
Impact of Subinhibitory Concentrations of Metronidazole on Morphology, Motility, Biofilm Formation and Colonization of Clostridioides difficile

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
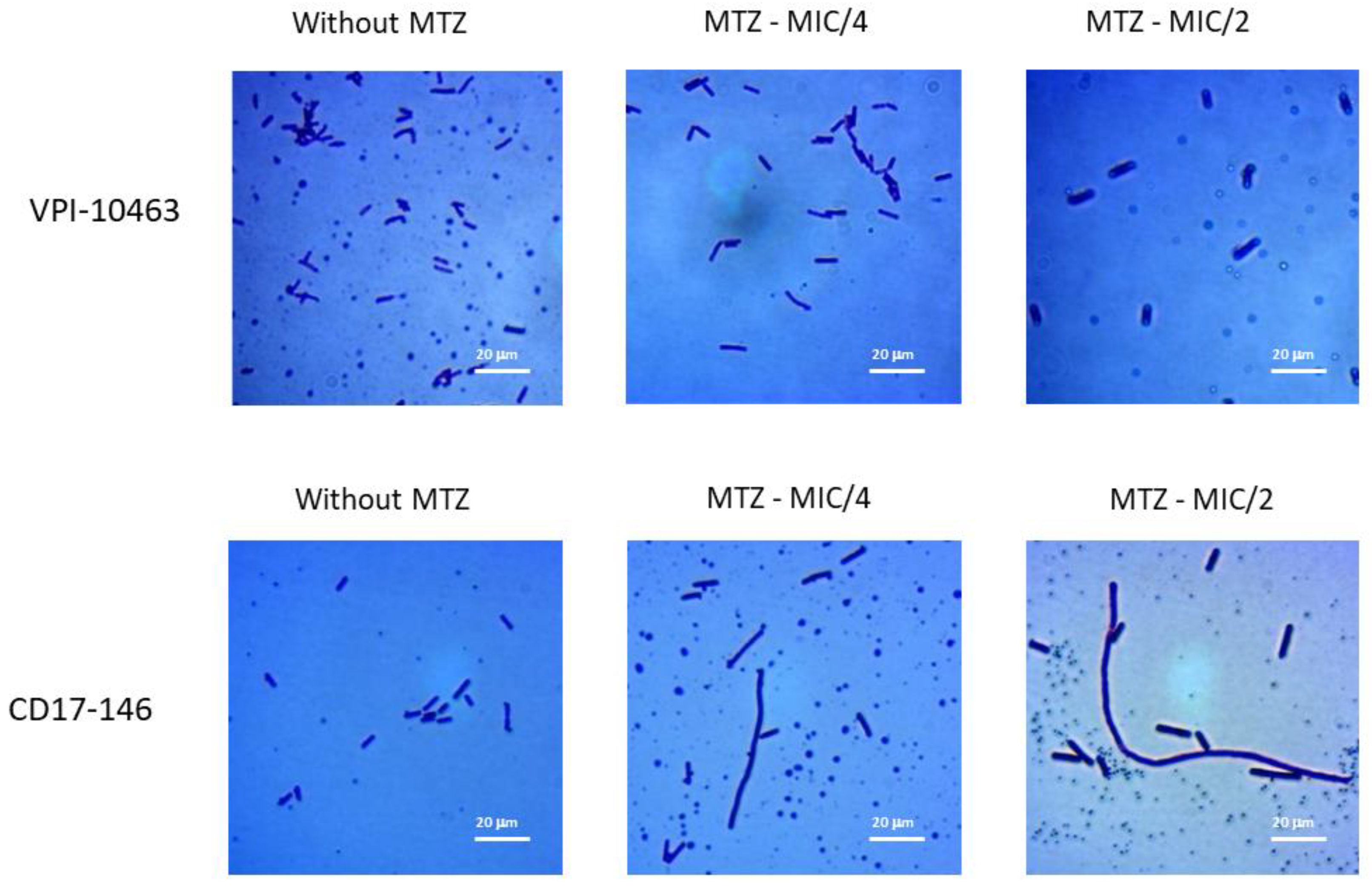
2.1. Impact of MTZ on the Morphology of C. difficile
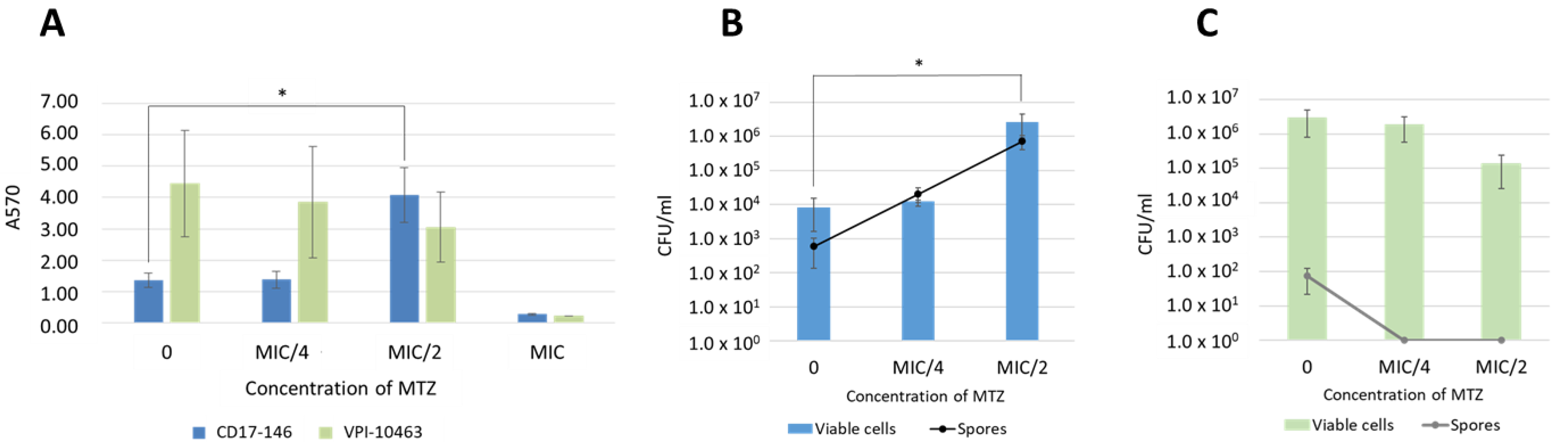
2.2. C. difficile Biofilm Production in Absence and Presence of MTZ
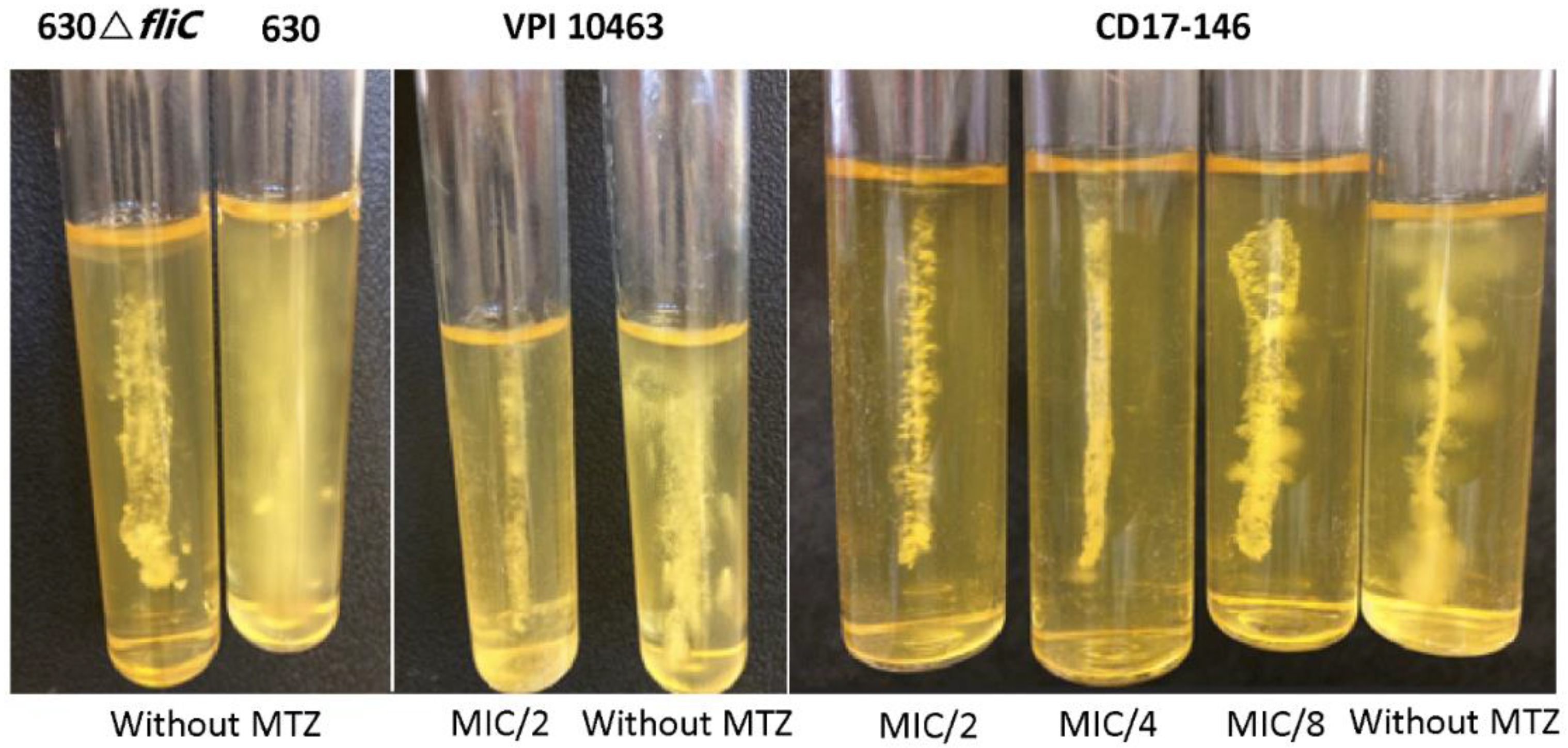
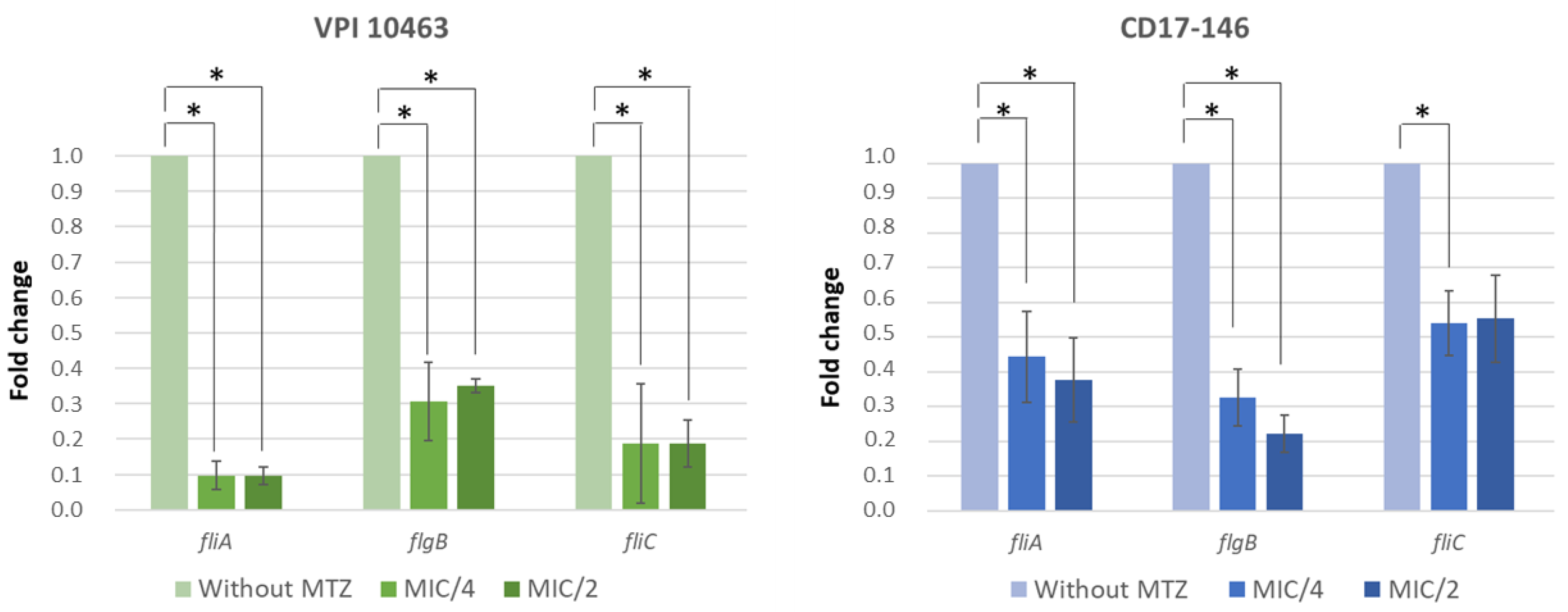
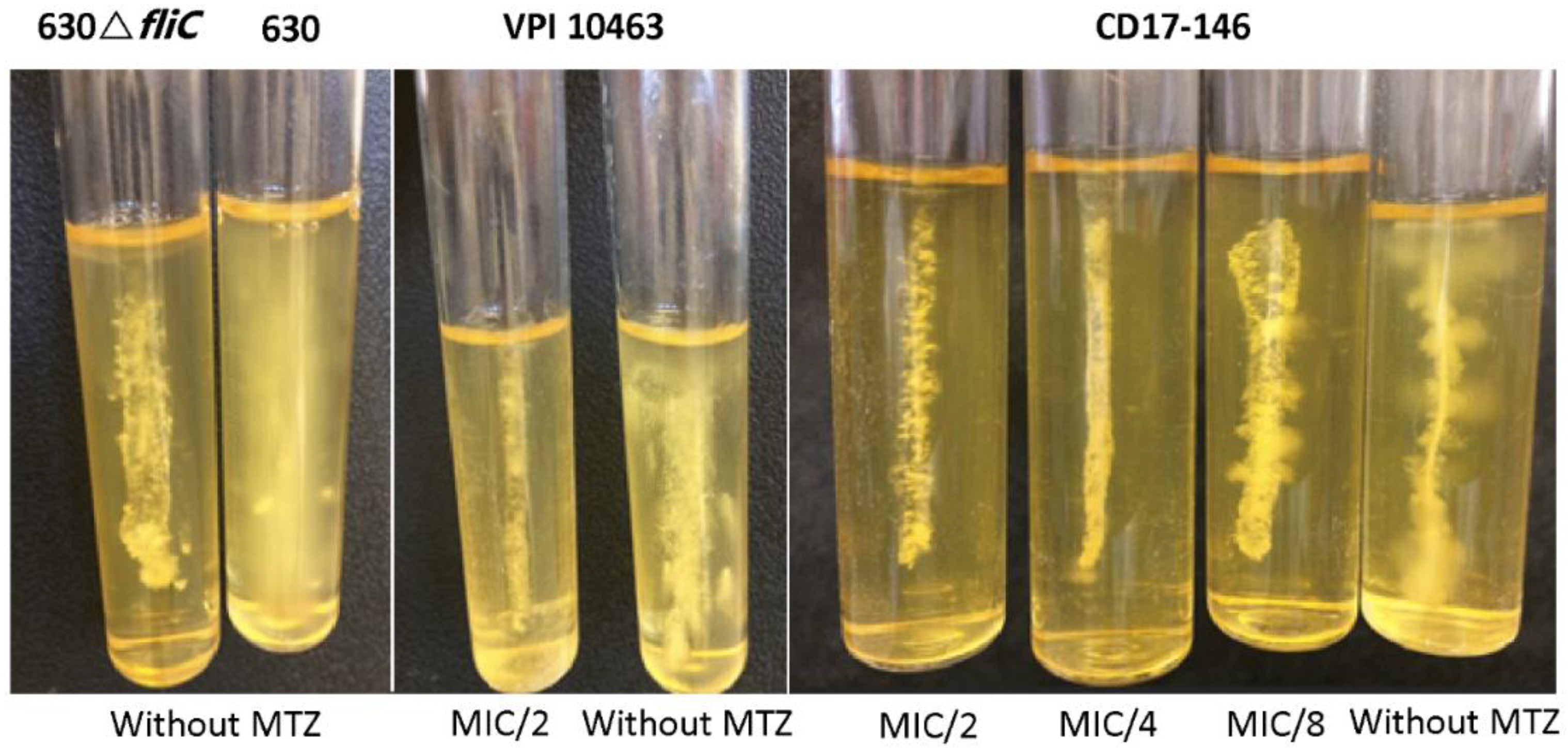
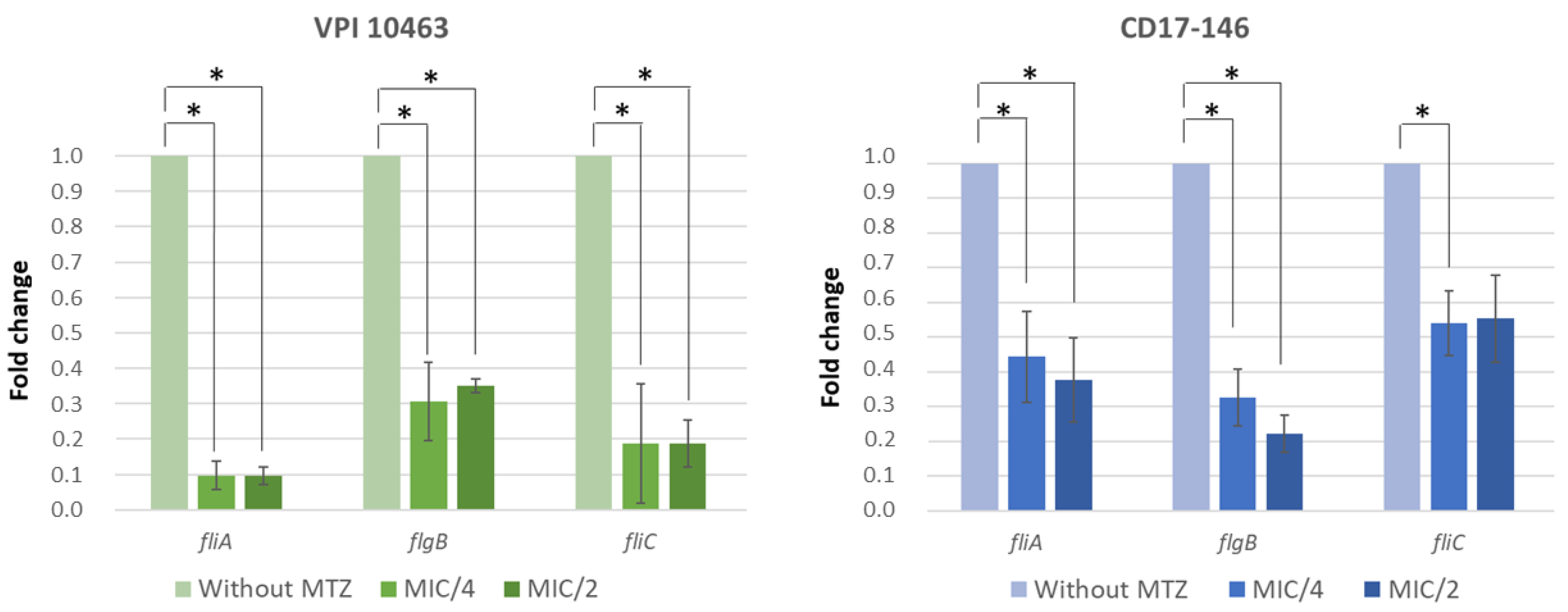
2.3. Impact of MTZ on the Motility of C. difficile and Transcriptions of Flagellar Genes
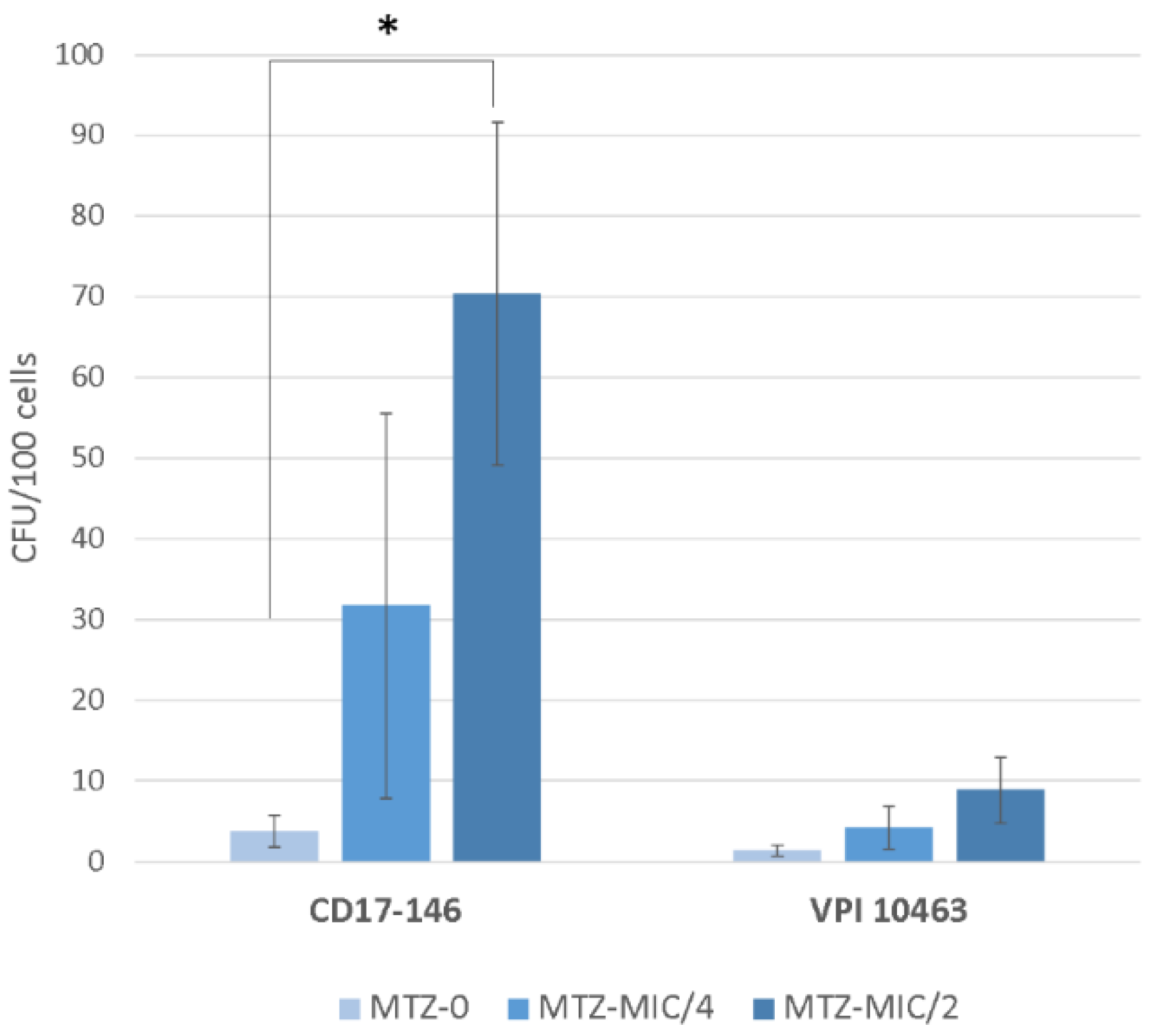
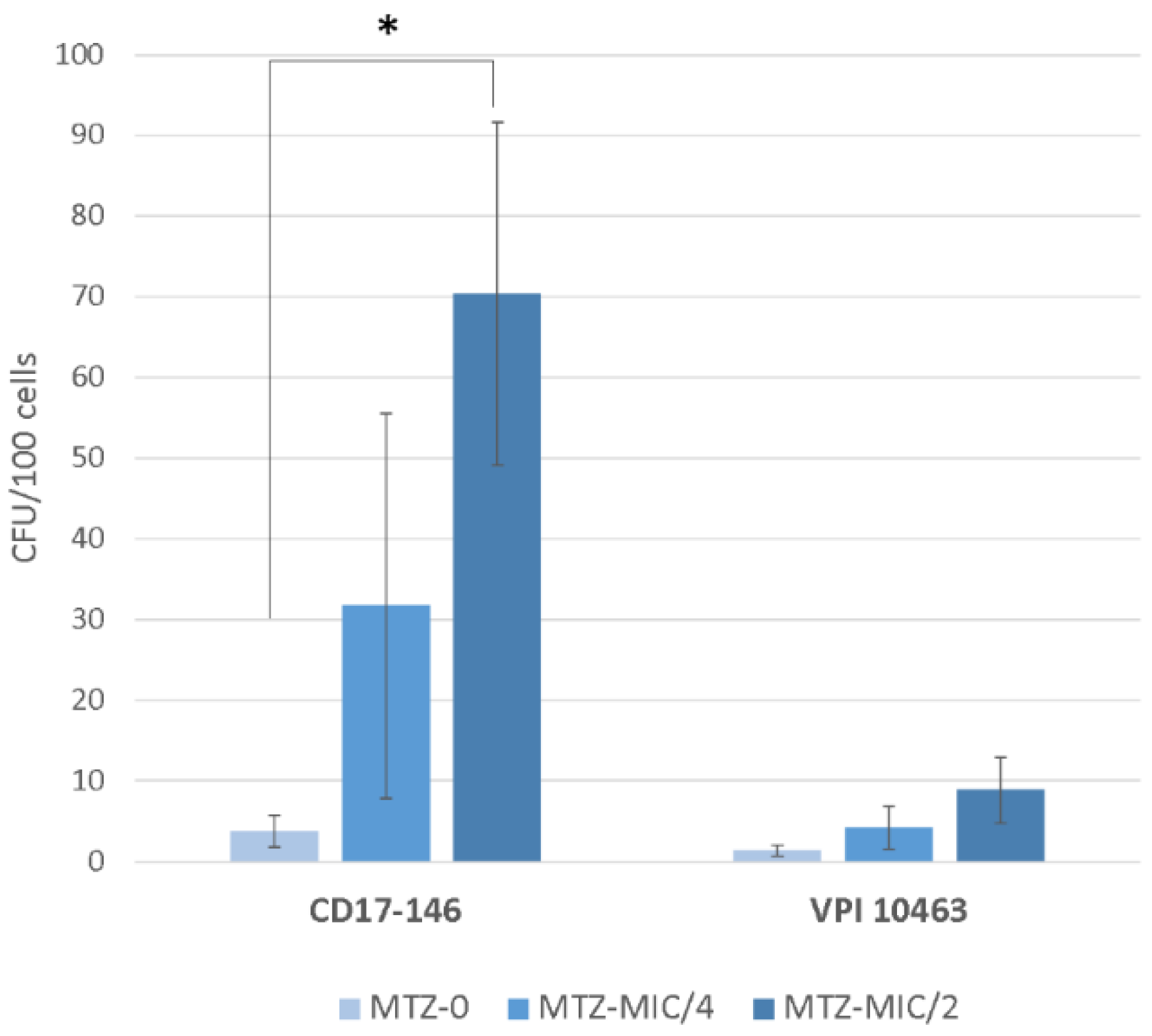
2.4. Effect of Subinhibitory Concentrations of MTZ on C. difficile Adherence
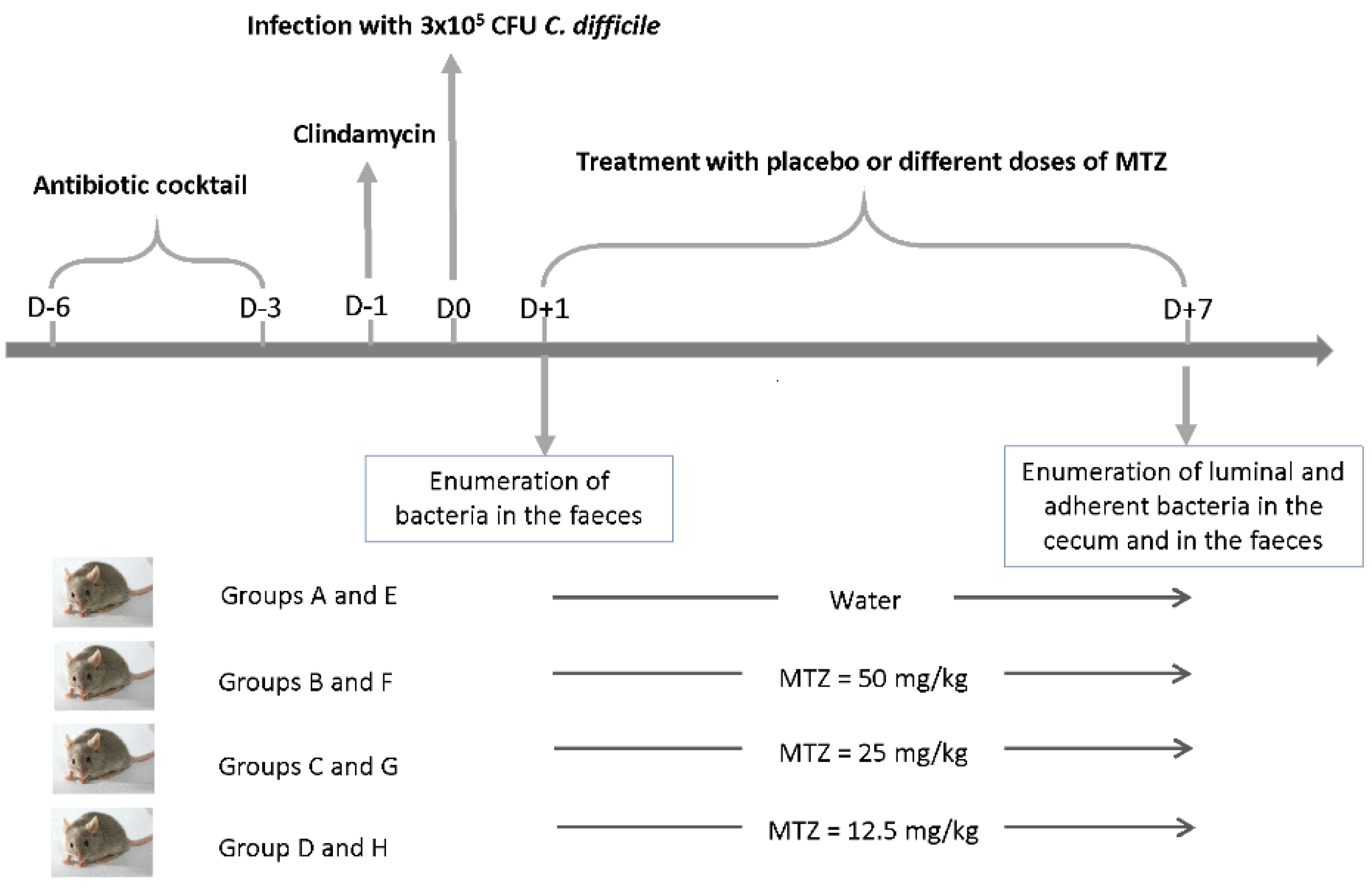
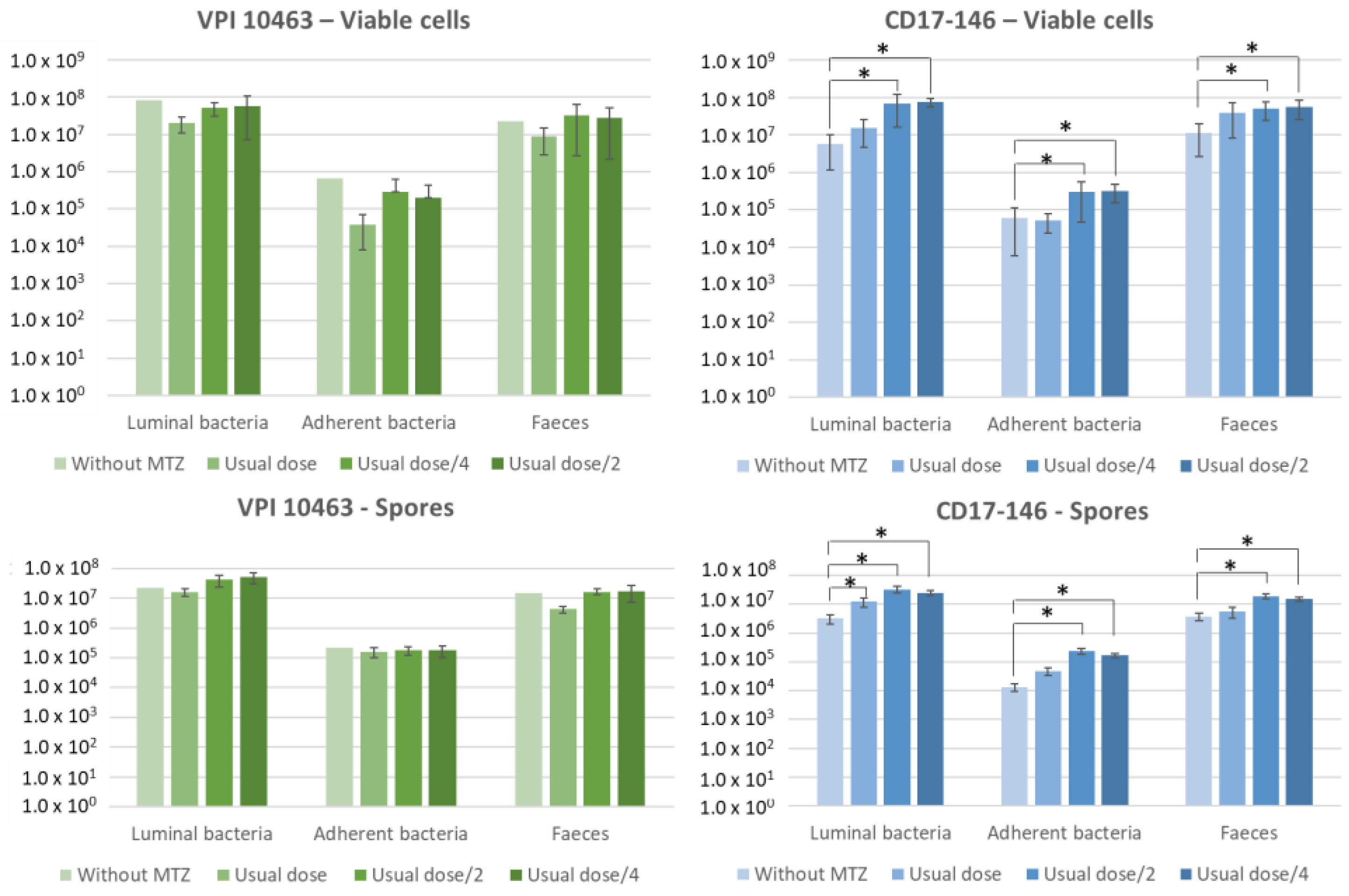
2.5. Subinhibitory Concentrations of MTZ Stimulate Cecum Colonization by CD17-146 in the Conventional Mouse Model
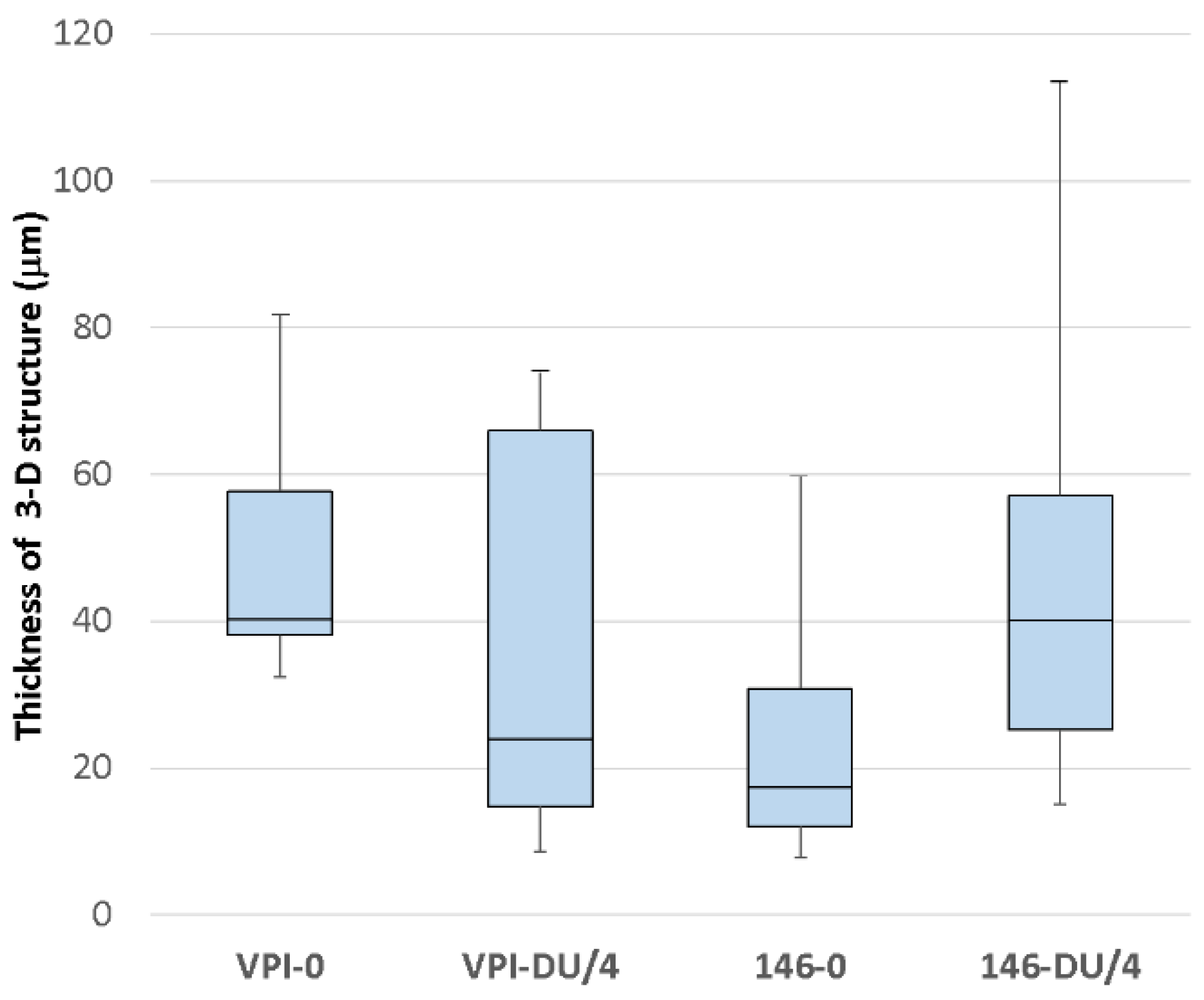
2.6. Visualisation of Bacterial Distribution in the Cecum by CLSM
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Antibiotic Susceptibility
4.2. Morphology Observation
4.3. In Vitro Biofilm
4.4. Motility Assays
4.5. RNA Extraction and Quantitative RT-PCR Analysis on Flagellar Genes
4.6. Adherence Assays
4.7. Model of Conventional Mice to Study the Impact of MTZ on the Intestinal Colonization of C. difficile
4.8. Monoxenic Mouse Model to Visualize C. difficile Intestinal Distribution in the Cecum When Exposed to MTZ
4.8.1. Animal Model
