
Strategies to Combat Caries by Maintaining the Integrity of Biofilm and Homeostasis during the Rapid Phase of Supragingival Plaque Formation

 ,
,  ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. From Homeostatic to Dysbiotic Supragingival Biofilm
3. Benefits Derived from Biofilms in Homeostasis and the Disadvantages of Dysbiosis
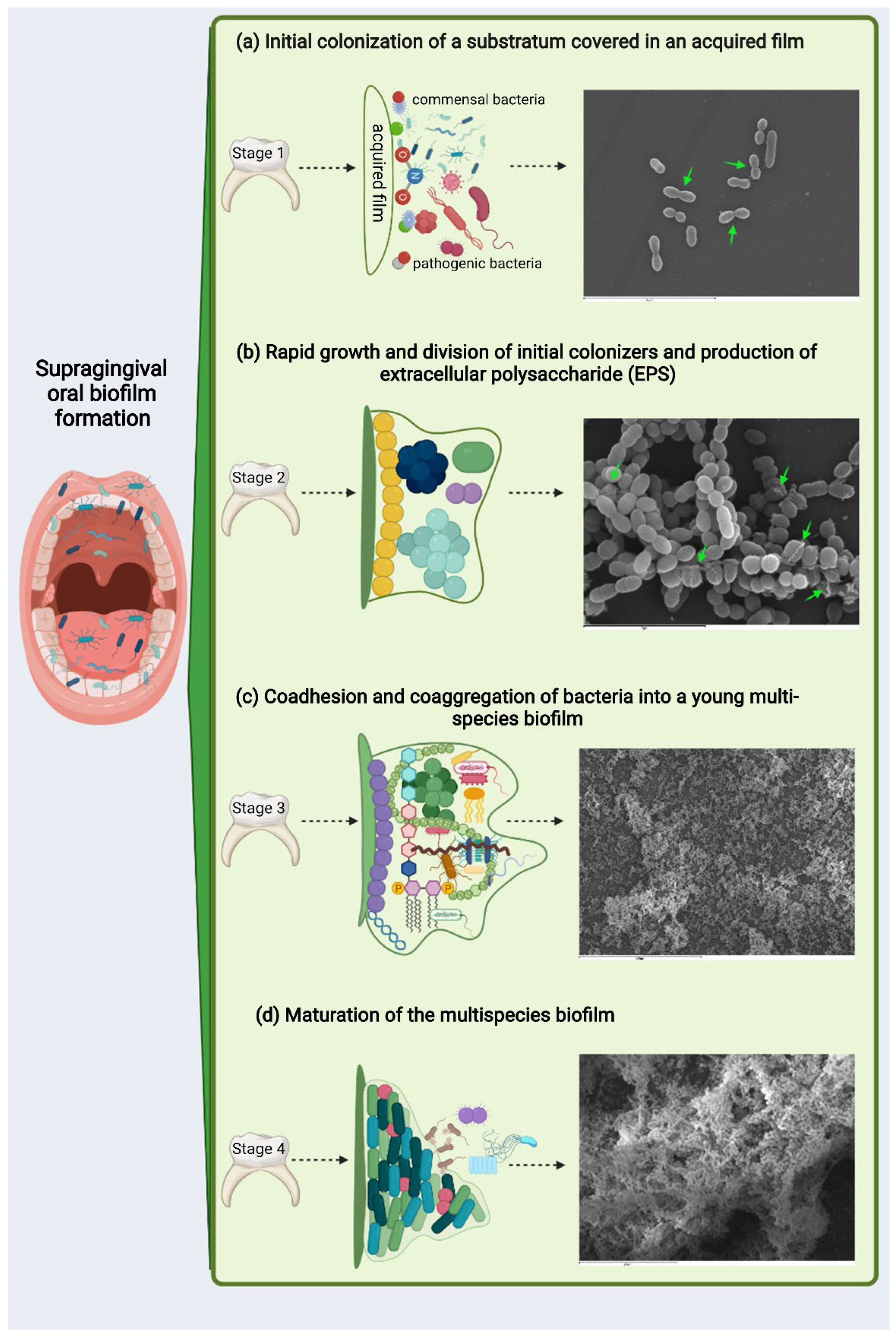
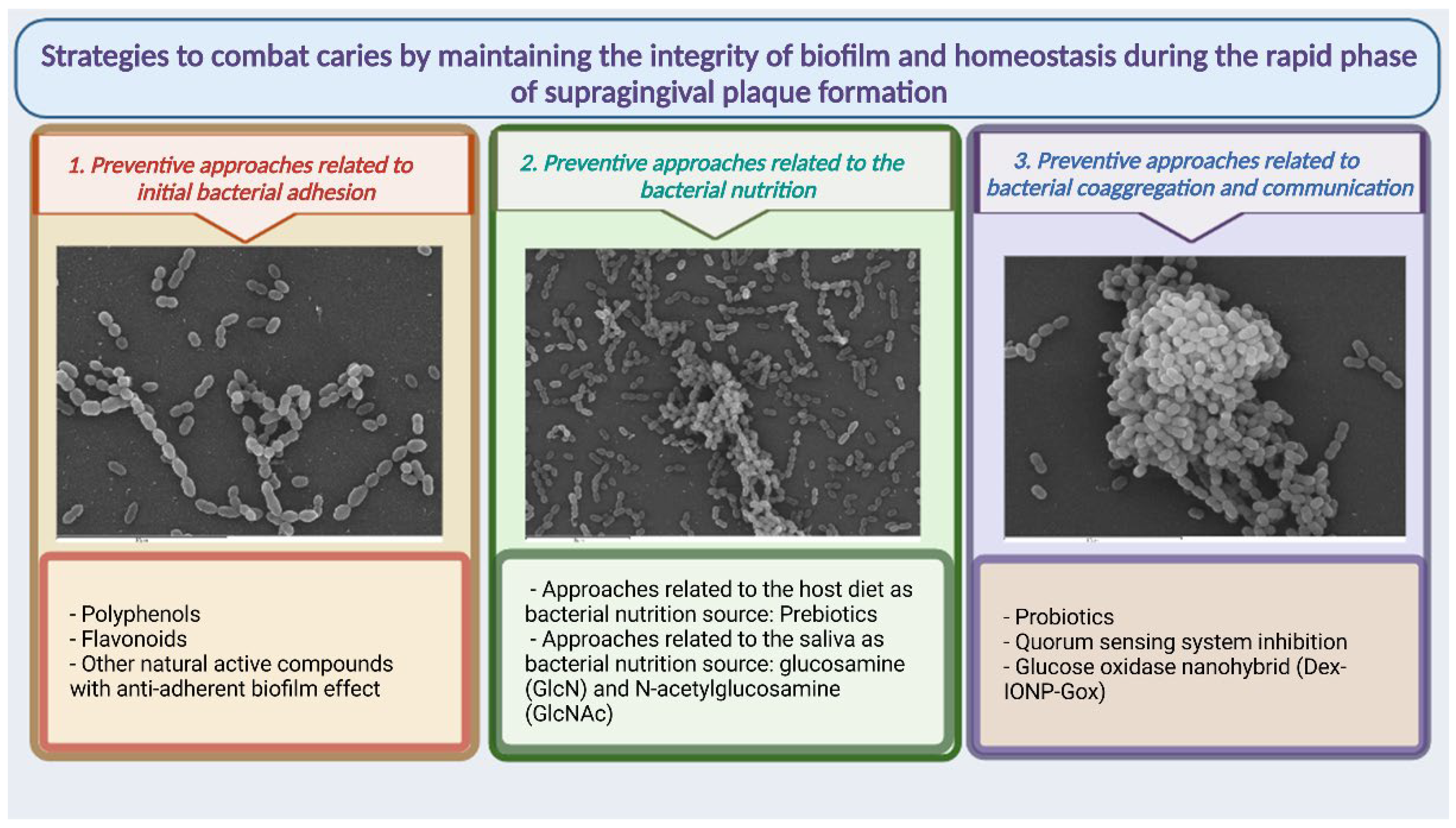
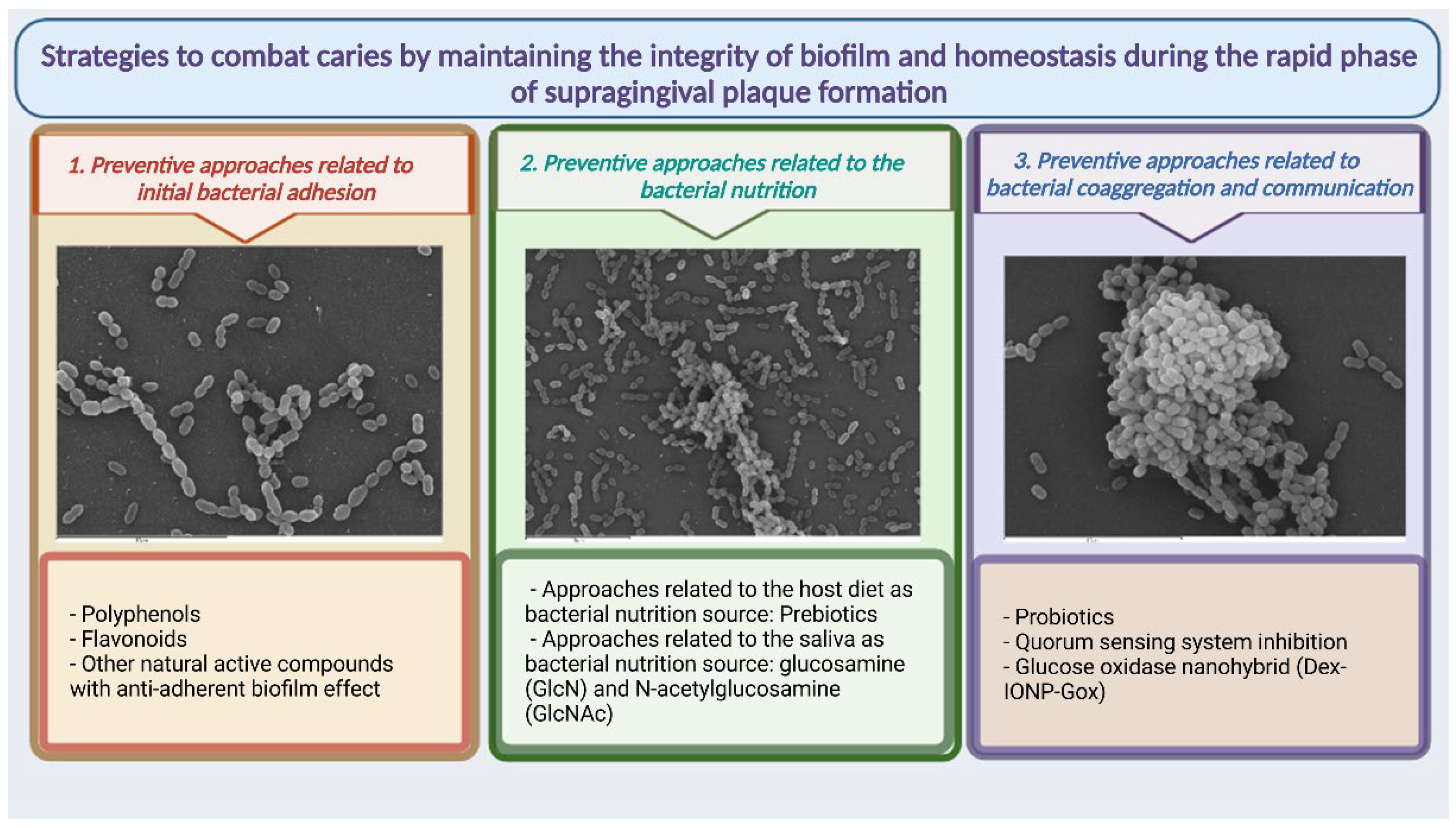
4. Strategies to Combat Caries by Maintaining the Integrity of Biofilm and Homeostasis during the Rapid Phase of Supragingival Plaque Formation
4.1. Preventive Approaches Related to Initial Bacterial Adhesion
4.1.1. Polyphenols
4.1.2. Flavonoids
4.1.3. Other Natural Active Compounds with Anti-Adhesion Biofilm Effect
4.2. Preventive Approaches Related to Bacterial Nutrition
4.2.1. Preventive Approaches Related to Host Diet as Bacterial Nutrition Source
4.2.2. Preventive Approaches Related to Saliva as a Bacterial Nutrition Source
4.3. Preventive Approaches Related to Bacterial Coaggregation and Communication
4.3.1. Probiotics and Supragingival Homeostasis
4.3.2. QS System Inhibition
4.3.3. Glucose Oxidase Nanohybrid (Dex-IONP-Gox)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Global Burden of Disease Collaborative Network. Global Burden of Disease Study; Institute of Health Metrics and Evaluation (IHME): Seattle, WA, USA, 2019. [Google Scholar]
- Marsh, P.; Head, D.A.; Devine, D.A. Dental plaque as a biofilm and a microbial community-implications for treatment. J. Oral Biosci. 2015, 57, 7. [Google Scholar] [CrossRef]
- Lif Holgerson, P.; Ohman, C.; Ronnlund, A.; Johansson, I. Maturation of Oral Microbiota in Children with or without Dental Caries. PLoS ONE 2015, 10, e0128534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, P.D.; Moter, A.; Devine, D.A. Dental plaque biofilms: Communities, conflict and control. Periodontol 2000 2011, 55, 16–35. [Google Scholar] [CrossRef] [PubMed]
- Marsh, P.D. Dental plaque as a microbial biofilm. Caries Res. 2004, 38, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Aas, J.A.; Griffen, A.L.; Dardis, S.R.; Lee, A.M.; Olsen, I.; Dewhirst, F.E.; Leys, E.J.; Paster, B.J. Bacteria of dental caries in primary and permanent teeth in children and young adults. J. Clin. Microbiol. 2008, 46, 1407–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksson, L.; Lif Holgerson, P.; Esberg, A.; Johansson, I. Microbial Complexes and Caries in 17-Year-Olds with and without Streptococcus mutans. J. Dent. Res. 2018, 97, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Zijnge, V.; van Leeuwen, M.B.; Degener, J.E.; Abbas, F.; Thurnheer, T.; Gmur, R.; Harmsen, H.J. Oral biofilm architecture on natural teeth. PLoS ONE 2010, 5, e9321. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, M.C.; Velapatino, A.; Llama-Palacios, A.; Valdes, A.; Cifuentes, A.; Ciudad, M.J.; Collado, L. Metataxonomic and metabolomic evidence of biofilm homeostasis disruption related to caries: An in vitro study. Mol. Oral Microbiol. 2022, 37, 81–96. [Google Scholar] [CrossRef]
- Johansson, I.; Witkowska, E.; Kaveh, B.; Lif Holgerson, P.; Tanner, A.C. The Microbiome in Populations with a Low and High Prevalence of Caries. J. Dent. Res. 2016, 95, 80–86. [Google Scholar] [CrossRef]
- Mason, M.R.; Chambers, S.; Dabdoub, S.M.; Thikkurissy, S.; Kumar, P.S. Characterizing oral microbial communities across dentition states and colonization niches. Microbiome 2018, 6, 67. [Google Scholar] [CrossRef]
- Mira, A.; Simon-Soro, A.; Curtis, M.A. Role of microbial communities in the pathogenesis of periodontal diseases and caries. J. Clin. Periodontol. 2017, 44 (Suppl. 18), S23–S38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munson, M.A.; Banerjee, A.; Watson, T.F.; Wade, W.G. Molecular analysis of the microflora associated with dental caries. J. Clin. Microbiol. 2004, 42, 3023–3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanner, A.C.; Kent, R.L., Jr.; Holgerson, P.L.; Hughes, C.V.; Loo, C.Y.; Kanasi, E.; Chalmers, N.I.; Johansson, I. Microbiota of severe early childhood caries before and after therapy. J. Dent. Res. 2011, 90, 1298–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wake, N.; Asahi, Y.; Noiri, Y.; Hayashi, M.; Motooka, D.; Nakamura, S.; Gotoh, K.; Miura, J.; Machi, H.; Iida, T.; et al. Temporal dynamics of bacterial microbiota in the human oral cavity determined using an in situ model of dental biofilms. NPJ Biofilms Microbiomes 2016, 2, 16018. [Google Scholar] [CrossRef] [PubMed]
