
Dietary Bacitracin Methylene Disalicylate Improves Growth Performance by Mediating the Gut Microbiota in Broilers


 and
and
Abstract
:1. Introduction
2. Results
2.1. Effects of BMD on Growth Performance
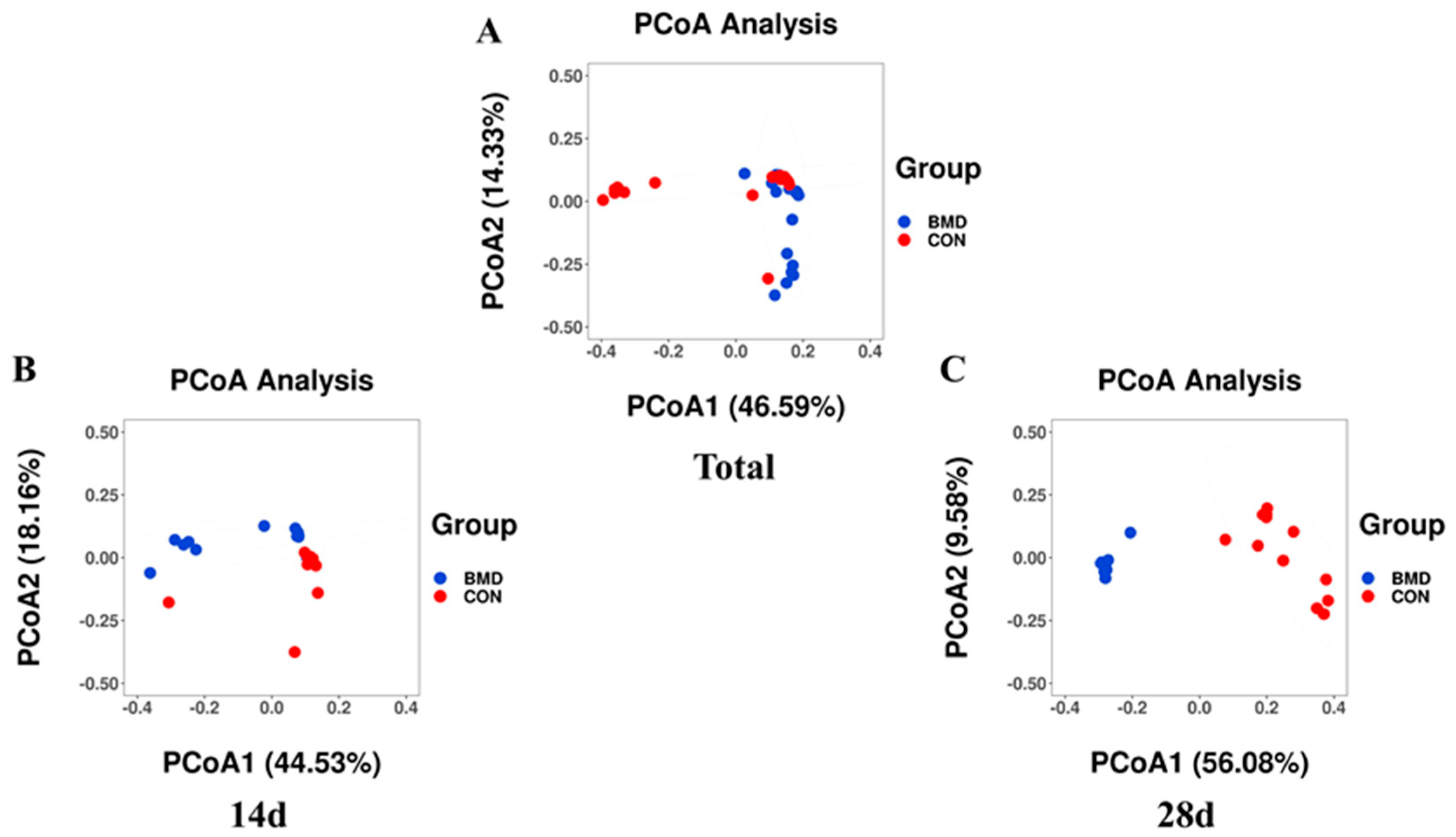
2.2. Effects of BMD on the Diversity and Structural Characteristics of Gut Microbiota in the Cecum
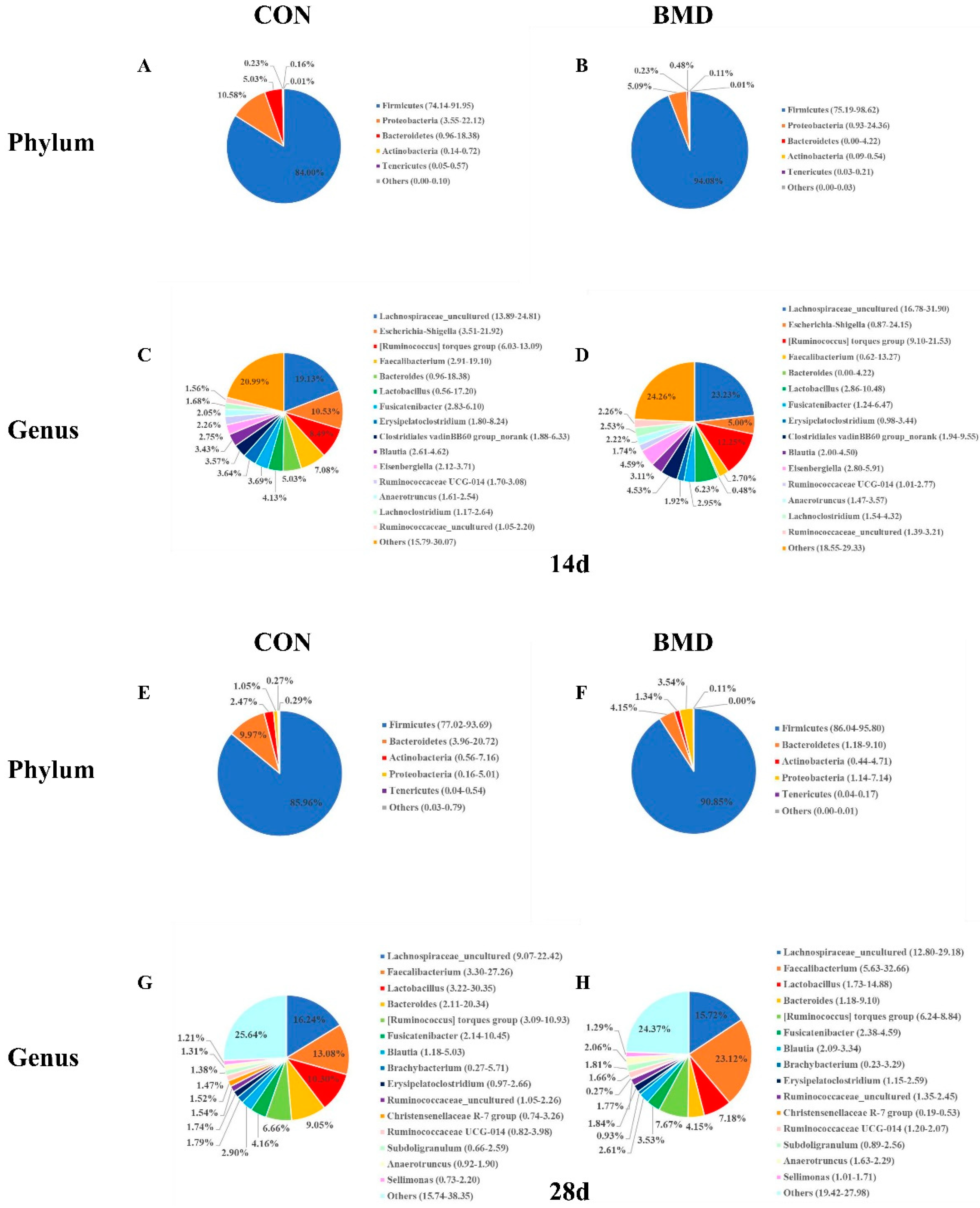
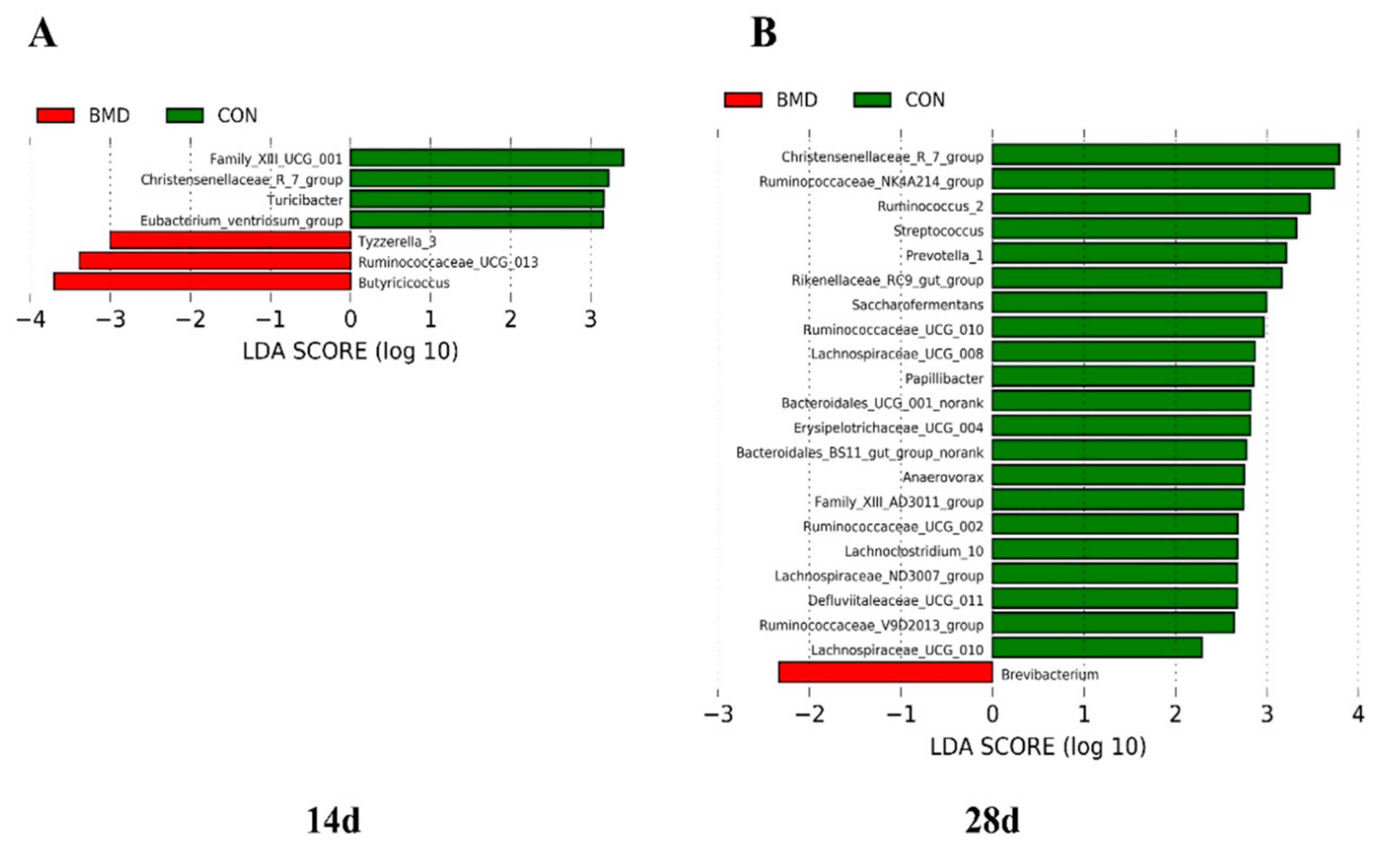
2.3. Effects of BMD on Gut Microbiota in the Cecum
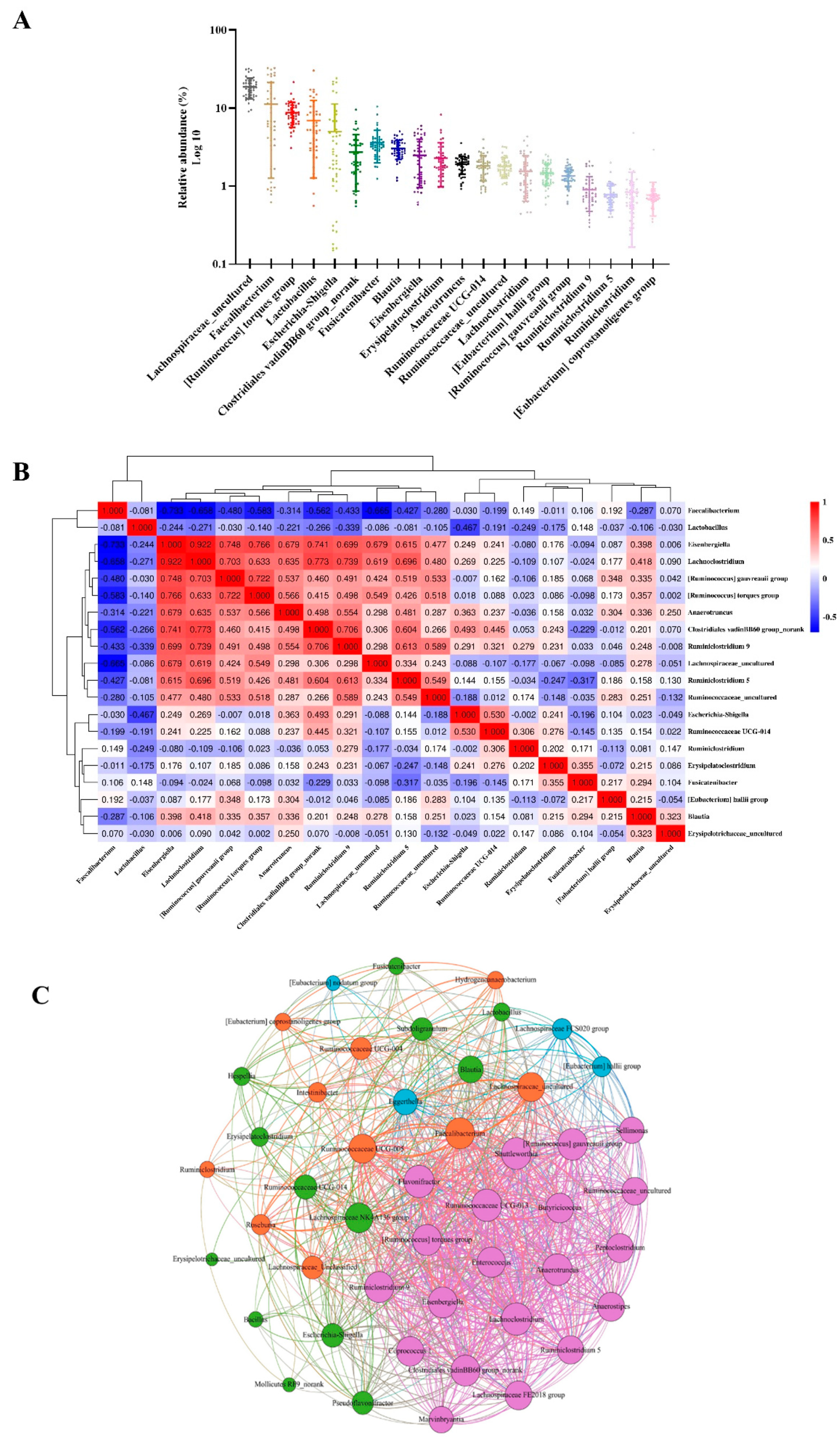
2.4. Genus-Level Core Gut Microbiota in the Caecum of Broilers
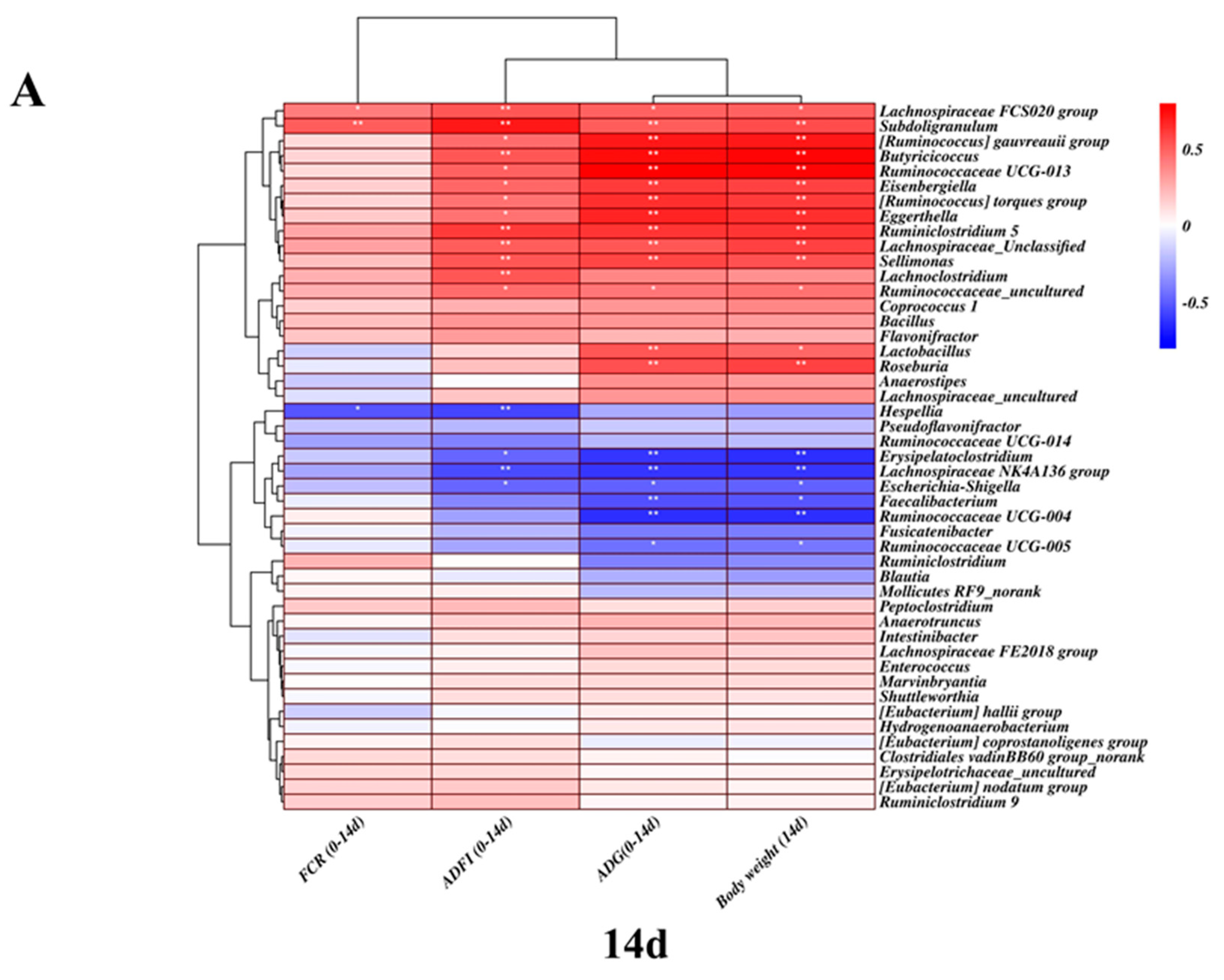
2.5. Correlation between the Microbiota Community and Its Growth Performance
3. Discussion
4. Materials and Methods
4.1. Animal Trials and Sample Collection
4.2. DNA Extraction and Sequencing
4.3. Bioinformatics Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rashidi, N.; Ghorbani, M.R.; Tatar, A.; Salari, S. Response of broiler chickens reared at high density to dietary supplementation with licorice extract and probiotic. J. Anim. Physiol. Anim. Nutr. 2019, 103, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Y.; Baek, D.H.; Kim, I.H. Effect of dietary supplemental medium chain fatty acids instead of antibiotics on the growth performance, digestibility and blood profiles in growing pigs. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1946–1951. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Lee, Y.; Goo, D.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.; Lillehoj, H.S. The effects of dietary Bacillus subtilis supplementation, as an alternative to antibiotics, on growth performance, intestinal immunity, and epithelial barrier integrity in broiler chickens infected with Eimeria maxima. Poult. Sci. 2020, 99, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Diana, A.; Boyle, L.A.; Leonard, F.C.; Carroll, C.; Sheehan, E.; Murphy, D.; Manzanilla, E.G. Removing prophylactic antibiotics from pig feed: How does it affect their performance and health? BMC Vet. Res. 2019, 15, 67. [Google Scholar] [CrossRef] [PubMed]
