, G.; Li, Z.; Li, A.; Zhang, J. Disinfectant resistance in bacteria: Mechanisms, spread, and resolution strategies. Environ. Res. 2021, 195, 110897. [Google Scholar] [CrossRef]
- Zou, L.; Meng, J.; McDermott, P.F.; Wang, F.; Yang, Q.; Cao, G.; Hoffmann, M.; Zhao, S. Presence of disinfectant resistance genes in Escherichia coli isolated from retail meats in the USA. J. Antimicrob. Chemother. 2014, 69, 2644–2649. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Wilksch, J.J.; Yang, J.; Clements, A.; Gabbe, J.L.; Short, K.R.; Cao, H.; Cavaliere, R.; James, C.E.; Whitchurch, C.B.; Schembri, M.A.; et al. MrkH, a novel c-di-GMP-dependent transcriptional activator, controls Klebsiella pneumoniae biofilm formation by regulating type 3 fimbriae expression. PLoS Pathog. 2011, 7, e1002204. [Google Scholar] [CrossRef]
- Huang, Y.-J.; Liao, H.-W.; Wu, C.-C.; Peng, H.-L. MrkF is a component of type 3 fimbriae in Klebsiella pneumoniae. Res. Microbiol. 2009, 160, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, J.; Wang, M.; Liu, M.; Liu, G.; Qu, H.; Liu, J.; Deng, Z.; Sun, J.; Ou, H.Y.; et al. Mobilization of the nonconjugative virulence plasmid from hypervirulent Klebsiella pneumoniae. Genome Med. 2021, 13, 119. [Google Scholar] [CrossRef] [PubMed]
- Furuya, N.; Nisioka, T.; Komano, T. Nucleotide sequence and functions of the oriT operon in IncI1 plasmid R64. J. Bacteriol. 1991, 173, 2231–2237. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xiaomin, S.; Yiming, L.; Yuying, Y.; Zhangqi, S.; Yongning, W.; Shaolin, W. Global impact of mcr-1-positive Enterobacteriaceae bacteria on “one health”. Crit. Rev. Microbiol. 2020, 46, 565–577. [Google Scholar] [CrossRef]
- Yang, X.; Ye, L.; Chan, E.W.-C.; Zhang, R.; Chen, S. Characterization of an IncFIB/IncHI1B Plasmid Encoding Efflux Pump TMexCD1-TOprJ1 in a Clinical Tigecycline-and Carbapenem-Resistant Klebsiella pneumoniae Strain. Antimicrob. Agents Chemother. 2021, 65, e02340-20. [Google Scholar] [CrossRef]
- Lin, D.L.; Traglia, G.M.; Baker, R.; Sherratt, D.J.; Ramirez, M.S.; Tolmasky, M.E. Functional Analysis of the Acinetobacter baumannii XerC and XerD Site-Specific Recombinases: Potential Role in Dissemination of Resistance Genes. Antibiotics 2020, 9, 405. [Google Scholar] [CrossRef]
- Santos Costa, S.; Viveiros, M.; Rosato, A.E.; Melo-Cristino, J.; Couto, I. Impact of efflux in the development of multidrug resistance phenotypes in Staphylococcus aureus. BMC Microbiol. 2015, 15, 232. [Google Scholar] [CrossRef]
- LaBreck, P.T.; Bochi-Layec, A.C.; Stanbro, J.; Dabbah-Krancher, G.; Simons, M.P.; Merrell, D.S. Systematic Analysis of Efflux Pump-Mediated Antiseptic Resistance in Staphylococcus aureus Suggests a Need for Greater Antiseptic Stewardship. Msphere 2020, 5, e00959-19. [Google Scholar] [CrossRef]