- Ximenes, M.; Armas, R.D.; Triches, T.C.; Cardoso, M.; Vieira, R.S. Structure of the Bacterial Community in Different Stages of Early Childhood Caries. Oral Health Prev. Dent. 2018, 16, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Haffajee, A.D.; Teles, R.P.; Patel, M.R.; Song, X.; Veiga, N.; Socransky, S.S. Factors affecting human supragingival biofilm composition. I. Plaque mass. J. Periodontal Res. 2009, 44, 511–519. [Google Scholar] [CrossRef]
- Rath, S.; Bal, S.C.B.; Dubey, D. Oral Biofilm: Development Mechanism, Multidrug Resistance, and Their Effective Management with Novel Techniques. Rambam Maimonides Med. J. 2021, 12, e0004. [Google Scholar] [CrossRef]
- Featherstone, J.D. The continuum of dental caries—Evidence for a dynamic disease process. J. Dent. Res. 2004, 83, C39–C42. [Google Scholar] [CrossRef]
- Radaic, A.; Kapila, Y.L. The oralome and its dysbiosis: New insights into oral microbiome-host interactions. Comput. Struct. Biotechnol. J. 2021, 19, 1335–1360. [Google Scholar] [CrossRef]
- Takahashi, N.; Nyvad, B. The role of bacteria in the caries process: Ecological perspectives. J. Dent. Res. 2011, 90, 294–303. [Google Scholar] [CrossRef]
- Fejerskov, O. Changing paradigms in concepts on dental caries: Consequences for oral health care. Caries Res. 2004, 38, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Kidd, E.A.; Fejerskov, O. What constitutes dental caries? Histopathology of carious enamel and dentin related to the action of cariogenic biofilms. J. Dent. Res. 2004, 83, C35–C38. [Google Scholar] [CrossRef] [PubMed]
- Selwitz, R.H.; Ismail, A.I.; Pitts, N.B. Dental caries. Lancet 2007, 369, 51–59. [Google Scholar] [CrossRef]
- Schmidt, J.C.; Zaugg, C.; Weiger, R.; Walter, C. Brushing without brushing?—A review of the efficacy of powered toothbrushes in noncontact biofilm removal. Clin. Oral Investig. 2013, 17, 687–709. [Google Scholar] [CrossRef]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef]
- Sedghi, L.; DiMassa, V.; Harrington, A.; Lynch, S.V.; Kapila, Y.L. The oral microbiome: Role of key organisms and complex networks in oral health and disease. Periodontol 2000 2021, 87, 107–131. [Google Scholar] [CrossRef]
- Kolenbrander, P.E.; Andersen, R.N.; Blehert, D.S.; Egland, P.G.; Foster, J.S.; Palmer, R.J., Jr. Communication among oral bacteria. Microbiol. Mol. Biol. Rev. 2002, 66, 486–505. [Google Scholar] [CrossRef] [Green Version]
- Palmer, R.J., Jr.; Gordon, S.M.; Cisar, J.O.; Kolenbrander, P.E. Coaggregation-mediated interactions of streptococci and actinomyces detected in initial human dental plaque. J. Bacteriol. 2003, 185, 3400–3409. [Google Scholar] [CrossRef] [Green Version]
- Kolenbrander, P.E.; Palmer, R.J., Jr.; Periasamy, S.; Jakubovics, N.S. Oral multispecies biofilm development and the key role of cell-cell distance. Nat. Rev. Microbiol. 2010, 8, 471–480. [Google Scholar] [CrossRef]
- Sun, Z.; Xi, J.; Yang, C.; Cong, W. Quorum sensing regulation methods and their effects on biofilm in biological waste treatment systems: A review. Front. Environ. Sci. Eng. 2022, 16, 87. [Google Scholar] [CrossRef]