4.8.2. Confocal Laser Scanning Microscopy (CLSM)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- De Roo, A.C.; Regenbogen, S.E. Clostridium difficile Infection: An Epidemiology Update. Clin. Colon Rectal Surg. 2020, 33, 49–57. [Google Scholar] [CrossRef] [PubMed]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; Coffin, S.E.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical Practice Guidelines for Clostridium difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, e1–e48. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Louie, T.J.; Gerding, D.N.; Cornely, O.; Chasan-Taber, S.; Fitts, D.; Gelone, S.P.; Broom, C.; Davidson, D.M. Polymer Alternative for CDI Treatment (PACT) investigators Vancomycin, Metronidazole, or Tolevamer for Clostridium difficile Infection: Results from Two Multinational, Randomized, Controlled Trials. Clin. Infect. Dis. 2014, 59, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, A.H.; Lam, N.P.; Piscitelli, S.C.; Wilkes, L.; Danziger, L.H. Clinical Pharmacokinetics of Metronidazole and Other Nitroimidazole Anti-Infectives. Clin. Pharmacokinet. 1992, 23, 328–364. [Google Scholar] [CrossRef]
- Bolton, R.P.; Culshaw, A.M. Faecal Metronidazole Concentrations during Oral and Intravenous Therapy for Antibiotic Associated Colitis Due to Clostridium Difficile. Gut 1986, 27, 1169–1172. [Google Scholar] [CrossRef] [Green Version]
- Peláez, T.; Cercenado, E.; Alcalá, L.; Marín, M.; Martín-López, A.; Martínez-Alarcón, J.; Catalán, P.; Sánchez-Somolinos, M.; Bouza, E. Metronidazole Resistance in Clostridium difficile Is Heterogeneous. J. Clin. Microbiol. 2008, 46, 3028–3032. [Google Scholar] [CrossRef] [Green Version]
- Andersson, D.I.; Hughes, D. Microbiological Effects of Sublethal Levels of Antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef]
