
Synergistic Antifungal Activity of Synthetic Peptides and Antifungal Drugs against Candida albicans and C. parapsilosis Biofilms


 ,
,
Abstract
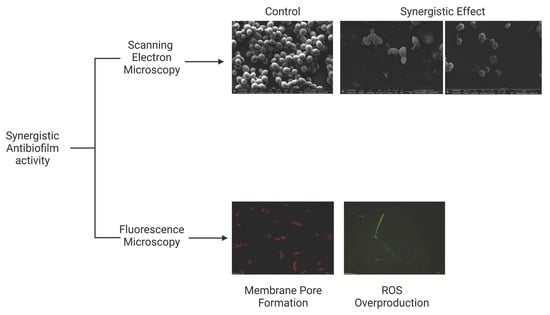
:
1. Introduction
2. Results
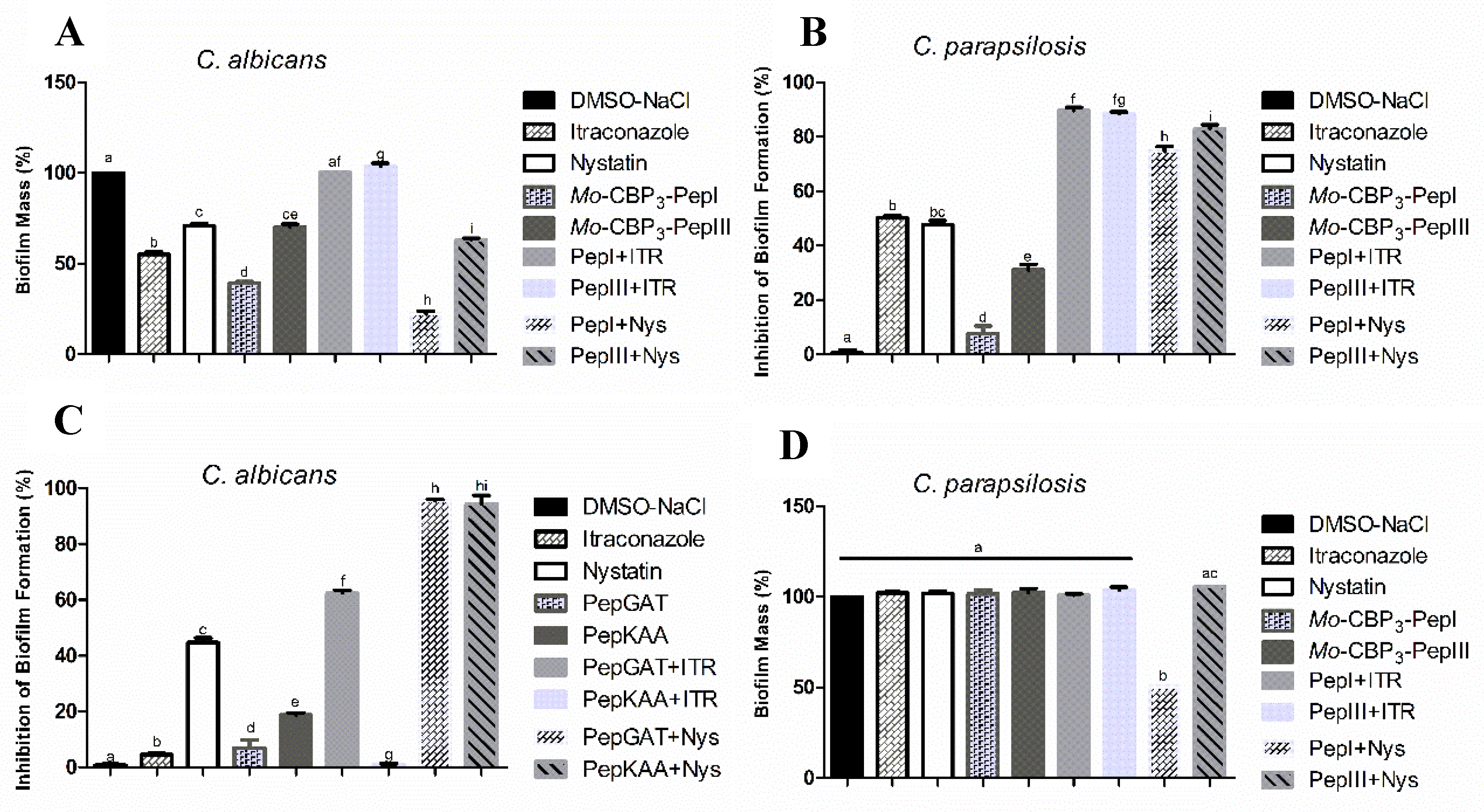
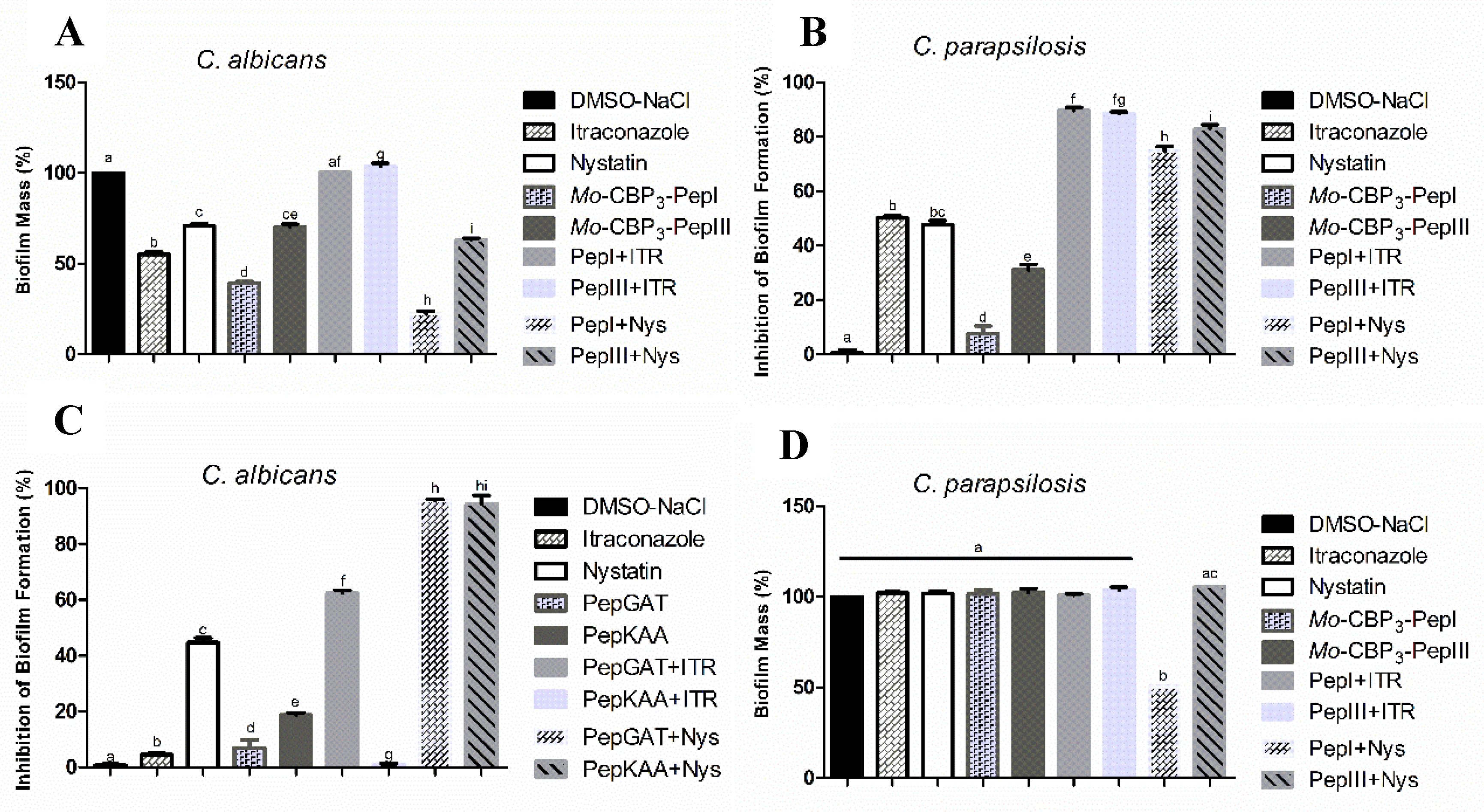
2.1. Antibiofilm Activity of Synthetic Peptides and Two Commercial Drugs
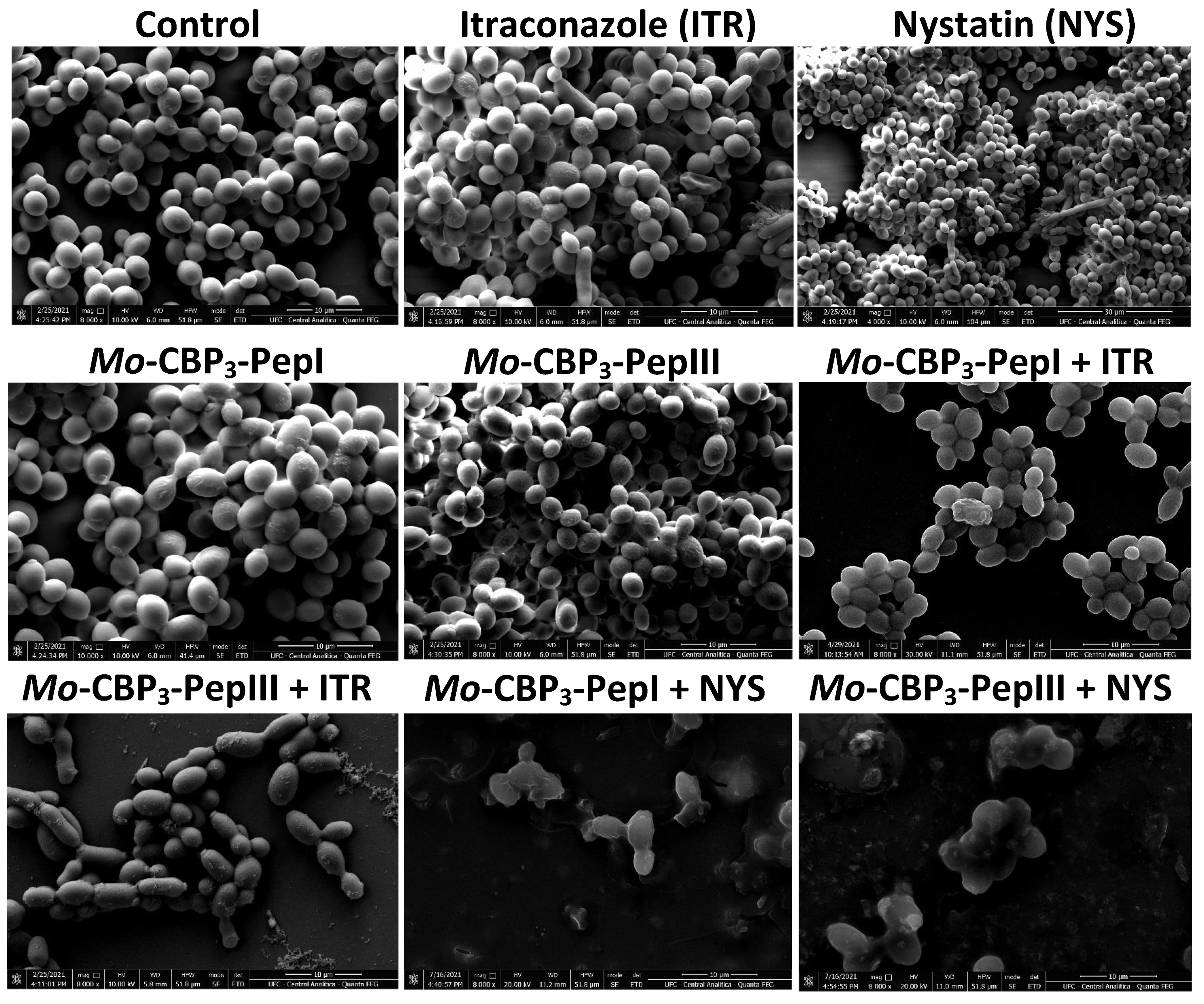
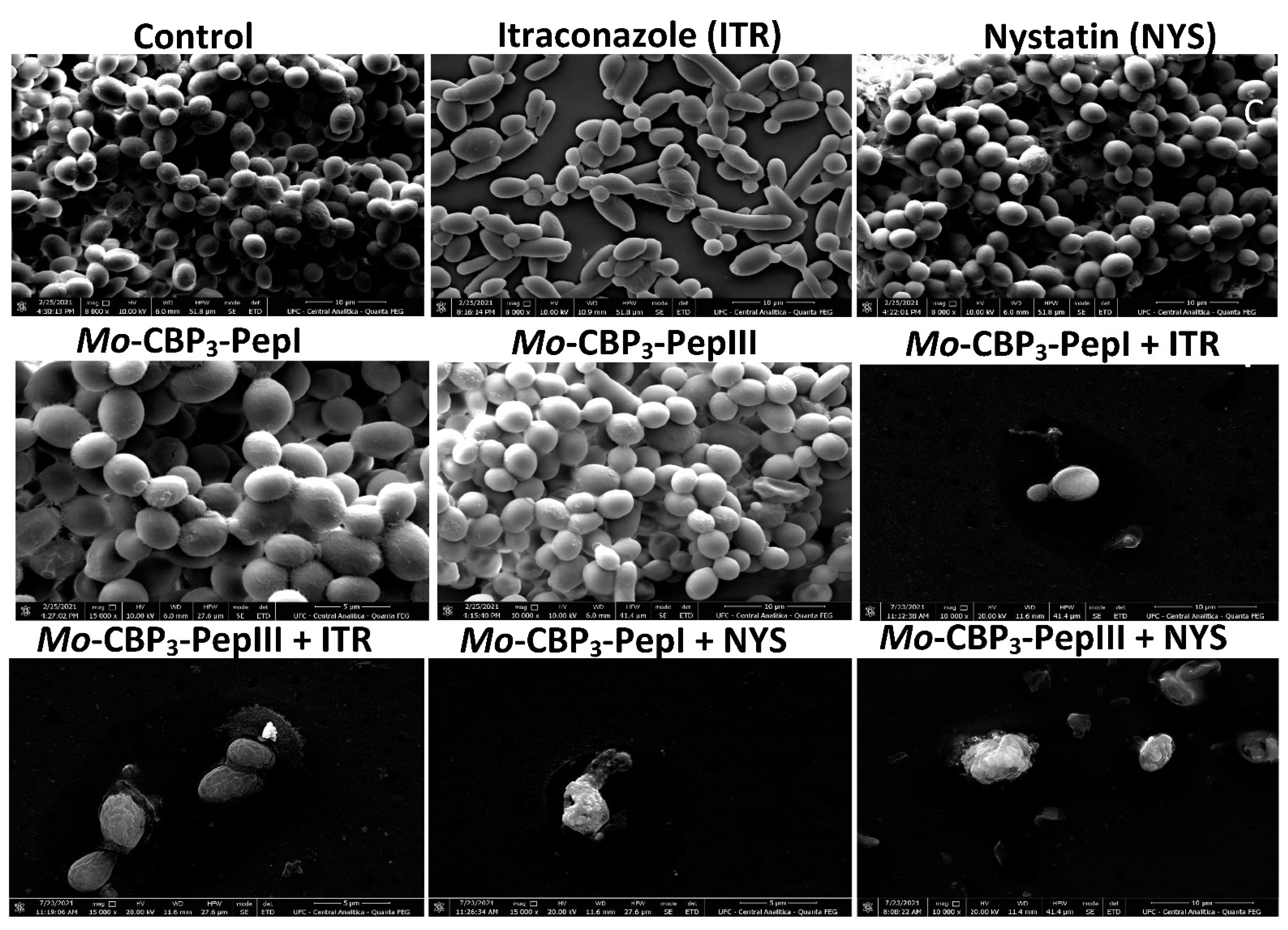
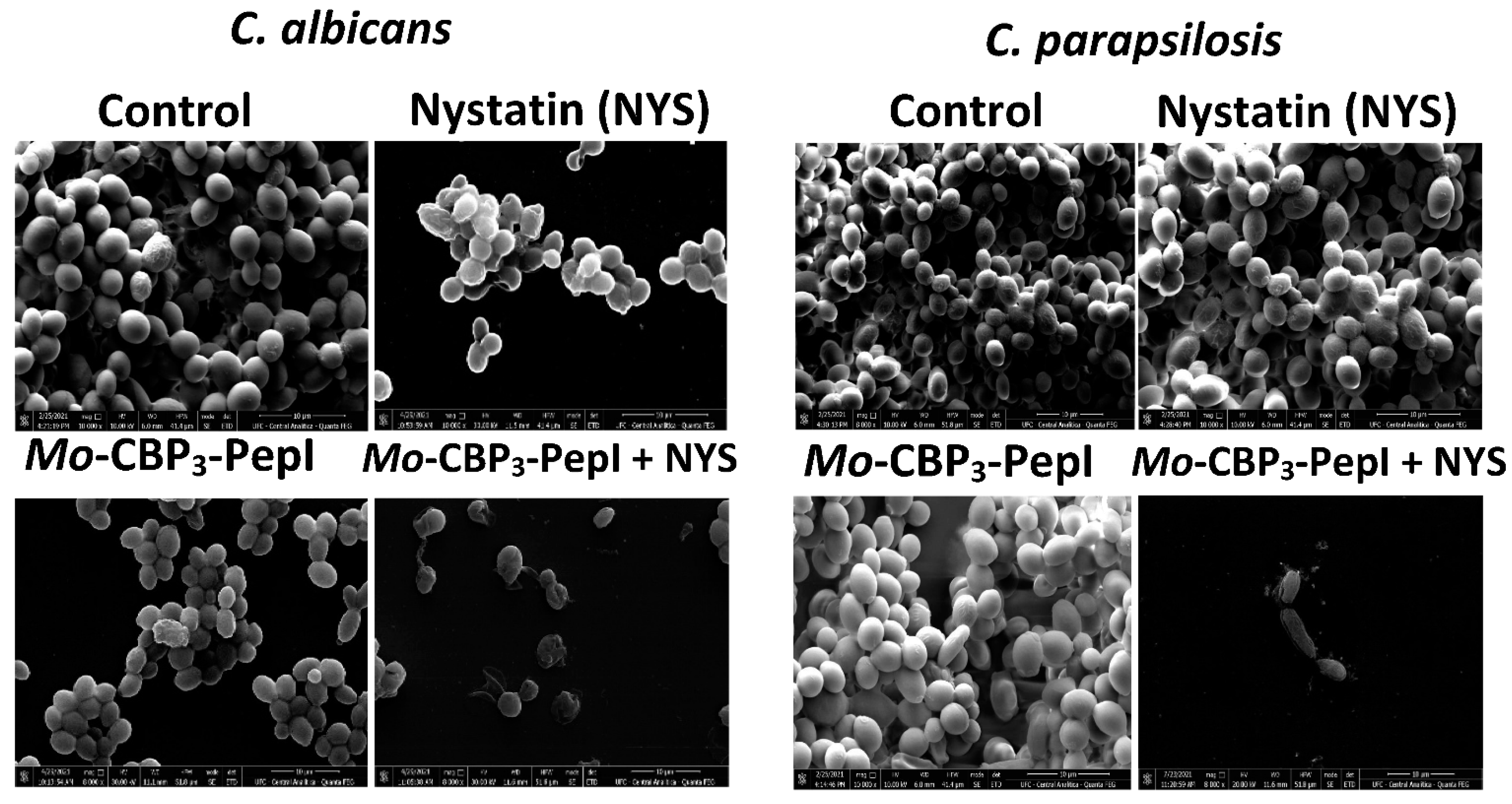
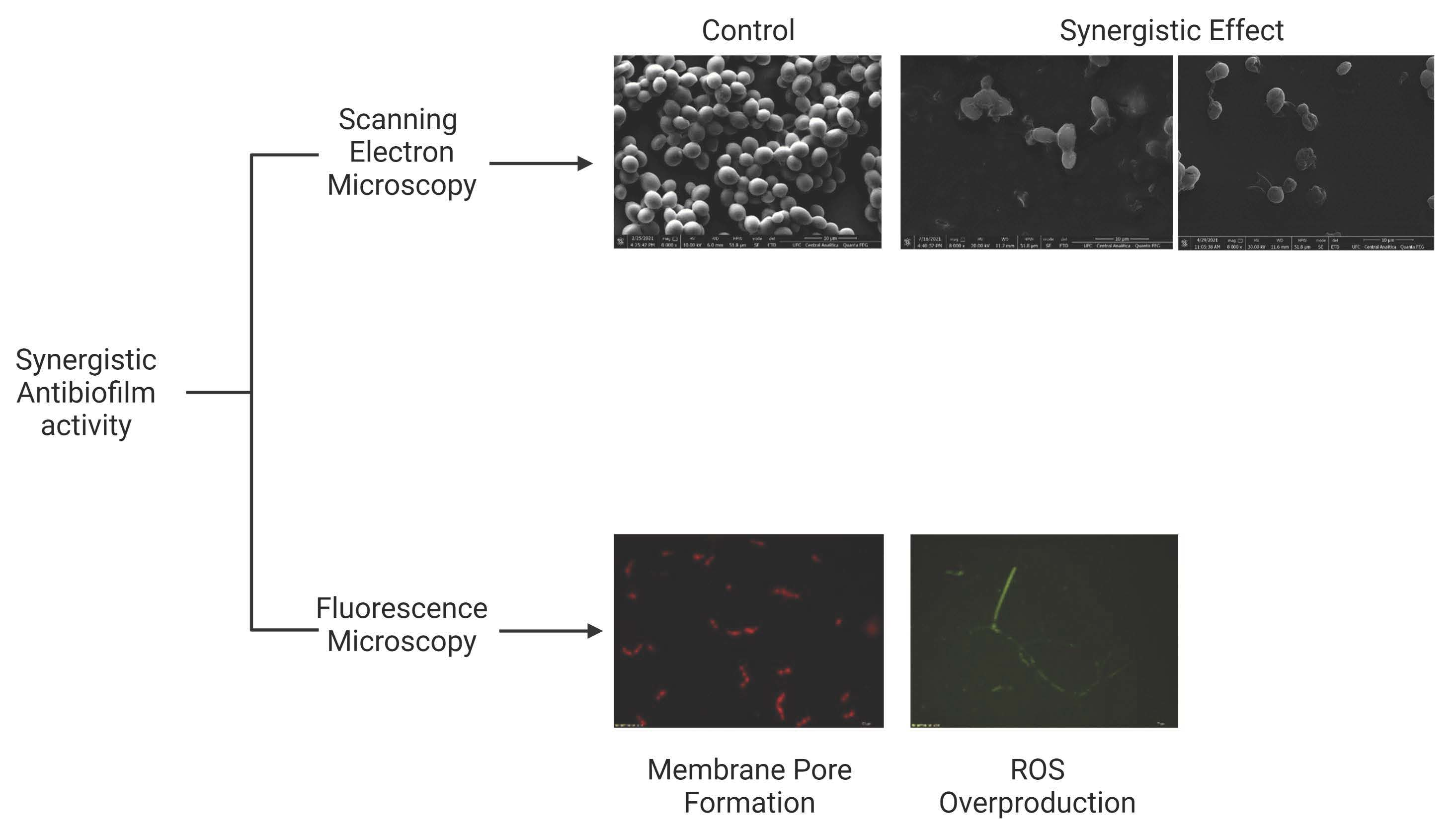
2.2. Analysis of Candida Biofilm Morphology
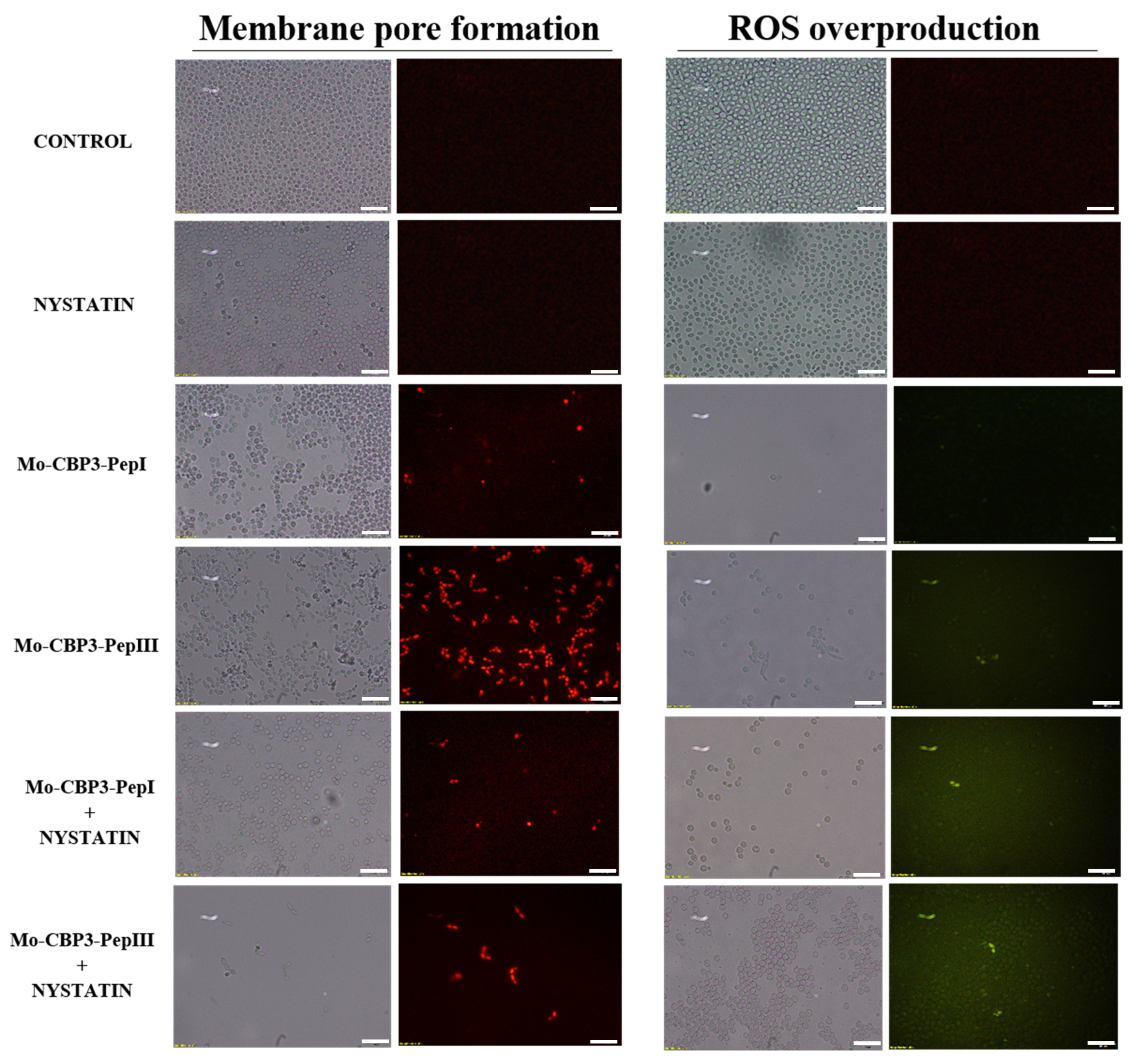
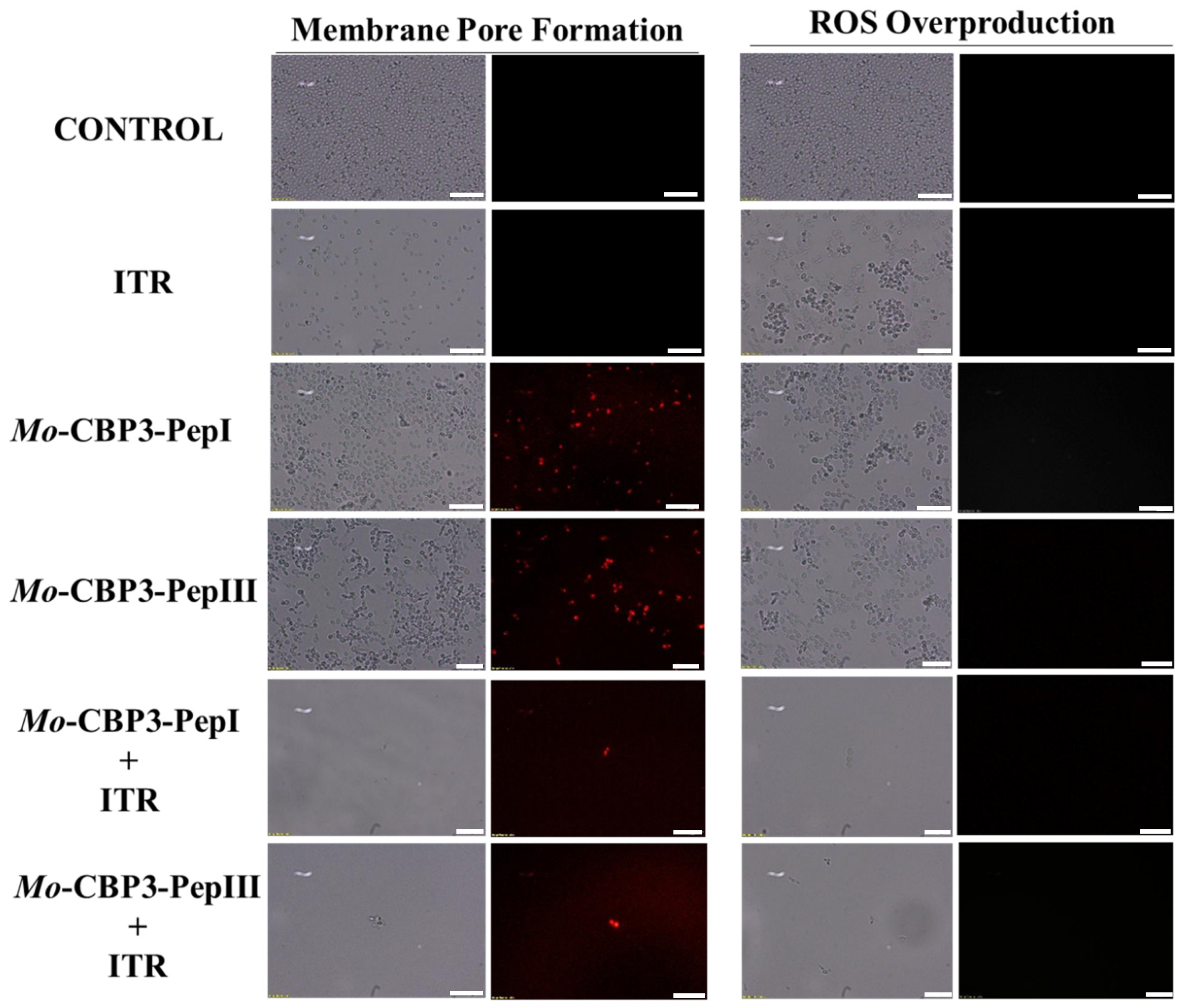
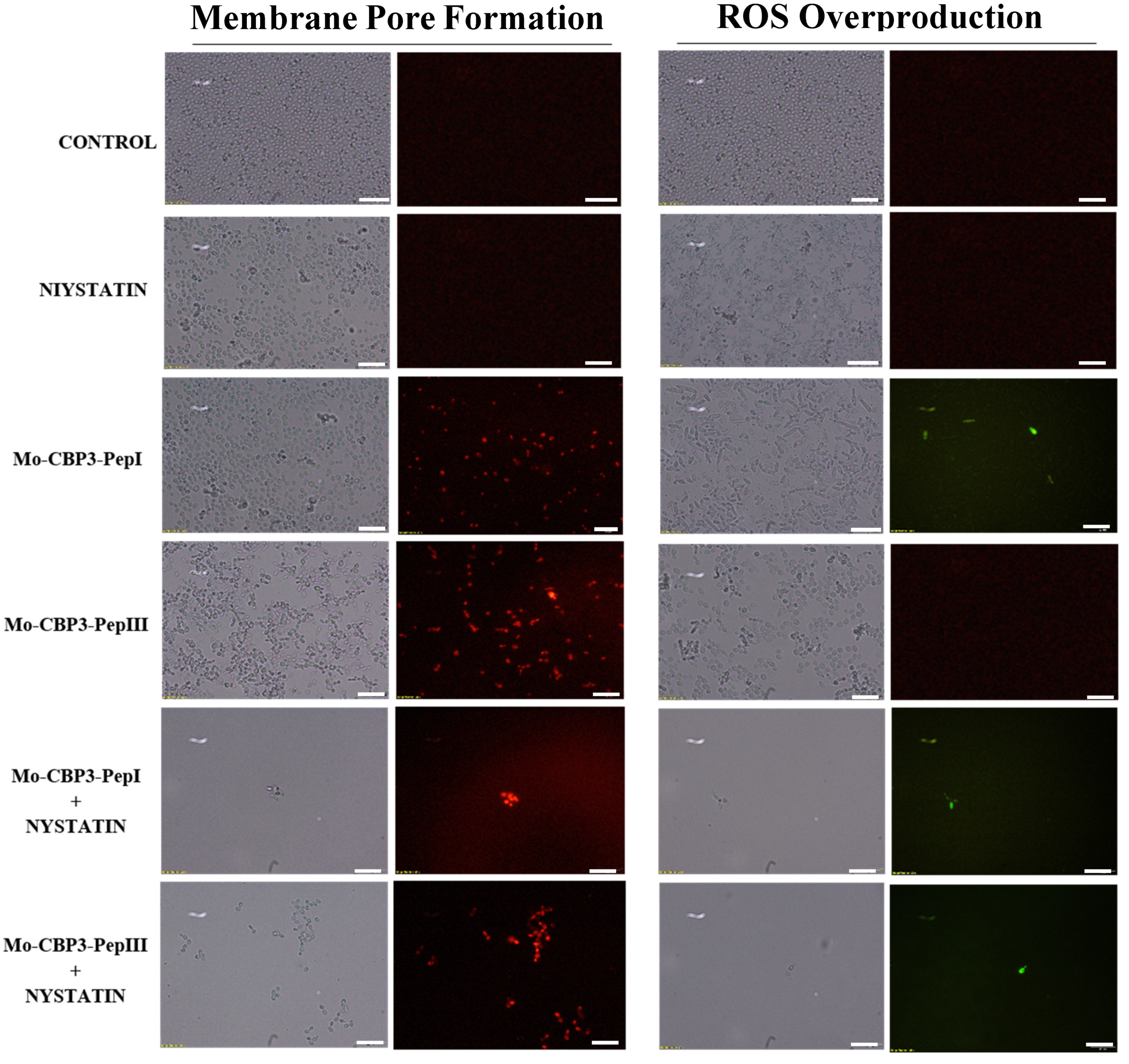
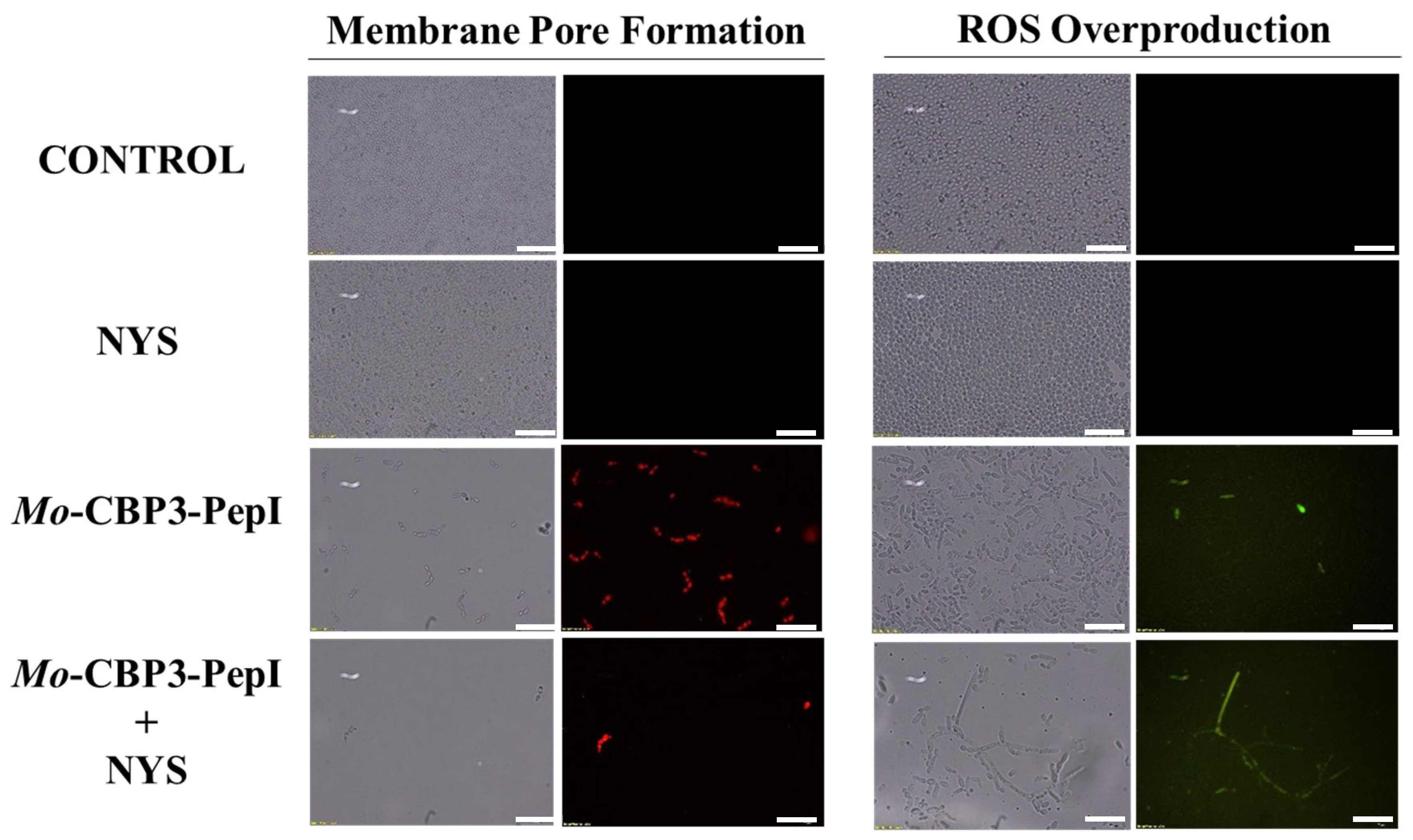
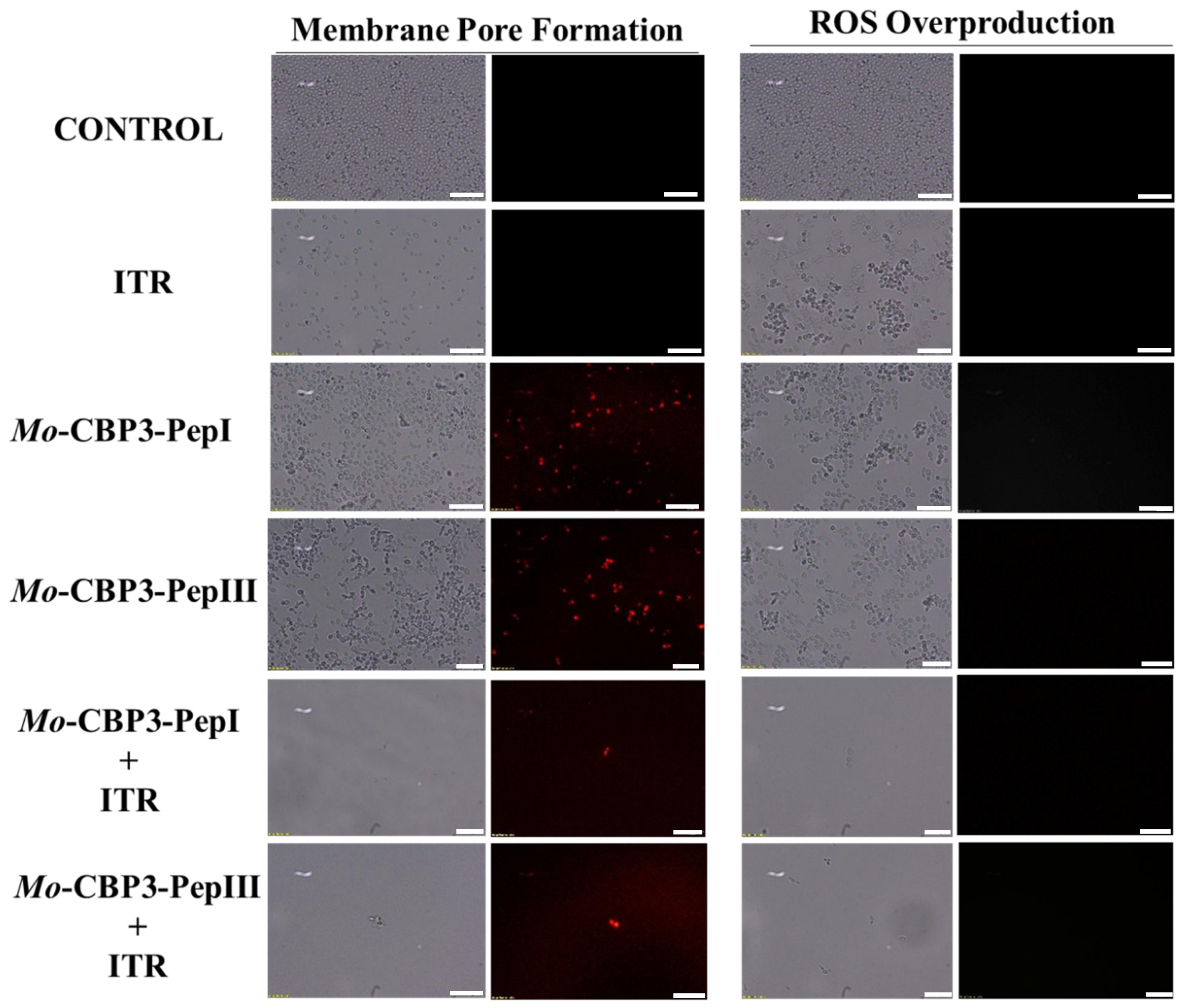
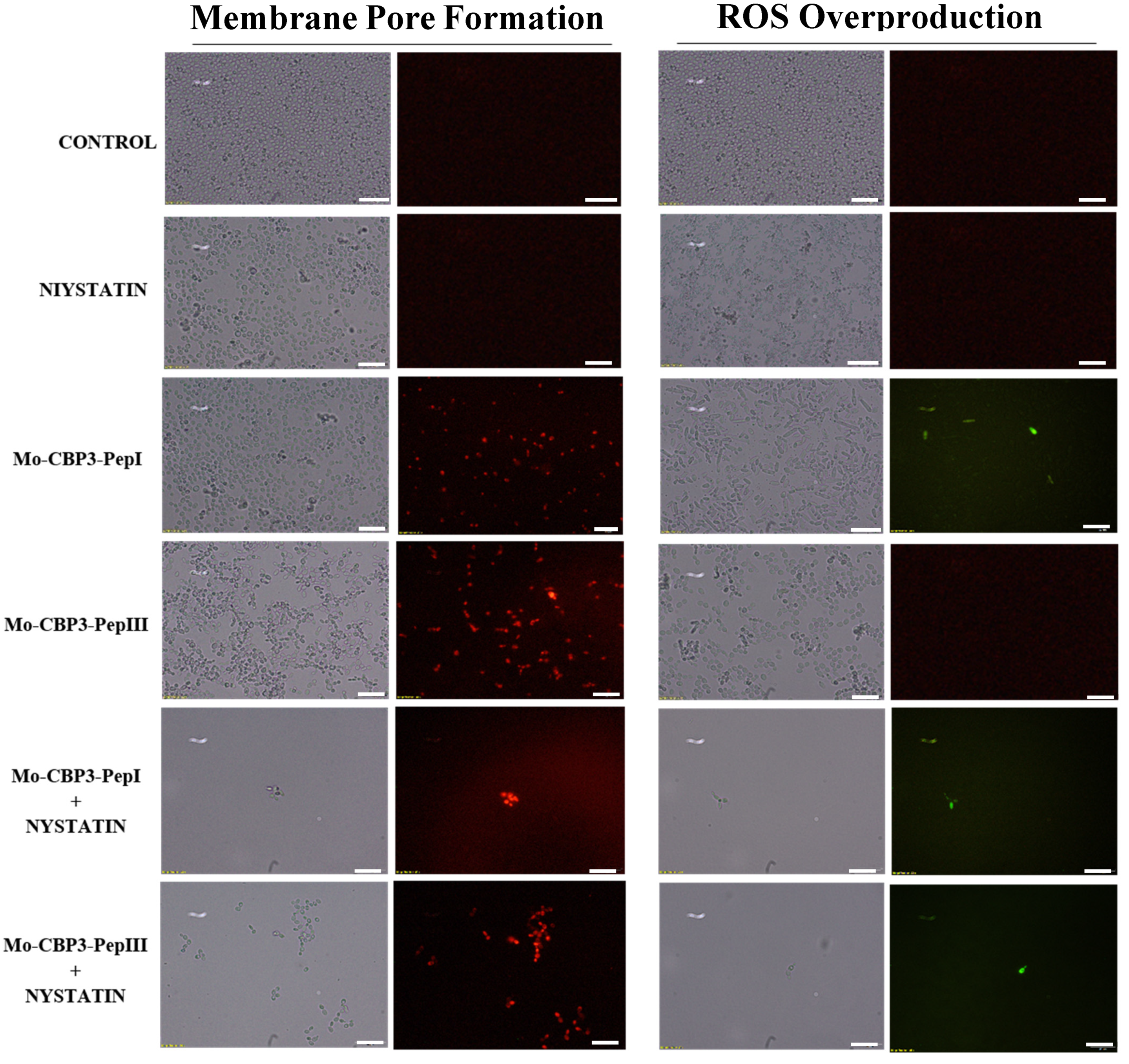
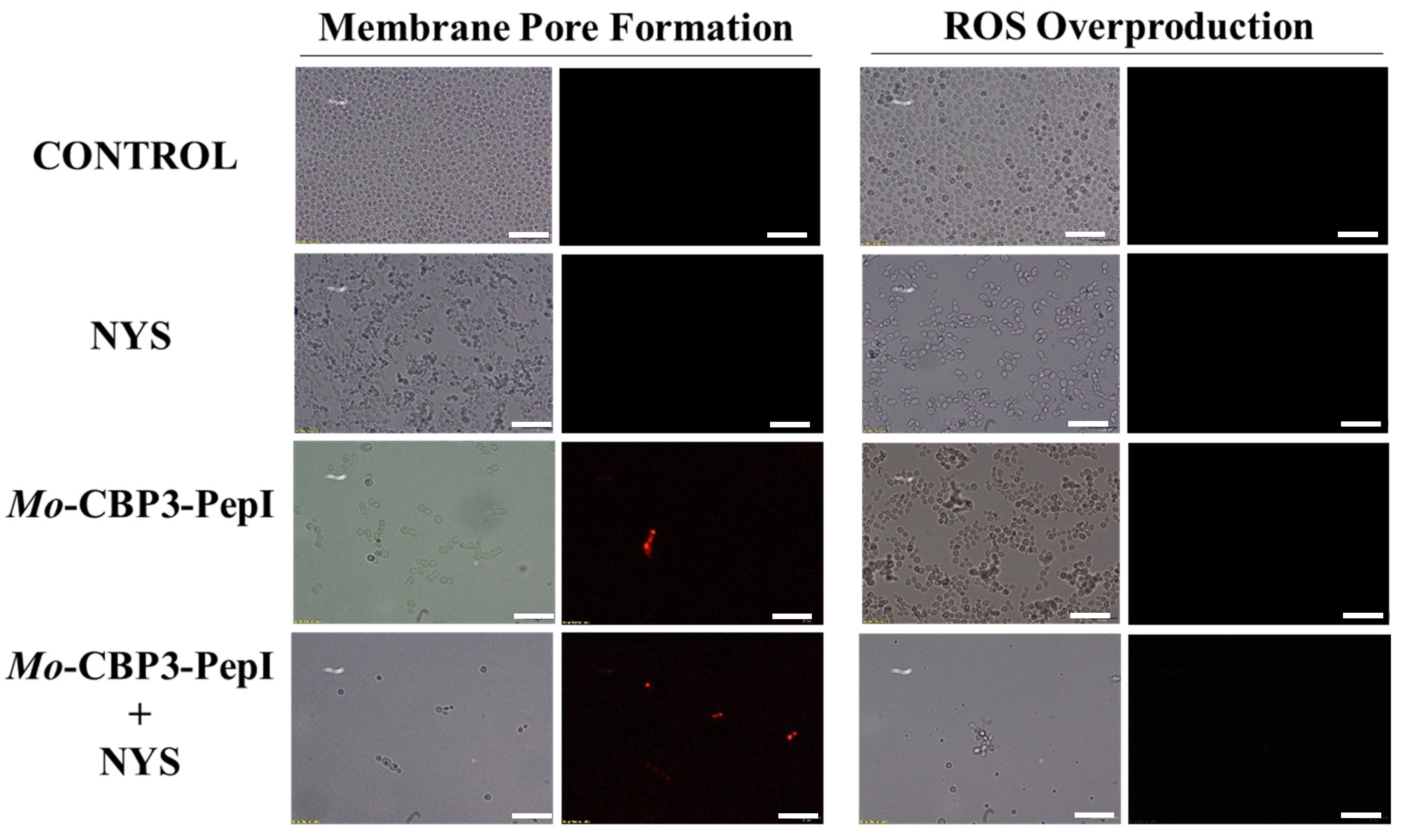
2.3. Membrane Pore Formation
2.4. Molecular Docking
2.5. Hemolytic Assay
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Biological and Chemical Materials
4.3. Peptide Synthesis
4.4. Biological Activity
Antibiofilm Assay
4.5. Overproduction of Reactive Oxygen Species (ROS)
4.6. Cell Membrane Integrity Assay
4.7. Scanning Electron Microscopy (SEM) Analysis
4.8. Obtainment, File Preparation, and Molecular Docking