- Kolenbrander, P.E.; Palmer, R.J., Jr.; Rickard, A.H.; Jakubovics, N.S.; Chalmers, N.I.; Diaz, P.I. Bacterial interactions and successions during plaque development. Periodontol 2000 2006, 42, 47–79. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Li, M.; Gregory, R.L. Bacterial interactions in dental biofilm. Virulence 2011, 2, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Diaz, P.I.; Valm, A.M. Microbial Interactions in Oral Communities Mediate Emergent Biofilm Properties. J. Dent. Res. 2020, 99, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Mark Welch, J.L.; Rossetti, B.J.; Rieken, C.W.; Dewhirst, F.E.; Borisy, G.G. Biogeography of a human oral microbiome at the micron scale. Proc. Natl. Acad. Sci. USA 2016, 113, E791–E800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreth, J.; Zhang, Y.; Herzberg, M.C. Streptococcal antagonism in oral biofilms: Streptococcus sanguinis and Streptococcus gordonii interference with Streptococcus mutans. J. Bacteriol. 2008, 190, 4632–4640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreth, J.; Merritt, J.; Qi, F. Bacterial and host interactions of oral streptococci. DNA Cell Biol. 2009, 28, 397–403. [Google Scholar] [CrossRef]
- Marsh, P.D.; Zaura, E. Dental biofilm: Ecological interactions in health and disease. J. Clin. Periodontol. 2017, 44 (Suppl. 18), S12–S22. [Google Scholar] [CrossRef]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef]
- Chen, X.; Daliri, E.B.; Tyagi, A.; Oh, D.H. Cariogenic Biofilm: Pathology-Related Phenotypes and Targeted Therapy. Microorganisms 2021, 9, 1311. [Google Scholar] [CrossRef]
- Jakubovics, N.S.; Yassin, S.A.; Rickard, A.H. Community interactions of oral streptococci. Adv. Appl. Microbiol. 2014, 87, 43–110. [Google Scholar] [CrossRef]
- Ho, M.H.; Lamont, R.J.; Xie, H. A novel peptidic inhibitor derived from Streptococcus cristatus ArcA attenuates virulence potential of Porphyromonas gingivalis. Sci. Rep. 2017, 7, 16217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, C.J.; Burns, L.H.; Jack, A.A.; Back, C.R.; Dutton, L.C.; Nobbs, A.H.; Lamont, R.J.; Jenkinson, H.F. Microbial interactions in building of communities. Mol. Oral Microbiol. 2013, 28, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xie, H. Role of arginine deiminase of Streptococcus cristatus in Porphyromonas gingivalis colonization. Antimicrob. Agents Chemother. 2010, 54, 4694–4698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Kreth, J. The role of hydrogen peroxide in environmental adaptation of oral microbial communities. Oxid. Med. Cell Longev. 2012, 2012, 717843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubovics, N.S.; Gill, S.R.; Vickerman, M.M.; Kolenbrander, P.E. Role of hydrogen peroxide in competition and cooperation between Streptococcus gordonii and Actinomyces naeslundii. FEMS Microbiol. Ecol. 2008, 66, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Perera, D.; McLean, A.; Morillo-Lopez, V.; Cloutier-Leblanc, K.; Almeida, E.; Cabana, K.; Mark Welch, J.; Ramsey, M. Mechanisms underlying interactions between two abundant oral commensal bacteria. ISME J. 2022, 16, 948–957. [Google Scholar] [CrossRef]
- Huang, X.; Browngardt, C.M.; Jiang, M.; Ahn, S.J.; Burne, R.A.; Nascimento, M.M. Diversity in Antagonistic Interactions between Commensal Oral Streptococci and Streptococcus mutans. Caries Res. 2018, 52, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Bondonno, C.P.; Liu, A.H.; Croft, K.D.; Considine, M.J.; Puddey, I.B.; Woodman, R.J.; Hodgson, J.M. Antibacterial mouthwash blunts oral nitrate reduction and increases blood pressure in treated hypertensive men and women. Am. J. Hypertens. 2015, 28, 572–575. [Google Scholar] [CrossRef] [Green Version]
- Kapil, V.; Haydar, S.M.; Pearl, V.; Lundberg, J.O.; Weitzberg, E.; Ahluwalia, A. Physiological role for nitrate-reducing oral bacteria in blood pressure control. Free Radic. Biol. Med. 2013, 55, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Buskermolen, J.K.; Janus, M.M.; Roffel, S.; Krom, B.P.; Gibbs, S. Saliva-Derived Commensal and Pathogenic Biofilms in a Human Gingiva Model. J. Dent. Res. 2018, 97, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Vyhnalova, T.; Danek, Z.; Gachova, D.; Linhartova, P.B. The Role of the Oral Microbiota in the Etiopathogenesis of Oral Squamous Cell Carcinoma. Microorganisms 2021, 9, 1549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Chen, R.; Rudney, J.D. Streptococcus cristatus attenuates Fusobacterium nucleatum-induced interleukin-8 expression in oral epithelial cells. J. Periodontal Res. 2008, 43, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.S.; Goudarzi, H.; Ghalavand, Z.; Hajikhani, B.; Rafeieiatani, Z.; Hakemi-Vala, M. Anti-proliferative effects of cell wall, cytoplasmic extract of Lactococcus lactis and nisin through down-regulation of cyclin D1 on SW480 colorectal cancer cell line. Iran J. Microbiol. 2020, 12, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Ghollasi, M.; Hosseini, H.M. The apoptotic impact of nisin as a potent bacteriocin on the colon cancer cells. Microb. Pathog. 2017, 111, 193–197. [Google Scholar] [CrossRef]