- Moura, I.B.; Spigaglia, P.; Barbanti, F.; Mastrantonio, P. Analysis of Metronidazole Susceptibility in Different Clostridium difficile PCR Ribotypes. J. Antimicrob. Chemother. 2013, 68, 362–365. [Google Scholar] [CrossRef] [Green Version]
- Vuotto, C.; Moura, I.; Barbanti, F.; Donelli, G.; Spigaglia, P. Subinhibitory Concentrations of Metronidazole Increase Biofilm Formation in Clostridium difficile Strains. Pathog. Dis. 2016, 74, ftv114. [Google Scholar] [CrossRef] [Green Version]
- Ðapa, T.; Dapa, T.; Leuzzi, R.; Ng, Y.K.; Baban, S.T.; Adamo, R.; Kuehne, S.A.; Scarselli, M.; Minton, N.P.; Serruto, D.; et al. Multiple Factors Modulate Biofilm Formation by the Anaerobic Pathogen Clostridium difficile. J. Bacteriol. 2013, 195, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Surawicz, C.M.; Alexander, J. Treatment of Refractory and Recurrent Clostridium difficile Infection. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 330–339. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Stoodley, P. Evolving Concepts in Biofilm Infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef]
- Li, D.; Renzoni, A.; Estoppey, T.; Bisognano, C.; Francois, P.; Kelley, W.L.; Lew, D.P.; Schrenzel, J.; Vaudaux, P. Induction of Fibronectin Adhesins in Quinolone-Resistant Staphylococcus aureus by Subinhibitory Levels of Ciprofloxacin or by Sigma B Transcription Factor Activity Is Mediated by Two Separate Pathways. Antimicrob. Agents Chemother. 2005, 49, 916–924. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, L.R.; D’Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside Antibiotics Induce Bacterial Biofilm Formation. Nature 2005, 436, 1171–1175. [Google Scholar] [CrossRef]
- Moura, I.; Monot, M.; Tani, C.; Spigaglia, P.; Barbanti, F.; Norais, N.; Dupuy, B.; Bouza, E.; Mastrantonio, P. Multidisciplinary Analysis of a Nontoxigenic Clostridium difficile Strain with Stable Resistance to Metronidazole. Antimicrob. Agents Chemother. 2014, 58, 4957–4960. [Google Scholar] [CrossRef] [Green Version]