- Binversie, E.S.; Ruegg, P.L.; Combs, D.K.; Ollivett, T.L. Randomized clinical trial to assess the effect of antibiotic therapy on health and growth of preweaned dairy calves diagnosed with respiratory disease using respiratory scoring and lung ultrasound. J. Dairy Sci. 2020, 103, 11723–11735. [Google Scholar] [CrossRef]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef]
- Wohlleben, W.; Mast, Y.; Stegmann, E.; Ziemert, N. Antibiotic drug discovery. Microb. Biotechnol. 2016, 9, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Kemper, N.; Volkmann, N.; Schulz, J. Resistance of Enterococcus spp. in Dust from Farm Animal Houses: A Retrospective Study. Front. Microbiol. 2018, 9, 3074. [Google Scholar] [CrossRef]
- Hof, H. Is there a serious risk of resistance development to azoles among fungi due to the widespread use and long-term application of azole antifungals in medicine? Drug Resist. Updates 2008, 11, 25–31. [Google Scholar] [CrossRef]
- Bacanli, M.; Basaran, N. Importance of antibiotic residues in animal food. Food Chem. Toxicol. 2019, 125, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Muaz, K.; Riaz, M.; Akhtar, S.; Park, S.; Ismail, A. Antibiotic Residues in Chicken Meat: Global Prevalence, Threats, and Decontamination Strategies: A Review. J. Food Prot. 2018, 81, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, G.R.; Syed, B.; Haldar, S.; Pender, C. Phytogenic Feed Additives as an Alternative to Antibiotic Growth Promoters in Broiler Chickens. Front. Vet. Sci. 2015, 2, 21. [Google Scholar] [PubMed] [Green Version]
- Proctor, A.; Phillips, G.J. Differential Effects of Bacitracin Methylene Disalicylate (BMD) on the Distal Colon and Cecal Microbiota of Young Broiler Chickens. Front. Vet. Sci. 2019, 6, 114. [Google Scholar] [CrossRef]