- Radaic, A.; Ye, C.; Parks, B.; Gao, L.; Kuraji, R.; Malone, E.; Kamarajan, P.; Zhan, L.; Kapila, Y.L. Modulation of pathogenic oral biofilms towards health with nisin probiotic. J. Oral Microbiol. 2020, 12, 1809302. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Dong, L.; Zhou, L.; Tao, R.; Ni, L. Nisin inhibits dental caries-associated microorganism in vitro. Peptides 2010, 31, 2003–2008. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Ni, L.; Ling, J. Antibacterial peptide nisin: A potential role in the inhibition of oral pathogenic bacteria. Peptides 2014, 60, 32–40. [Google Scholar] [CrossRef]
- Sanz, M.; Beighton, D.; Curtis, M.A.; Cury, J.A.; Dige, I.; Dommisch, H.; Ellwood, R.; Giacaman, R.A.; Herrera, D.; Herzberg, M.C.; et al. Role of microbial biofilms in the maintenance of oral health and in the development of dental caries and periodontal diseases. Consensus report of group 1 of the Joint EFP/ORCA workshop on the boundaries between caries and periodontal disease. J. Clin. Periodontol. 2017, 44 (Suppl. 18), S5–S11. [Google Scholar] [CrossRef]
- Yoo, S.-M.; Ahn, G.-S. Correlation of Oral Microorganism and Carboxylic Acid in Oral Cavity. Int. J. Clin. Prev. Dent. 2015, 11, 6. [Google Scholar] [CrossRef]
- Cugini, C.; Ramasubbu, N.; Tsiagbe, V.K.; Fine, D.H. Dysbiosis From a Microbial and Host Perspective Relative to Oral Health and Disease. Front. Microbiol. 2021, 12, 617485. [Google Scholar] [CrossRef]
- Takenaka, S.; Ohsumi, T.; Noiri, Y. Evidence-based strategy for dental biofilms: Current evidence of mouthwashes on dental biofilm and gingivitis. Jpn Dent. Sci. Rev. 2019, 55, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Socransky, S.S.; Manganiello, A.D.; Propas, D.; Oram, V.; van Houte, J. Bacteriological studies of developing supragingival dental plaque. J. Periodontal Res. 1977, 12, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Nyvad, B.; Kilian, M. Microbiology of the early colonization of human enamel and root surfaces in vivo. Scand. J. Dent. Res. 1987, 95, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Moran, J.; Addy, M.; Newcombe, R.; Warren, P. The comparative effects on plaque regrowth of phenolic chlorhexidine and anti-adhesive mouthrinses. J. Clin. Periodontol. 1995, 22, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Abiko, Y.; Washio, J.; Luo, Y.; Zhang, L.; Takahashi, N. Green Tea-Derived Epigallocatechin Gallate Inhibits Acid Production and Promotes the Aggregation of Streptococcus mutans and Non-Mutans Streptococci. Caries Res. 2021, 55, 205–214. [Google Scholar] [CrossRef]
- Wu, C.D.; Huang, E.; Li, W.; White, M.; Jung, S.; Xie, Q. Beverages Containing Plant-Derived Polyphenols Inhibit Growth and Biofilm Formation of Streptococcus mutans and Children’s Supragingival Plaque Bacteria. Beverages 2021, 7, 43. [Google Scholar] [CrossRef]
- Palombo, E.A. Traditional Medicinal Plant Extracts and Natural Products with Activity against Oral Bacteria: Potential Application in the Prevention and Treatment of Oral Diseases. Evid. Based Complement. Alternat. Med. 2011, 2011, 680354. [Google Scholar] [CrossRef] [Green Version]
- Souissi, M.; Ben Lagha, A.; Chaieb, K.; Grenier, D. Effect of a Berry Polyphenolic Fraction on Biofilm Formation, Adherence Properties and Gene Expression of Streptococcus mutans and Its Biocompatibility with Oral Epithelial Cells. Antibiotics 2021, 10, 46. [Google Scholar] [CrossRef]
- Yamanaka, A.; Kimizuka, R.; Kato, T.; Okuda, K. Inhibitory effects of cranberry juice on attachment of oral streptococci and biofilm formation. Oral. Microbiol. Immunol. 2004, 19, 150–154. [Google Scholar] [CrossRef]