- Boekhoud, I.M.; Hornung, B.V.H.; Sevilla, E.; Harmanus, C.; Bos-Sanders, I.M.J.G.; Terveer, E.M.; Bolea, R.; Corver, J.; Kuijper, E.J.; Smits, W.K. Plasmid-Mediated Metronidazole Resistance in Clostridioides difficile. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Alauzet, C.; Lozniewski, A.; Marchandin, H. Metronidazole Resistance and Nim Genes in Anaerobes: A Review. Anaerobe 2019, 55, 40–53. [Google Scholar] [CrossRef]
- Deshpande, A.; Wu, X.; Huo, W.; Palmer, K.L.; Hurdle, J.G. Chromosomal Resistance to Metronidazole in Clostridioides difficile Can Be Mediated by Epistasis between Iron Homeostasis and Oxidoreductases. Antimicrob. Agents Chemother. 2020, 64, e00415-20. [Google Scholar] [CrossRef]
- Britz, M.L.; Wilkinson, R.G. Isolation and Properties of Metronidazole-Resistant Mutants of Bacteroides fragilis. Antimicrob. Agents Chemother. 1979, 16, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.; Salem, A.; Coombs, G.H. The In-Vitro Activity of Metronidazole against Strains of Escherichia coli with Impaired DNA Repair Systems. J. Antimicrob. Chemother. 1984, 13, 227–236. [Google Scholar] [CrossRef]
- Tally, F.P.; Sutter, V.L.; Finegold, S.M. Treatment of Anaerobic Infections with Metronidazole. Antimicrob. Agents Chemother. 1975, 7, 672–675. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.W.; Lee, S.Y. Effect of Sub-Minimal Inhibitory Concentration Antibiotics on Morphology of Periodontal Pathogens. Int. J. Oral. Biol. 2014, 39, 115–120. [Google Scholar] [CrossRef]