- Gadde, U.D.; Oh, S.; Lillehoj, H.S.; Lillehoj, E.P. Antibiotic growth promoters virginiamycin and bacitracin methylene disalicylate alter the chicken intestinal metabolome. Sci. Rep. 2018, 8, 3592. [Google Scholar] [CrossRef] [Green Version]
- Droumev, D. Review of antimicrobial growth promoting agents available. Vet. Res. Commun. 1983, 7, 85–99. [Google Scholar] [CrossRef]
- Butaye, P.; Devriese, L.A.; Haesebrouck, F. Antimicrobial growth promoters used in animal feed: Effects of less well known antibiotics on gram-positive bacteria. Clin. Microbiol. Rev. 2003, 16, 175–188. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.L.; Weinberg, E.D. Mechanisms of antibacterial action of bacitracin. J. Gen. Microbiol. 1962, 28, 559–569. [Google Scholar] [CrossRef] [Green Version]
- Damron, B.L.; Wilson, H.R.; Fell, R.V. Growth and performance of broiler breeders fed bacitracin methylene disalicylate and zinc bacitracin. Poult. Sci. 1991, 70, 1487–1492. [Google Scholar] [CrossRef]
- Manafi, M.; Khalaji, S.; Hedayati, M.; Pirany, N. Efficacy of Bacillus subtilis and bacitracin methylene disalicylate on growth performance, digestibility, blood metabolites, immunity, and intestinal microbiota after intramuscular inoculation with Escherichia coli in broilers. Poult. Sci. 2017, 96, 1174–1183. [Google Scholar] [CrossRef]
- Kim, H.S.; Whon, T.W.; Sung, H.; Jeong, Y.S.; Jung, E.S.; Shin, N.R.; Hyun, D.-W.; Kim, P.S.; Lee, J.-Y.; Lee, C.H.; et al. Longitudinal evaluation of fecal microbiota transplantation for ameliorating calf diarrhea and improving growth performance. Nat. Commun. 2021, 12, 161. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Liu, S.; Hou, L.; Li, K.; Wang, L.; Gao, K.; Yang, X.; Jiang, Z. Supplemental Choline Modulates Growth Performance and Gut Inflammation by Altering the Gut Microbiota and Lipid Metabolism in Weaned Piglets. J. Nutr. 2021, 151, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Yu, B.; Degroote, J.; Spranghers, T.; Van Noten, N.; Majdeddin, M.; Van Poucke, M.; Peelman, L.; De Vrieze, J.; Boon, N.; et al. Antibiotic affects the gut microbiota composition and expression of genes related to lipid metabolism and myofiber types in skeletal muscle of piglets. BMC Vet. Res. 2020, 16, 392. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, C.; Liu, L.; Zhang, M. Lactobacillus reuteri KT260178 Supplementation Reduced Morbidity of Piglets through Its Targeted Colonization, Improvement of Cecal Microbiota Profile, and Immune Functions. Probiotics Antimicrob. Proteins 2020, 12, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Xie, Y.; Zhang, H.; Jin, J.; Xiong, L.; Liu, H. Effects of a probiotic-fermented herbal blend on the growth performance, intestinal flora and immune function of chicks infected with Salmonella pullorum. Poult. Sci. 2021, 100, 101196. [Google Scholar] [CrossRef]
- Xiao, X.; Fu, Z.; Li, N.; Yang, H.; Wang, W.; Lyu, W. Modulation of the Intestinal Microbiota by the Early Intervention with Clostridium Butyricum in Muscovy Ducks. Antibiotics 2021, 10, 826. [Google Scholar] [CrossRef]
- Gong, Y.; Yang, H.; Wang, X.; Xia, W.; Lv, W.; Xiao, Y.; Zou, X. Early Intervention with Cecal Fermentation Broth Regulates the Colonization and Development of Gut Microbiota in Broiler Chickens. Front. Microbiol. 2019, 10, 1422. [Google Scholar] [CrossRef]
- Asiamah, C.; Xue, Y.; Lu, L.L.; Zou, K.; Zhao, Z.; Su, Y. Evaluation of growth performance on family breeding of the Leizhou Black Duck: A preliminary study. Vet. Med. Sci. 2020, 6, 500–510. [Google Scholar] [CrossRef]
- Hermesch, S.; Ludemann, C.I.; Amer, P.R. Economic weights for performance and survival traits of growing pigs. J. Anim. Sci. 2014, 92, 5358–5366. [Google Scholar] [CrossRef]