- Rahim, Z.H.; Khan, H.B. Comparative studies on the effect of crude aqueous (CA) and solvent (CM) extracts of clove on the cariogenic properties of Streptococcus mutans. J. Oral Sci. 2006, 48, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Karygianni, L.; Cecere, M.; Argyropoulou, A.; Hellwig, E.; Skaltsounis, A.L.; Wittmer, A.; Tchorz, J.P.; Al-Ahmad, A. Compounds from Olea europaea and Pistacia lentiscus inhibit oral microbial growth. BMC Complement. Altern. Med. 2019, 19, 51. [Google Scholar] [CrossRef] [PubMed]
- Philip, N.; Bandara, H.; Leishman, S.J.; Walsh, L.J. Inhibitory effects of fruit berry extracts on Streptococcus mutans biofilms. Eur. J. Oral. Sci. 2019, 127, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, A.; Sarveswari, H.B.; Vasudevan, S.; Shanmugam, K.; Solomon, A.P.; Neelakantan, P. Baicalein Inhibits Streptococcus mutans Biofilms and Dental Caries-Related Virulence Phenotypes. Antibiotics 2021, 10, 215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, Y.; Lu, M.; Lyu, X.; Gong, T.; Tang, B.; Wang, L.; Zeng, J.; Li, Y. Rhodiola rosea extract inhibits the biofilm formation and the expression of virulence genes of cariogenic oral pathogen Streptococcus mutans. Arch. Oral Biol. 2020, 116, 104762. [Google Scholar] [CrossRef]
- Shafiei, Z.; Rahim, Z.H.A.; Philip, K.; Thurairajah, N.; Yaacob, H. Potential effects of Psidium sp., Mangifera sp., Mentha sp. and its mixture (PEM) in reducing bacterial populations in biofilms, adherence and acid production of S. sanguinis and S. mutans. Arch. Oral Biol. 2020, 109, 104554. [Google Scholar] [CrossRef] [PubMed]
- Shafiei, Z.; Haji Abdul Rahim, Z.; Philip, K.; Thurairajah, N. Antibacterial and anti-adherence effects of a plant extract mixture (PEM) and its individual constituent extracts (Psidium sp., Mangifera sp., and Mentha sp.) on single- and dual-species biofilms. PeerJ 2016, 4, e2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Kim, S.C.; Kim, J.; Do, A.; Han, S.Y.; Lee, B.D.; Lee, H.H.; Lee, M.C.; Lee, S.H.; Oh, T.; et al. Synergistic inhibition of Streptococcal biofilm by ribose and xylitol. Arch. Oral Biol. 2015, 60, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Taheri, J.B.; Azimi, S.; Rafieian, N.; Zanjani, H.A. Herbs in dentistry. Int. Dent. J. 2011, 61, 287–296. [Google Scholar] [CrossRef]
- Nascimento, M.M.; Alvarez, A.J.; Huang, X.; Browngardt, C.; Jenkins, R.; Sinhoreti, M.C.; Ribeiro, A.P.D.; Dilbone, D.A.; Richards, V.P.; Garrett, T.J.; et al. Metabolic Profile of Supragingival Plaque Exposed to Arginine and Fluoride. J. Dent. Res. 2019, 98, 1245–1252. [Google Scholar] [CrossRef]
- Skinner, J.; Byun, R.; Blinkhorn, A.; Johnson, G. Sugary drink consumption and dental caries in New South Wales teenagers. Aust. Dent. J. 2015, 60, 169–175. [Google Scholar] [CrossRef]
- Tahmassebi, J.F.; Duggal, M.S.; Malik-Kotru, G.; Curzon, M.E. Soft drinks and dental health: A review of the current literature. J. Dent. 2006, 34, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chakraborty, B.; Zou, J.; Burne, R.A.; Zeng, L. Amino Sugars Modify Antagonistic Interactions between Commensal Oral Streptococci and Streptococcus mutans. Appl. Environ. Microbiol. 2019, 85, e00370-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moye, Z.D.; Burne, R.A.; Zeng, L. Uptake and metabolism of N-acetylglucosamine and glucosamine by Streptococcus mutans. Appl. Environ. Microbiol. 2014, 80, 5053–5067. [Google Scholar] [CrossRef] [Green Version]
- Roberfroid, M. Prebiotics: The concept revisited. J. Nutr. 2007, 137, 830S–837S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velmurugan, S.; Gan, J.M.; Rathod, K.S.; Khambata, R.S.; Ghosh, S.M.; Hartley, A.; Van Eijl, S.; Sagi-Kiss, V.; Chowdhury, T.A.; Curtis, M.; et al. Dietary nitrate improves vascular function in patients with hypercholesterolemia: A randomized, double-blind, placebo-controlled study. Am. J. Clin. Nutr. 2016, 103, 25–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jockel-Schneider, Y.; Schlagenhauf, U.; Stolzel, P.; Gossner, S.; Carle, R.; Ehmke, B.; Prior, K.; Hagenfeld, D. Nitrate-rich diet alters the composition of the oral microbiota in periodontal recall patients. J. Periodontol. 2021, 92, 1536–1545. [Google Scholar] [CrossRef] [PubMed]