- Bedran, T.B.L.; Grignon, L.; Spolidorio, D.P.; Grenier, D. Subinhibitory Concentrations of Triclosan Promote Streptococcus mutans Biofilm Formation and Adherence to Oral Epithelial Cells. PLoS ONE 2014, 9, e89059. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Li, X.; Gunawardana, M.; Maguire, K.; Guerrero-Given, D.; Schaudinn, C.; Wang, C.; Baum, M.M.; Webster, P. Beta- Lactam Antibiotics Stimulate Biofilm Formation in Non-Typeable Haemophilus influenzae by Up-Regulating Carbohydrate Metabolism. PLoS ONE 2014, 9, e99204. [Google Scholar] [CrossRef] [Green Version]
- Abu Rahmoun, L.; Azrad, M.; Peretz, A. Antibiotic Resistance and Biofilm Production Capacity in Clostridioides difficile. Front. Cell. Infect. Microbiol. 2021, 11, 683464. [Google Scholar] [CrossRef]
- Dingle, T.C.; Mulvey, G.L.; Armstrong, G.D. Mutagenic Analysis of the Clostridium difficile Flagellar Proteins, FliC and FliD, and Their Contribution to Virulence in Hamsters. Infect. Immun. 2011, 79, 4061–4067. [Google Scholar] [CrossRef] [Green Version]
- Baban, S.T.; Kuehne, S.A.; Barketi-Klai, A.; Cartman, S.T.; Kelly, M.L.; Hardie, K.R.; Kansau, I.; Collignon, A.; Minton, N.P. The Role of Flagella in Clostridium difficile Pathogenesis: Comparison between a Non-Epidemic and an Epidemic Strain. PLoS ONE 2013, 8, e73026. [Google Scholar] [CrossRef]
- Kasendra, M.; Barrile, R.; Leuzzi, R.; Soriani, M. Clostridium difficile Toxins Facilitate Bacterial Colonization by Modulating the Fence and Gate Function of Colonic Epithelium. J. Infect. Dis. 2014, 209, 1095–1104. [Google Scholar] [CrossRef] [Green Version]
- Erikstrup, L.T.; Aarup, M.; Hagemann-Madsen, R.; Dagnaes-Hansen, F.; Kristensen, B.; Olsen, K.E.P.; Fuursted, K. Treatment of Clostridium difficile Infection in Mice with Vancomycin Alone Is as Effective as Treatment with Vancomycin and Metronidazole in Combination. BMJ Open Gastroenterol. 2015, 2, e000038. [Google Scholar] [CrossRef] [Green Version]