- Xu, T.; Xu, S.; Hu, L.; Zhao, N.; Liu, Z.; Ma, L.; Liu, H.; Zhao, X. Effect of Dietary Types on Feed Intakes, Growth Performance and Economic Benefit in Tibetan sheep and Yaks on the Qinghai-Tibet Plateau during Cold Season. PLoS ONE 2017, 12, e0169187. [Google Scholar] [CrossRef]
- Madzimure, J.; Motsi, R.; Bakare, A.G.; Zimondi, M. Growth performance, economic benefits and sensory characteristics of meat from broilers finished on Acacia anguistissima leaf meal-based diets. Trop. Anim. Health Prod. 2018, 50, 1485–1491. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Haldar, S.; Ghosh, T.K. Comparative efficacy of an organic Acid blend and bacitracin methylene disalicylate as growth promoters in broiler chickens: Effects on performance, gut histology, and small intestinal milieu. Vet. Med. Int. 2010, 2010, 645150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoeler, M.; Caesar, R. Dietary lipids, gut microbiota and lipid metabolism. Rev. Endocr. Metab. Disord. 2019, 20, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Brussow, H. Growth promotion and gut microbiota: Insights from antibiotic use. Environ. Microbiol. 2015, 17, 2216–2227. [Google Scholar] [CrossRef] [PubMed]
- Butel, M.J.; Waligora-Dupriet, A.J.; Wydau-Dematteis, S. The developing gut microbiota and its consequences for health. J. Dev. Orig. Health Dis. 2018, 9, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Adewole, D.; Akinyemi, F. Gut Microbiota Dynamics, Growth Performance, and Gut Morphology in Broiler Chickens Fed Diets Varying in Energy Density with or without Bacitracin Methylene Disalicylate (BMD). Microorganisms 2021, 9, 787. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Pan, X.; Cao, S.; Xu, F.; Lan, L.; Zhang, Y.; Lian, S.; Yan, M.; Li, A. iTRAQ-based quantitative proteomic analysis provides insights into strong broodiness in Muscovy duck (Cairina moschata) combined with metabolomics analysis. J. Proteom. 2019, 204, 103401. [Google Scholar] [CrossRef]