- Kolderman, E.; Bettampadi, D.; Samarian, D.; Dowd, S.E.; Foxman, B.; Jakubovics, N.S.; Rickard, A.H. L-arginine destabilizes oral multi-species biofilm communities developed in human saliva. PLoS ONE 2015, 10, e0121835. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhang, K.; Deng, M.; Exterkate, R.A.M.; Liu, C.; Zhou, X.; Cheng, L.; Ten Cate, J.M. Effect of arginine on the growth and biofilm formation of oral bacteria. Arch. Oral Biol. 2017, 82, 256–262. [Google Scholar] [CrossRef]
- Huang, X.; Schulte, R.M.; Burne, R.A.; Nascimento, M.M. Characterization of the arginolytic microflora provides insights into pH homeostasis in human oral biofilms. Caries Res. 2015, 49, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Velsko, I.M.; Chakraborty, B.; Nascimento, M.M.; Burne, R.A.; Richards, V.P. Species Designations Belie Phenotypic and Genotypic Heterogeneity in Oral Streptococci. mSystems 2018, 3, e00158-18. [Google Scholar] [CrossRef] [Green Version]
- Agnello, M.; Cen, L.; Tran, N.C.; Shi, W.; McLean, J.S.; He, X. Arginine Improves pH Homeostasis via Metabolism and Microbiome Modulation. J. Dent. Res. 2017, 96, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Kamaguch, A.; Nakayama, K.; Ohyama, T.; Watanabe, T.; Okamoto, M.; Baba, H. Coaggregation of Porphyromonas gingivalis and Prevotella intermedia. Microbiol. Immunol. 2001, 45, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Tester, R.F.; Al-Ghazzewi, F.H. A preliminary study of the synbiotic effects of konjac glucomannan hydrolysates (GMH) and lactobacilli on the growth of the oral bacterium Streptococcus mutans. Nutr. Food Sci. 2011, 41, 5. [Google Scholar] [CrossRef]
- Bijle, M.N.; Ekambaram, M.; Lo, E.C.M.; Yiu, C.K.Y. Synbiotics in caries prevention: A scoping review. PLoS ONE 2020, 15, e0237547. [Google Scholar] [CrossRef] [PubMed]
- Slomka, V.; Hernandez-Sanabria, E.; Herrero, E.R.; Zaidel, L.; Bernaerts, K.; Boon, N.; Quirynen, M.; Teughels, W. Nutritional stimulation of commensal oral bacteria suppresses pathogens: The prebiotic concept. J. Clin. Periodontol. 2017, 44, 344–352. [Google Scholar] [CrossRef]
- Gasmi Benahmed, A.; Gasmi, A.; Arshad, M.; Shanaida, M.; Lysiuk, R.; Peana, M.; Pshyk-Titko, I.; Adamiv, S.; Shanaida, Y.; Bjorklund, G. Health benefits of xylitol. Appl. Microbiol. Biotechnol. 2020, 104, 7225–7237. [Google Scholar] [CrossRef]
- Haukioja, A.; Soderling, E.; Tenovuo, J. Acid production from sugars and sugar alcohols by probiotic lactobacilli and bifidobacteria in vitro. Caries Res. 2008, 42, 449–453. [Google Scholar] [CrossRef]
- Hedberg, M.; Hasslof, P.; Sjostrom, I.; Twetman, S.; Stecksen-Blicks, C. Sugar fermentation in probiotic bacteria—An in vitro study. Oral Microbiol. Immunol. 2008, 23, 482–485. [Google Scholar] [CrossRef]
- Salli, K.M.; Forssten, S.D.; Lahtinen, S.J.; Ouwehand, A.C. Influence of sucrose and xylitol on an early Streptococcus mutans biofilm in a dental simulator. Arch. Oral Biol. 2016, 70, 39–46. [Google Scholar] [CrossRef]
- Kaur, K.; Nekkanti, S.; Madiyal, M.; Choudhary, P. Effect of chewing gums containing probiotics and xylitol on oral health in children: A randomized controlled trial. J. Int. Oral Health 2018, 10, 7. [Google Scholar] [CrossRef]
- Kojima, Y.; Ohshima, T.; Seneviratne, C.J.; Maeda, N. Combining prebiotics and probiotics to develop novel synbiotics that suppress oral pathogens. J. Oral Biosci. 2016, 58, 6. [Google Scholar] [CrossRef]
- Cleaver, L.M.; Moazzez, R.V.; Carpenter, G.H. Evidence for Proline Utilization by Oral Bacterial Biofilms Grown in Saliva. Front. Microbiol. 2020, 11, 619968. [Google Scholar] [CrossRef] [PubMed]
- Christgen, S.L.; Becker, D.F. Role of Proline in Pathogen and Host Interactions. Antioxid. Redox Signal. 2019, 30, 683–709. [Google Scholar] [CrossRef]
- Culp, D.J.; Hull, W.; Bremgartner, M.J.; Atherly, T.A.; Christian, K.N.; Killeen, M.; Dupuis, M.R.; Schultz, A.C.; Chakraborty, B.; Lee, K.; et al. In Vivo Colonization with Candidate Oral Probiotics Attenuates Colonization and Virulence of Streptococcus mutans. Appl. Environ. Microbiol. 2020, 87, e02490-20. [Google Scholar] [CrossRef]