- Cooper, R.; Bjarnsholt, T.; Alhede, M. Biofilms in Wounds: A Review of Present Knowledge. J. Wound Care 2014, 23, 570–580. [Google Scholar] [CrossRef] [Green Version]
- Mihai, M.M.; Holban, A.M.; Giurcaneanu, C.; Popa, L.G.; Oanea, R.M.; Lazar, V.; Chifiriuc, M.C.; Popa, M.; Popa, M.I. Microbial Biofilms: Impact on the Pathogenesis of Periodontitis, Cystic Fibrosis, Chronic Wounds and Medical Device-Related Infections. Curr. Top. Med. Chem. 2015, 15, 1552–1576. [Google Scholar] [CrossRef] [PubMed]
- Lund-Palau, H.; Turnbull, A.R.; Bush, A.; Bardin, E.; Cameron, L.; Soren, O.; Wierre-Gore, N.; Alton, E.W.F.W.; Bundy, J.G.; Connett, G.; et al. Pseudomonas aeruginosa Infection in Cystic Fibrosis: Pathophysiological Mechanisms and Therapeutic Approaches. Expert Rev. Respir. Med. 2016, 10, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Semenyuk, E.G.; Laning, M.L.; Foley, J.; Johnston, P.F.; Knight, K.L.; Gerding, D.N.; Driks, A. Spore Formation and Toxin Production in Clostridium difficile Biofilms. PLoS ONE 2014, 9, e87757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowther, G.S.; Chilton, C.H.; Todhunter, S.L.; Nicholson, S.; Freeman, J.; Baines, S.D.; Wilcox, M.H. Comparison of Planktonic and Biofilm-Associated Communities of Clostridium difficile and Indigenous Gut Microbiota in a Triple-Stage Chemostat Gut Model. J. Antimicrob. Chemother. 2014, 69, 2137–2147. [Google Scholar] [CrossRef]
- Doan, T.-H.-D.; Yen-Nicolaÿ, S.; Bernet-Camard, M.-F.; Martin-Verstraete, I.; Péchiné, S. Impact of Subinhibitory Concentrations of Metronidazole on Proteome of Clostridioides difficile Strains with Different Levels of Susceptibility. PLoS ONE 2020, 15, e0241903. [Google Scholar] [CrossRef]