- Knapp, L.; Rushton, L.; Stapleton, H.; Sass, A.; Stewart, S.; Amezquita, A.; McClure, P.; Mahenthiralingam, E.; Maillard, J.Y. The effect of cationic microbicide exposure against Burkholderia cepacia complex (Bcc); the use of Burkholderia lata strain 383 as a model bacterium. J. Appl. Microbiol. 2013, 115, 1117–1126. [Google Scholar] [CrossRef]
- Kampf, G. Biocidal Agents Used for Disinfection Can Enhance Antibiotic Resistance in Gram-Negative Species. Antibiotics 2018, 7, 110. [Google Scholar] [CrossRef]
- Short, F.L.; Lee, V.; Mamun, R.; Malmberg, R.; Li, L.; Espinosa, M.I.; Venkatesan, K.; Paulsen, I.T. Benzalkonium chloride antagonises aminoglycoside antibiotics and promotes evolution of resistance. EBioMedicine 2021, 73, 103653. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Wang, Y.; Xiao, Y. Prevalence and transmission of mobilized colistin resistance (mcr) gene in bacteria common to animals and humans. Biosaf. Health 2020, 2, 71–78. [Google Scholar] [CrossRef]
- Anyanwu, M.U.; Jaja, I.F.; Nwobi, O.C. Occurrence and Characteristics of Mobile Colistin Resistance (mcr) Gene-Containing Isolates from the Environment: A Review. Int. J. Environ. Res. Public Health 2020, 17, 1028. [Google Scholar] [CrossRef] [PubMed]
- Shridhar, P.B.; Worley, J.N.; Gao, X.; Yang, X.; Noll, L.W.; Shi, X.; Bai, J.; Meng, J.; Nagaraja, T.G. Analysis of virulence potential of Escherichia coli O145 isolated from cattle feces and hide samples based on whole genome sequencing. PLoS ONE 2019, 14, e0225057. [Google Scholar] [CrossRef]
- Fernández Márquez, M.L.; Burgos, M.J.; Pulido, R.P.; Gálvez, A.; López, R.L. Biocide Tolerance and Antibiotic Resistance in Salmonella Isolates from Hen Eggshells. Foodborne Pathog. Dis. 2017, 14, 89–95. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Thomsen, M.C.; Ahrenfeldt, J.; Cisneros, J.L.; Jurtz, V.; Larsen, M.V.; Hasman, H.; Aarestrup, F.M.; Lund, O. A Bacterial Analysis Platform: An Integrated System for Analysing Bacterial Whole Genome Sequencing Data for Clinical Diagnostics and Surveillance. PLoS ONE 2016, 11, e0157718. [Google Scholar] [CrossRef]
- Hsiao, W.; Wan, I.; Jones, S.J.; Brinkman, F.S. IslandPath: Aiding detection of genomic islands in prokaryotes. Bioinformatics 2003, 19, 418–420. [Google Scholar] [CrossRef]
- Li, X.; Xie, Y.; Liu, M.; Tai, C.; Sun, J.; Deng, Z.; Ou, H.Y. oriTfinder: A web-based tool for the identification of origin of transfers in DNA sequences of bacterial mobile genetic elements. Nucleic Acids Res. 2018, 46, W229–W234. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate ID | MIC (µg/mL) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| TGC | MEM | AMP | CST | PME | FOX | CRO | CHL | TET | BAC | CPC | |
| EC740 | 2 | 0.06 | ≥256 | 1 | 1 | 2 | <0.6 | 16 | 8 | 16 | 8 |
| EC799 | 2 | 0.03 | 32 | 2 | 1 | 2 | <0.6 | 32 | 32 | 16 | 8 |