- Amargianitakis, M.; Antoniadou, M.; Rahiotis, C.; Varzakas, T. Probiotics, Prebiotics, Synbiotics and Dental Caries. New Perspectives, Suggestions, and Patient Coaching Approach for a Cavity-Free Mouth. Appl. Sci. 2021, 11, 5472. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. A Review of the Role of Probiotic Supplementation in Dental Caries. Probiotics Antimicrob. Proteins 2020, 12, 1300–1309. [Google Scholar] [CrossRef]
- Zheng, L.; Chen, Z.; Itzek, A.; Herzberg, M.C.; Kreth, J. CcpA regulates biofilm formation and competence in Streptococcus gordonii. Mol. Oral Microbiol. 2012, 27, 83–94. [Google Scholar] [CrossRef]
- Alforaidi, S.; Bresin, A.; Almosa, N.; Lehrkinder, A.; Lingstrom, P. Effect of drops containing Lactobacillus reuteri (DSM 17938 and ATCC PTA 5289) on plaque acidogenicity and other caries-related variables in orthodontic patients. BMC Microbiol. 2021, 21, 271. [Google Scholar] [CrossRef]
- Chen, Z.; Schlafer, S.; Gostemeyer, G.; Schwendicke, F. Probiotic Effects on Multispecies Biofilm Composition, Architecture, and Caries Activity In Vitro. Microorganisms 2020, 8, 1272. [Google Scholar] [CrossRef]
- Sliepen, I.; Van Damme, J.; Van Essche, M.; Loozen, G.; Quirynen, M.; Teughels, W. Microbial interactions influence inflammatory host cell responses. J. Dent. Res. 2009, 88, 1026–1030. [Google Scholar] [CrossRef]
- Cosseau, C.; Devine, D.A.; Dullaghan, E.; Gardy, J.L.; Chikatamarla, A.; Gellatly, S.; Yu, L.L.; Pistolic, J.; Falsafi, R.; Tagg, J.; et al. The commensal Streptococcus salivarius K12 downregulates the innate immune responses of human epithelial cells and promotes host-microbe homeostasis. Infect. Immun. 2008, 76, 4163–4175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Mendoza, A.; Liebana, J.; Castillo, A.M.; de la Higuera, A.; Piedrola, G. Evaluation of the capacity of oral streptococci to produce hydrogen peroxide. J. Med. Microbiol. 1993, 39, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Thurnheer, T.; Belibasakis, G.N. Streptococcus oralis maintains homeostasis in oral biofilms by antagonizing the cariogenic pathogen Streptococcus mutans. Mol. Oral Microbiol. 2018, 33, 234–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wescombe, P.A.; Heng, N.C.; Burton, J.P.; Chilcott, C.N.; Tagg, J.R. Streptococcal bacteriocins and the case for Streptococcus salivarius as model oral probiotics. Future Microbiol. 2009, 4, 819–835. [Google Scholar] [CrossRef] [Green Version]
- Burton, J.P.; Drummond, B.K.; Chilcott, C.N.; Tagg, J.R.; Thomson, W.M.; Hale, J.D.F.; Wescombe, P.A. Influence of the probiotic Streptococcus salivarius strain M18 on indices of dental health in children: A randomized double-blind, placebo-controlled trial. J. Med. Microbiol. 2013, 62, 875–884. [Google Scholar] [CrossRef]
- Zare Javid, A.; Amerian, E.; Basir, L.; Ekrami, A.; Haghighizadeh, M.H.; Maghsoumi-Norouzabad, L. Effects of the Consumption of Probiotic Yogurt Containing Bifidobacterium lactis Bb12 on the Levels of Streptococcus mutans and Lactobacilli in Saliva of Students with Initial Stages of Dental Caries: A Double-Blind Randomized Controlled Trial. Caries Res. 2020, 54, 68–74. [Google Scholar] [CrossRef]
- Tester, R.F.; Al-Ghazzewi, F.H. Role of prebiotics and probiotics in oral health. Nutr. Food Sci. 2018, 48, 14. [Google Scholar] [CrossRef]
- Jang, H.J.; Kim, J.H.; Lee, N.K.; Paik, H.D. Inhibitory effects of Lactobacillus brevis KU15153 against Streptococcus mutans KCTC 5316 causing dental caries. Microb. Pathog. 2021, 157, 104938. [Google Scholar] [CrossRef]
- Culp, D.J.; Hull, W.; Schultz, A.C.; Bryant, A.S.; Lizarraga, C.A.; Dupuis, M.R.; Chakraborty, B.; Lee, K.; Burne, R.A. Testing of candidate probiotics to prevent dental caries induced by Streptococcus mutans in a mouse model. J. Appl. Microbiol. 2022, 132, 3853–3869. [Google Scholar] [CrossRef]