- Vermeulen, K.; Verspreet, J.; Courtin, C.M.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Reduced particle size wheat bran is butyrogenic and lowers Salmonella colonization, when added to poultry feed. Vet. Microbiol. 2017, 198, 64–71. [Google Scholar] [CrossRef]
- Pourabedin, M.; Guan, L.; Zhao, X. Xylo-oligosaccharides and virginiamycin differentially modulate gut microbial composition in chickens. Microbiome 2015, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Rychlik, I. Composition and Function of Chicken Gut Microbiota. Animals 2020, 10, 103. [Google Scholar] [CrossRef] [Green Version]
- Sohail, M.U.; Hume, M.E.; Byrd, J.A.; Nisbet, D.J.; Shabbir, M.Z.; Ijaz, A.; Rehman, H. Molecular analysis of the caecal and tracheal microbiome of heat-stressed broilers supplemented with prebiotic and probiotic. Avian Pathol. 2015, 44, 67–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.; Sun, P.; Zhang, B.; Kong, L.; Xiao, C.; Song, Z. Progress on Gut Health Maintenance and Antibiotic Alternatives in Broiler Chicken Production. Front. Nutr. 2021, 8, 692839. [Google Scholar] [CrossRef] [PubMed]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Nordentoft, S.; Molbak, L.; Bjerrum, L.; De Vylder, J.; Van Immerseel, F.; Pedersen, K. The influence of the cage system and colonisation of Salmonella Enteritidis on the microbial gut flora of laying hens studied by T-RFLP and 454 pyrosequencing. BMC Microbiol. 2011, 11, 187. [Google Scholar] [CrossRef] [Green Version]