| EC875 | 2 | 0.03 | 128 | 1 | 1 | 8 | <0.6 | 256 | 256 | 16 | 8 |
| EC2299 | 2 | 0.06 | ≥256 | 1 | 0.5 | 4 | ≥32 | 8 | 128 | 16 | 8 |
| EC2639 | 2 | 0.03 | 256 | 8 | 4 | 4 | <0.6 | 8 | 16 | 16 | 8 |
| EC2641 | 2 | 0.12 | 32 | 4 | 4 | 4 | <0.6 | 16 | 16 | 16 | 8 |
| EC2783 | 2 | 0.06 | ≥256 | 0.5 | 0.5 | 4 | ≥32 | 16 | 128 | 16 | 8 |
| SY3705 | 2 | 0.03 | 32 | 2 | 2 | 16 | <0.6 | 8 | 4 | 16 | 8 |
| Isolate | Plasmid Replicons | Serotype | STs | Antimicrobial Resistance Profile | Resistance Determinants Identified Based on WGS |
|---|---|---|---|---|---|
| EC740 | Col156 | O104: H27 | ST2795 | TGC-AMP-BAC-CPC | mdf (A), blaZEG-1, formA |
| EC799 | Col440I, IncFIA (HI1), IncFIB (K), IncY | O81: H9 | ST1571 | TGC-AMP-CHL-TET-BAC-CPC | mdf (A), blaZEG-1, formA |
| EC875 | IncFIB (K) | OND: H16 | ST4537 | TGC-AMP-CHL-TET-BAC-CPC | aadA1, mdf (A), blaOXA-10, qnrS1, dfrA14, cmlA1, floR, ARR-2, tet (A), formA, blaMIR-2 |
| EC2299 | IncY | O159: H34 | ST218 | TGC-AMP-CRO-TET-BAC-CPC | mdf (A), blaZEG-1, formA |
| EC2639 | Col440I, Col156, IncR | O155: H27 | ST2795 | TGC-AMP-CST-PME-TET-BAC-CPC | mdf (A), blaZEG-1, formA |
| EC2641 | Col440I, FIA (pBK30683), IncFIB (K), IncHI1A, IncHI1B (R27) | O112ab: H9 | ST3907 | TGC-AMP-CST-PME-TET-BAC-CPC | mdf (A), mcr-10, blaZEG-1, formA |
| EC2783 | IncFIA, IncFIB (AP001918), IncFIC (FII), IncI1-I (Alpha), IncX9 | O128ac: H34 | ST3076 | TGC-AMP-CST-PME-TET-BAC-CPC | blaCTX-M-55, blaTEM-1B, qnrS1, tet (A), sitABCD, strB, dfrA14, mdf (A), sul2 |
| SY3705 | IncFIB (K), IncFII, IncY | O83: H19 | ST5783 | TGC-AMP-BAC-CPC | mdf (A), blaZEG-1, formA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Sun, H.; Lao, G.; Zhou, Z.; Liu, Z.; Cai, J.; Sun, Q. Identification of Mobile Colistin Resistance Gene mcr-10 in Disinfectant and Antibiotic Resistant Escherichia coli from Disinfected Tableware. Antibiotics 2022, 11, 883. https://doi.org/10.3390/antibiotics11070883
Zhang S, Sun H, Lao G, Zhou Z, Liu Z, Cai J, Sun Q. Identification of Mobile Colistin Resistance Gene mcr-10 in Disinfectant and Antibiotic Resistant Escherichia coli from Disinfected Tableware. Antibiotics. 2022; 11(7):883. https://doi.org/10.3390/antibiotics11070883
Chicago/Turabian StyleZhang, Senlin, Honghu Sun, Guangjie Lao, Zhiwei Zhou, Zhuochong Liu, Jiong Cai, and Qun Sun. 2022. "Identification of Mobile Colistin Resistance Gene mcr-10 in Disinfectant and Antibiotic Resistant Escherichia coli from Disinfected Tableware" Antibiotics 11, no. 7: 883. https://doi.org/10.3390/antibiotics11070883
APA StyleZhang, S., Sun, H., Lao, G., Zhou, Z., Liu, Z., Cai, J., & Sun, Q. (2022). Identification of Mobile Colistin Resistance Gene mcr-10 in Disinfectant and Antibiotic Resistant Escherichia coli from Disinfected Tableware. Antibiotics, 11(7), 883. https://doi.org/10.3390/antibiotics11070883