- Pantaléon, V.; Soavelomandroso, A.P.; Bouttier, S.; Briandet, R.; Roxas, B.; Chu, M.; Collignon, A.; Janoir, C.; Vedantam, G.; Candela, T. The Clostridium difficile Protease Cwp84 Modulates Both Biofilm Formation and Cell-Surface Properties. PLoS ONE 2015, 10, e0124971. [Google Scholar] [CrossRef]
- Liao, S.; Bitoun, J.; Nguyen, A.; Bozner, D.; Yao, X.; Wen, Z.T. Deficiency of PdxR in Streptococcus mutans Affects Vitamin B6 Metabolism, Acid Tolerance Response and Biofilm Formation. Mol. Oral Microbiol. 2015, 30, 255–268. [Google Scholar] [CrossRef] [Green Version]
- Soavelomandroso, A.P.; Gaudin, F.; Hoys, S.; Nicolas, V.; Vedantam, G.; Janoir, C.; Bouttier, S. Biofilm Structures in a Mono-Associated Mouse Model of Clostridium difficile Infection. Front. Microbiol. 2017, 8, 2086. [Google Scholar] [CrossRef]
- Pantaléon, V.; Monot, M.; Eckert, C.; Hoys, S.; Collignon, A.; Janoir, C.; Candela, T. Clostridium difficile Forms Variable Biofilms on Abiotic Surface. Anaerobe 2018, 53, 34–37. [Google Scholar] [CrossRef]
- Purcell, E.B.; McKee, R.W.; McBride, S.M.; Waters, C.M.; Tamayo, R. Cyclic Diguanylate Inversely Regulates Motility and Aggregation in Clostridium difficile. J. Bacteriol. 2012, 194, 3307–3316. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, D.; Sharif, S.; Weese, J.S. Evaluation of Candidate Reference Genes in Clostridium difficile for Gene Expression Normalization. Anaerobe 2010, 16, 439–443. [Google Scholar] [CrossRef]
- Denève, C.; Deloménie, C.; Barc, M.-C.; Collignon, A.; Janoir, C. Antibiotics Involved in Clostridium difficile-Associated Disease Increase Colonization Factor Gene Expression. J. Med. Microbiol. 2008, 57, 732–738. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Katchar, K.; Goldsmith, J.D.; Nanthakumar, N.; Cheknis, A.; Gerding, D.N.; Kelly, C.P. A Mouse Model of Clostridium difficile—Associated Disease. Gastroenterology 2008, 135, 1984–1992. [Google Scholar] [CrossRef]