- Richards, P.; Fothergill, J.; Bernardeau, M.; Wigley, P. Development of the Caecal Microbiota in Three Broiler Breeds. Front. Vet. Sci. 2019, 6, 201. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Li, J.; Xing, T.; Jiang, Y.; Zhang, L.; Gao, F. Dietary resistant starch modifies the composition and function of caecal microbiota of broilers. J. Sci. Food Agric. 2020, 100, 1274–1284. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Zakham, F.; Pillonel, T.; Brunel, A.-S.; Zambelli, P.-Y.; Greub, G.; Croxatto, A.; Bertelli, C. Molecular diagnosis and enrichment culture identified a septic pseudoarthrosis due to an infection with Erysipelatoclostridium ramosum. Int. J. Infect. Dis. 2019, 81, 167–169. [Google Scholar] [CrossRef] [Green Version]
- Eeckhaut, V.; Wang, J.; Van Parys, A.; Haesebrouck, F.; Joossens, M.; Falony, G.; Raes, J.; Ducatelle, R.; van Immerseel, F. The Probiotic Butyricicoccus pullicaecorum Reduces Feed Conversion and Protects from Potentially Harmful Intestinal Microorganisms and Necrotic Enteritis in Broilers. Front. Microbiol. 2016, 7, 1416. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.C.; Shen, M.H.; Liu, C.Y.; Pu, C.M.; Hu, J.M.; Huang, C.J. A gut butyrate-producing bacterium Butyricicoccus pullicaecorum regulates short-chain fatty acid transporter and receptor to reduce the progression of 1,2-dimethylhydrazine-associated colorectal cancer. Oncol. Lett. 2020, 20, 327. [Google Scholar] [CrossRef]
- Robinson, K.; Xiao, Y.; Johnson, T.J.; Chen, B.; Yang, Q.; Lyu, W.; Wang, J.; Fansler, N.; Becker, S.; Liu, J.; et al. Chicken Intestinal Mycobiome: Initial Characterization and Its Response to Bacitracin Methylene Disalicylate. Appl. Environ. Microbiol. 2020, 86, e00304-20. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xiang, Y.; Robinson, K.; Wang, J.; Zhang, G.; Zhao, J.; Xiao, Y. Gut Microbiota Is a Major Contributor to Adiposity in Pigs. Front. Microbiol. 2018, 9, 3045. [Google Scholar] [CrossRef] [PubMed]
- Gossiel, F.; Altaher, H.; Reid, D.M.; Roux, C.; Felsenberg, D.; Glüer, C.C.; Eastell, R. Bone turnover markers after the menopause: T-score approach. Bone 2018, 111, 44–48. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | CON | BMD | SEM | p-Value |