4.9. Hemolytic Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lima, P.G.; Oliveira, J.T.A.; Amaral, J.L.; Freitas, C.D.T.; Souza, P.F.N. Synthetic antimicrobial peptides: Characteristics, design, and potential as alternative molecules to overcome microbial resistance. Life Sci. 2021, 278, 119647–119660. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Alam, A.; Rani, M.; Ehtesham, N.Z.; Hasnain, S.E. Biofilms: Survival and defense strategy for pathogens. Int. J. Med. Microbiol. 2017, 307, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Kovács, R.; Majoros, L. Fungal quorum-sensing molecules: A review of their antifungal effect against Candida biofilms. J. Fungi 2020, 6, 99. [Google Scholar] [CrossRef] [PubMed]
- Zarnowski, R.; Westler, W.M.; Lacmbouh, G.A.; Marita, J.M.; Bothe, J.R.; Bernhardt, J.; Lounes-Hadj Sahraoui, A.; Fontaine, J.; Sanchez, H.; Hatfield, R.D.; et al. Novel entries in a fungal biofilm matrix encyclopedia. MBio 2014, 5, e01333-14. [Google Scholar] [CrossRef] [Green Version]
- Fox, E.P.; Nobile, C.J. A sticky situation. Transcription 2012, 3, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, B.J.; Oude Lashof, A.M.L. Epidemiology of opportunistic invasive mycoses. Eur. J. Med. Res. 2002, 7, 183–191. [Google Scholar] [PubMed]
- Weig, M. Clinical aspects and pathogenesis of Candida Infection. Trends Microbiol. 1998, 6, 468–470. [Google Scholar] [CrossRef]
- Baillie, G.S. Matrix polymers of Candida biofilms and their possible role in biofilm resistance to antifungal agents. J. Antimicrob. Chemother. 2000, 46, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Sasso, M.; Roger, C.; Sasso, M.; Poujol, H.; Barbar, S.; Lefrant, J.-Y.; Lachaud, L. Changes in the distribution of colonising and infecting Candida Spp. isolates, antifungal drug consumption and susceptibility in a french intensive care unit: A 10-year study. Mycoses 2017, 60, 770–780. [Google Scholar] [CrossRef]