- Péchiné, S.; Janoir, C.; Boureau, H.; Gleizes, A.; Tsapis, N.; Hoys, S.; Fattal, E.; Collignon, A. Diminished Intestinal Colonization by Clostridium difficile and Immune Response in Mice after Mucosal Immunization with Surface Proteins of Clostridium Difficile. Vaccine 2007, 25, 3946–3954. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doan, T.-H.-D.; Bernet-Camard, M.-F.; Hoÿs, S.; Janoir, C.; Péchiné, S. Impact of Subinhibitory Concentrations of Metronidazole on Morphology, Motility, Biofilm Formation and Colonization of Clostridioides difficile. Antibiotics 2022, 11, 624. https://doi.org/10.3390/antibiotics11050624
Doan T-H-D, Bernet-Camard M-F, Hoÿs S, Janoir C, Péchiné S. Impact of Subinhibitory Concentrations of Metronidazole on Morphology, Motility, Biofilm Formation and Colonization of Clostridioides difficile. Antibiotics. 2022; 11(5):624. https://doi.org/10.3390/antibiotics11050624
Chicago/Turabian StyleDoan, Tri-Hanh-Dung, Marie-Françoise Bernet-Camard, Sandra Hoÿs, Claire Janoir, and Séverine Péchiné. 2022. "Impact of Subinhibitory Concentrations of Metronidazole on Morphology, Motility, Biofilm Formation and Colonization of Clostridioides difficile" Antibiotics 11, no. 5: 624. https://doi.org/10.3390/antibiotics11050624
APA StyleDoan, T.-H.-D., Bernet-Camard, M.-F., Hoÿs, S., Janoir, C., & Péchiné, S. (2022). Impact of Subinhibitory Concentrations of Metronidazole on Morphology, Motility, Biofilm Formation and Colonization of Clostridioides difficile. Antibiotics, 11(5), 624. https://doi.org/10.3390/antibiotics11050624