- Hoffmann, A.; Fischer, J.T.; Daniels, R. Development of probiotic orodispersible tablets using mucoadhesive polymers for buccal mucoadhesion. Drug Dev. Ind. Pharm. 2020, 46, 1753–1762. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Ştefănescu, B.E.; Pop, I.D.; Muntean, L.; Vodnar, D.C. Chitosan Coating Applications in Probiotic Microencapsulation. Coatings 2019, 9, 194. [Google Scholar] [CrossRef] [Green Version]
- Yeung, T.W.; Ucok, E.F.; Tiani, K.A.; McClements, D.J.; Sela, D.A. Microencapsulation in Alginate and Chitosan Microgels to Enhance Viability of Bifidobacterium longum for Oral Delivery. Front. Microbiol. 2016, 7, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas-Jara, M.J.; Ilabaca, A.; Vega, M.; Garcia, A. Biofilm Forming Lactobacillus: New Challenges for the Development of Probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Erdelyi, L.; Fenyvesi, F.; Gal, B.; Haimhoffer, A.; Vasvari, G.; Budai, I.; Remenyik, J.; Bereczki, I.; Feher, P.; Ujhelyi, Z.; et al. Investigation of the Role and Effectiveness of Chitosan Coating on Probiotic Microcapsules. Polymers 2022, 14, 1664. [Google Scholar] [CrossRef]
- Kiew, T.Y.; Cheow, W.S.; Hadinoto, K. Importance of biofilm age and growth medium on the viability of probiotic capsules containing Lactobacillus rhamnosus GG biofilm. LWT-Food Sci. Technol. 2014, 59, 8. [Google Scholar] [CrossRef]
- Muras, A.; Mallo, N.; Otero-Casal, P.; Pose-Rodriguez, J.M.; Otero, A. Quorum sensing systems as a new target to prevent biofilm-related oral diseases. Oral Dis. 2022, 28, 307–313. [Google Scholar] [CrossRef]
- Cho, Y.J.; Song, H.Y.; Ben Amara, H.; Choi, B.K.; Eunju, R.; Cho, Y.A.; Seol, Y.; Lee, Y.; Ku, Y.; Rhyu, I.C.; et al. In Vivo Inhibition of Porphyromonas gingivalis Growth and Prevention of Periodontitis with Quorum-Sensing Inhibitors. J. Periodontol. 2016, 87, 1075–1082. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Inaba, T.; Obana, N.; Habe, H.; Nomura, N. Biofilm Formation by Streptococcus mutans is Enhanced by Indole via the Quorum Sensing Pathway. Microbes Environ. 2020, 35, ME19164. [Google Scholar] [CrossRef]
- Muras, A.; Mayer, C.; Romero, M.; Camino, T.; Ferrer, M.D.; Mira, A.; Otero, A. Inhibition of Steptococcus mutans biofilm formation by extracts of Tenacibaculum sp. 20J, a bacterium with wide-spectrum quorum quenching activity. J. Oral Microbiol. 2018, 10, 1429788. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.H.; Zhang, L.H. Quorum sensing and quorum-quenching enzymes. J. Microbiol. 2005, 43, 101–109. [Google Scholar] [PubMed]
- Choi, B.-K. Quorum Sensing Regulation of Biofilm Formation by Periodontal Pathogens. Int. J. Oral Biol. 2018, 43, 6. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, Y.; Shah, S.; Kim, D.; Simon-Soro, A.; Ito, T.; Hajfathalian, M.; Li, Y.; Hsu, J.C.; Nieves, L.M.; et al. Precision targeting of bacterial pathogen via bi-functional nanozyme activated by biofilm microenvironment. Biomaterials 2021, 268, 120581. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández, P.; Sánchez, M.C.; Llama-Palacios, A.; Ciudad, M.J.; Collado, L. Strategies to Combat Caries by Maintaining the Integrity of Biofilm and Homeostasis during the Rapid Phase of Supragingival Plaque Formation. Antibiotics 2022, 11, 880. https://doi.org/10.3390/antibiotics11070880
Hernández P, Sánchez MC, Llama-Palacios A, Ciudad MJ, Collado L. Strategies to Combat Caries by Maintaining the Integrity of Biofilm and Homeostasis during the Rapid Phase of Supragingival Plaque Formation. Antibiotics. 2022; 11(7):880. https://doi.org/10.3390/antibiotics11070880
Chicago/Turabian StyleHernández, Paola, María C. Sánchez, Arancha Llama-Palacios, María J. Ciudad, and Luis Collado. 2022. "Strategies to Combat Caries by Maintaining the Integrity of Biofilm and Homeostasis during the Rapid Phase of Supragingival Plaque Formation" Antibiotics 11, no. 7: 880. https://doi.org/10.3390/antibiotics11070880
APA StyleHernández, P., Sánchez, M. C., Llama-Palacios, A., Ciudad, M. J., & Collado, L. (2022). Strategies to Combat Caries by Maintaining the Integrity of Biofilm and Homeostasis during the Rapid Phase of Supragingival Plaque Formation. Antibiotics, 11(7), 880. https://doi.org/10.3390/antibiotics11070880