- LaFleur, M.D.; Kumamoto, C.A.; Lewis, K. Candida albicans biofilms produce antifungal-tolerant persister cells. Antimicrob. Agents Chemother. 2006, 50, 3839–3846. [Google Scholar] [CrossRef] [Green Version]
- Ramage, G. Investigation of multidrug efflux pumps in relation to fluconazole resistance in Candida albicans biofilms. J. Antimicrob. Chemother. 2002, 49, 973–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendrup, M.C.; Patterson, T.F. Multidrug-resistant Candida: Epidemiology, molecular mechanisms, and treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katiyar, S.; Pfaller, M.; Edlind, T. Candida albicans and Candida glabrata clinical isolates exhibiting reduced echinocandin susceptibility. Antimicrob. Agents Chemother. 2006, 50, 2892–2894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, P.F.N.; Marques, L.S.M.; Oliveira, J.T.A.; Lima, P.G.; Dias, L.P.; Neto, N.A.S.; Lopes, F.E.S.; Sousa, J.S.; Silva, A.F.B.; Caneiro, R.F.; et al. Synthetic antimicrobial peptides: From choice of the best sequences to action mechanisms. Biochimie 2020, 175, 132–145. [Google Scholar] [CrossRef]
- Lima, P.G.; Souza, P.F.N.; Freitas, C.D.T.; Oliveira, J.T.A.; Dias, L.P.; Neto, J.X.S.; Vasconcelos, I.M.; Lopes, J.L.S.; Sousa, D.O.B. Anticandidal activity of synthetic peptides: Mechanism of action revealed by scanning electron and fluorescence microscopies and synergism effect with nystatin. J. Pept. Sci. 2020, 26, e3249. [Google Scholar] [CrossRef]