|---|---|---|---|---|
| Body weight (g) | ||||
| Day 0 | 45.58 | 45.81 | 0.36 | 0.755 |
| Day 14 | 393.57 | 442.59 | 5.32 | <0.001 |
| Day 28 | 1088.03 | 1362.18 | 29.71 | <0.001 |
| Average daily gain (g) | ||||
| 0–14 d | 24.86 | 28.34 | 0.38 | <0.001 |
| 15–28 d | 49.60 | 65.69 | 1.77 | <0.001 |
| 0–28 d | 37.23 | 47.01 | 1.06 | <0.001 |
| Average daily feed intake (g/d) | ||||
| 0–14 d | 40.58 | 49.66 | 1.32 | <0.001 |
| 15–28 d | 99.49 | 137.02 | 4.12 | <0.001 |
| 0–28 d | 70.03 | 93.34 | 2.56 | <0.001 |
| Feed conversion (g/g) | ||||
| 0–14 d | 1.64 | 1.75 | 0.04 | 0.123 |
| 15–28 d | 2.00 | 2.09 | 0.03 | 0.168 |
| 0–28 d | 1.88 | 1.99 | 0.03 | 0.037 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Xiao, Y.; Fan, Q.; Yang, H.; Yang, C.; Zhang, G.; Chen, S. Dietary Bacitracin Methylene Disalicylate Improves Growth Performance by Mediating the Gut Microbiota in Broilers. Antibiotics 2022, 11, 818. https://doi.org/10.3390/antibiotics11060818
Li J, Xiao Y, Fan Q, Yang H, Yang C, Zhang G, Chen S. Dietary Bacitracin Methylene Disalicylate Improves Growth Performance by Mediating the Gut Microbiota in Broilers. Antibiotics. 2022; 11(6):818. https://doi.org/10.3390/antibiotics11060818
Chicago/Turabian StyleLi, Jingshang, Yingping Xiao, Qian Fan, Hua Yang, Caimei Yang, Guolong Zhang, and Shengchang Chen. 2022. "Dietary Bacitracin Methylene Disalicylate Improves Growth Performance by Mediating the Gut Microbiota in Broilers" Antibiotics 11, no. 6: 818. https://doi.org/10.3390/antibiotics11060818
APA StyleLi, J., Xiao, Y., Fan, Q., Yang, H., Yang, C., Zhang, G., & Chen, S. (2022). Dietary Bacitracin Methylene Disalicylate Improves Growth Performance by Mediating the Gut Microbiota in Broilers. Antibiotics, 11(6), 818. https://doi.org/10.3390/antibiotics11060818