- Oliveira, J.T.A.; Souza, P.F.N.; Vasconcelos, I.M.; Dias, L.P.; Martins, T.F.; Van Tilburg, M.F.; Guedes, M.I.F.; Sousa, D.O.B. Mo-CBP3-PepI, Mo-CBP3-PepII, and Mo-CBP3-PepIII are synthetic antimicrobial peptides active against human pathogens by stimulating ROS generation and increasing plasma membrane permeability. Biochimie 2019, 157, 10–21. [Google Scholar] [CrossRef]
- Mason, A.J.; Moussaoui, W.; Abdelrahman, T.; Boukhari, A.; Bertani, P.; Marquette, A.; Shooshtarizaheh, P.; Moulay, G.; Boehm, N.; Guerold, B. Structural determinants of antimicrobial and antiplasmodial activity and selectivity in histidine-rich amphipathic cationic peptides. J. Biol. Chem. 2009, 284, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Mason, A.J.; Gasnier, C.; Kichler, A.; Prévost, G.; Aunis, D.; Metz-Boutigue, M.H.; Bechinger, B. Enhanced membrane disruption and antibiotic action against pathogenic bacteria by designed histidine-rich peptides at acidic PH. Antimicrob. Agents Chemother. 2006, 50, 3305–3311. [Google Scholar] [CrossRef] [Green Version]
- Galdiero, E.; de Alteriis, E.; De Natale, A.; D’Alterio, A.; Siciliano, A.; Guida, M.; Lombardi, L.; Falanga, A.; Galdiero, S. Eradication of Candida albicans persister cell biofilm by the membranotropic peptide GH625. Sci. Rep. 2020, 10, 5780–5791. [Google Scholar] [CrossRef] [Green Version]
- Belmadani, A.; Semlali, A.; Rouabhia, M. Dermaseptin-S1 decreases Candida albicans growth, biofilm formation and the expression of hyphal wall protein 1 and aspartic protease genes. J. Appl. Microbiol. 2018, 125, 72–83. [Google Scholar] [CrossRef]
- Sierra, J.M.; Fusté, E.; Rabanal, F.; Vinuesa, T.; Viñas, M. An overview of antimicrobial peptides and the latest advances in their development. Expert Opin. Biol. Ther. 2017, 17, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Seyedjavadi, S.S.; Khani, S.; Eslamifar, A.; Ajdary, S.; Goudarzi, M.; Halabian, R.; Akbari, R.; Zare-Zardini, H.; Imani Fooladi, A.A.; Amani, J.; et al. The antifungal peptide MCh-AMP1 derived from Matricaria chamomilla inhibits Candida albicans growth via inducing ROS generation and altering fungal cell membrane permeability. Front. Microbiol. 2020, 10, 3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, P.F.N.; Lima, P.G.; Freitas, C.D.T.; Sousa, D.O.B.; Neto, N.A.S.; Dias, L.P.; Vasconcelos, I.M.; Freitas, L.B.N.; Silva, R.G.G.; Sousa, J.S.; et al. Antidermatophytic activity of synthetic peptides: Action mechanisms and clinical application as adjuvants to enhance the activity and decrease the toxicity of griseofulvin. Mycoses 2020, 63, 979–992. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, M.S.A.; Ito, Y.; Nakamura, A.; Tanaka, N.; Fujita, K.; Fukui, H.; Takegawa, K. Nystatin effects on vacuolar function in Saccharomyce. Biosci. Biotechnol. Biochem. 1999, 63, 1075–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgers, M.; Van de Ven, M.-A. Mode of action of itraconazole: Morphological aspects. Mycoses 1989, 32 (Suppl. 1), 53–59. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Huang, J.; Chen, Y. Alpha-helical cationic antimicrobial peptides: Relationships of structure and function. Protein Cell 2010, 1, 143–152. [Google Scholar] [CrossRef]
- Benavent, C.; García-Herrero, V.; Torrado, C.; Torrado-Santiago, S. Nystatin antifungal micellar systems on endotracheal tubes: Development, characterization and in vitro evaluation. Pharmazie 2019, 74, 34–38. [Google Scholar] [CrossRef]
- Lang, B.; Liu, S.; McGinity, J.W.; Williams, R.O. Effect of hydrophilic additives on the dissolution and pharmacokinetic properties of itraconazole-enteric polymer hot-melt extruded amorphous solid dispersions. Drug Dev. Ind. Pharm. 2016, 42, 429–445. [Google Scholar] [CrossRef]
- Dias, L.P.; Souza, P.F.N.; Oliveira, J.T.A.; Vasconcelos, I.M.; Araújo, N.M.S.; Tilburg, M.F.V.; Guedes, M.I.F.; Carneiro, R.F.; Lopes, J.L.S.; Sousa, D.O.B. RcAlb-PepII, a synthetic small peptide bioinspired in the 2S albumin from the seed cake of Ricinus communis, is a potent antimicrobial agent against Klebsiella pneumoniae and Candida parapsilosis. Biochim. Biophys. Acta-Biomembr. 2020, 1862, 183092–183102. [Google Scholar] [CrossRef]
- Staniszewska, M.; Bondaryk, M.; Swoboda-Kopec, E.; Siennicka, K.; Sygitowicz, G.; Kurzatkowski, W. Candida albicans morphologies revealed by scanning electron microscopy analysis. Braz. J. Microbiol. 2013, 44, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Lamiable, A.; Thévenet, P.; Rey, J.; Vavrusa, M.; Derreumaux, P.; Tufféry, P. PEP-FOLD3: Faster de novo structure prediction for linear peptides in solution and in complex. Nucleic Acids Res. 2016, 44, W449–W454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Rosell, G.; Giorgino, T.; De Fabritiis, G. PlayMolecule ProteinPrepare: A web application for protein preparation for molecular dynamics simulations. J. Chem. Inf. Model. 2017, 57, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and autodocktools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides/Combinations | % Hemolysis | ||
|---|---|---|---|
| Type A Blood | Type B Blood | Type O Blood | |
| 0.1% Triton X-100 | 100 ± 0.002 | 100 ± 0.001 | 100 ± 0.007 |
| DMSO-NaCl Solution | 0 | 0 | 0 |
| NYS (1000 µg mL−1) | 100 ± 0.005 | 100 ± 0.001 | 100 ± 0.002 |
| ITR (1000 µg mL−1) | 75 ± 0.007 | 68 ± 0.004 | 58 ± 0.003 |
| Mo-CBP3-PepI (50 µg mL−1) | 0 | 0 | 0 |
| Mo-CBP3-PepIII (50 µg mL−1) | 0 | 0 | 0 |
| Mo-CBP3-PepI (50 µg mL−1) + NYS (1000 µg mL−1) | 14 ± 0.006 | 23 ± 0.009 | 2 ± 0.001 |
| Mo-CBP3-PepI (50 µg mL−1) + ITR (1000 µg mL−1) | 0 | 4 ± 0.003 | 8 ± 0.005 |
| Mo-CBP3-PepIII (50 µg mL−1) + NYS (1000 µg mL−1) | 45 ± 0.001 | 30 ± 0.001 | 18 ± 0.007 |
| Mo-CBP3-PepIII (50 µg mL−1) + ITR (1000 µg mL−1) | 50 ± 0.005 | 15 ± 0.008 | 2 ± 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezerra, L.P.; Freitas, C.D.T.; Silva, A.F.B.; Amaral, J.L.; Neto, N.A.S.; Silva, R.G.G.; Parra, A.L.C.; Goldman, G.H.; Oliveira, J.T.A.; Mesquita, F.P.; et al. Synergistic Antifungal Activity of Synthetic Peptides and Antifungal Drugs against Candida albicans and C. parapsilosis Biofilms. Antibiotics 2022, 11, 553. https://doi.org/10.3390/antibiotics11050553
Bezerra LP, Freitas CDT, Silva AFB, Amaral JL, Neto NAS, Silva RGG, Parra ALC, Goldman GH, Oliveira JTA, Mesquita FP, et al. Synergistic Antifungal Activity of Synthetic Peptides and Antifungal Drugs against Candida albicans and C. parapsilosis Biofilms. Antibiotics. 2022; 11(5):553. https://doi.org/10.3390/antibiotics11050553
Chicago/Turabian StyleBezerra, Leandro P., Cleverson D. T. Freitas, Ayrles F. B. Silva, Jackson L. Amaral, Nilton A. S. Neto, Rafael G. G. Silva, Aura L. C. Parra, Gustavo H. Goldman, Jose T. A. Oliveira, Felipe P. Mesquita, and et al. 2022. "Synergistic Antifungal Activity of Synthetic Peptides and Antifungal Drugs against Candida albicans and C. parapsilosis Biofilms" Antibiotics 11, no. 5: 553. https://doi.org/10.3390/antibiotics11050553
APA StyleBezerra, L. P., Freitas, C. D. T., Silva, A. F. B., Amaral, J. L., Neto, N. A. S., Silva, R. G. G., Parra, A. L. C., Goldman, G. H., Oliveira, J. T. A., Mesquita, F. P., & Souza, P. F. N. (2022). Synergistic Antifungal Activity of Synthetic Peptides and Antifungal Drugs against Candida albicans and C. parapsilosis Biofilms. Antibiotics, 11(5), 553. https://doi.org/10.3390/antibiotics11050553