
Iron Effects on Clostridioides difficile Toxin Production and Antimicrobial Susceptibilities


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
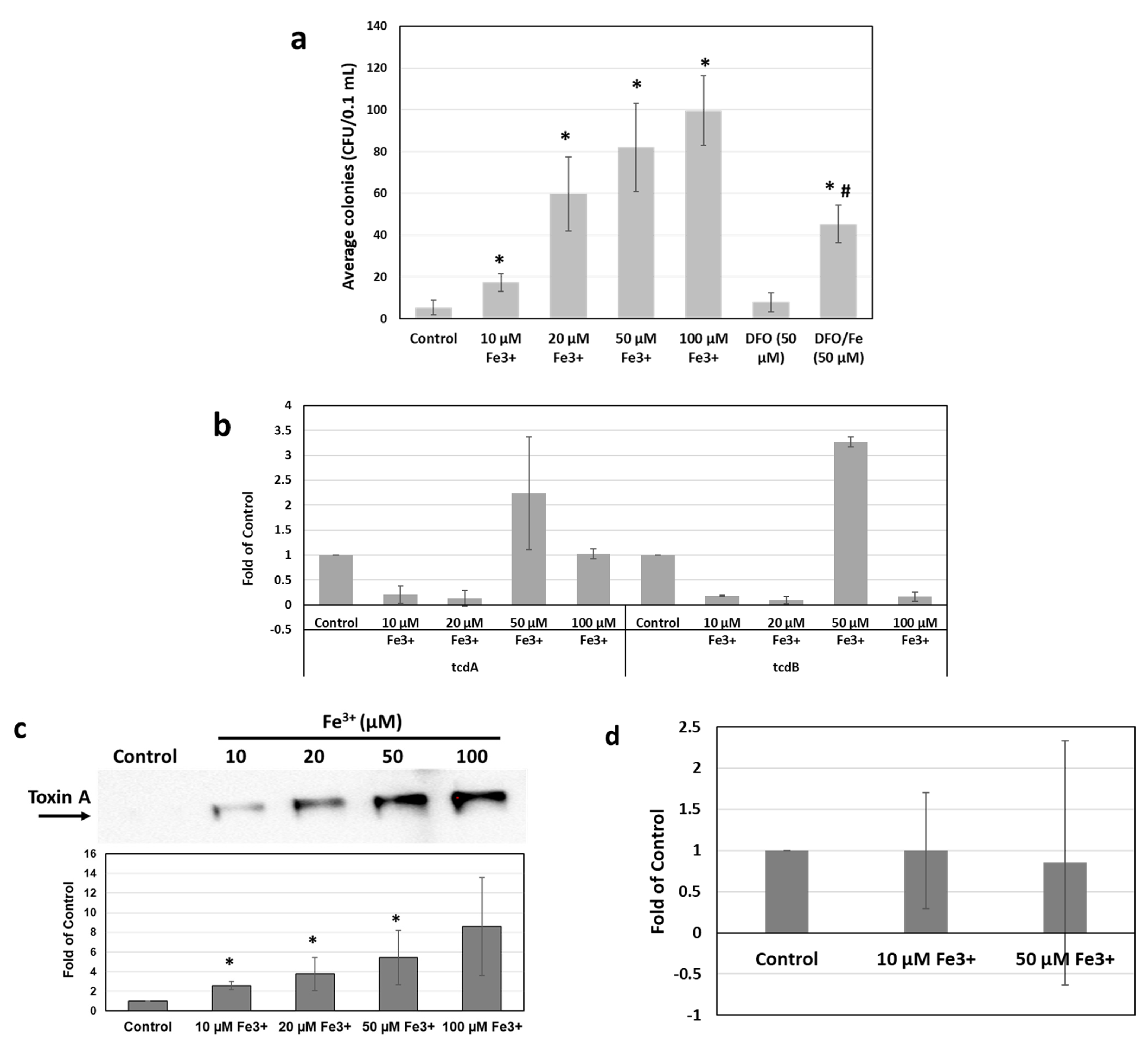
2.1. Iron Stimulates C. difficile Growth and Toxin Production In Vitro
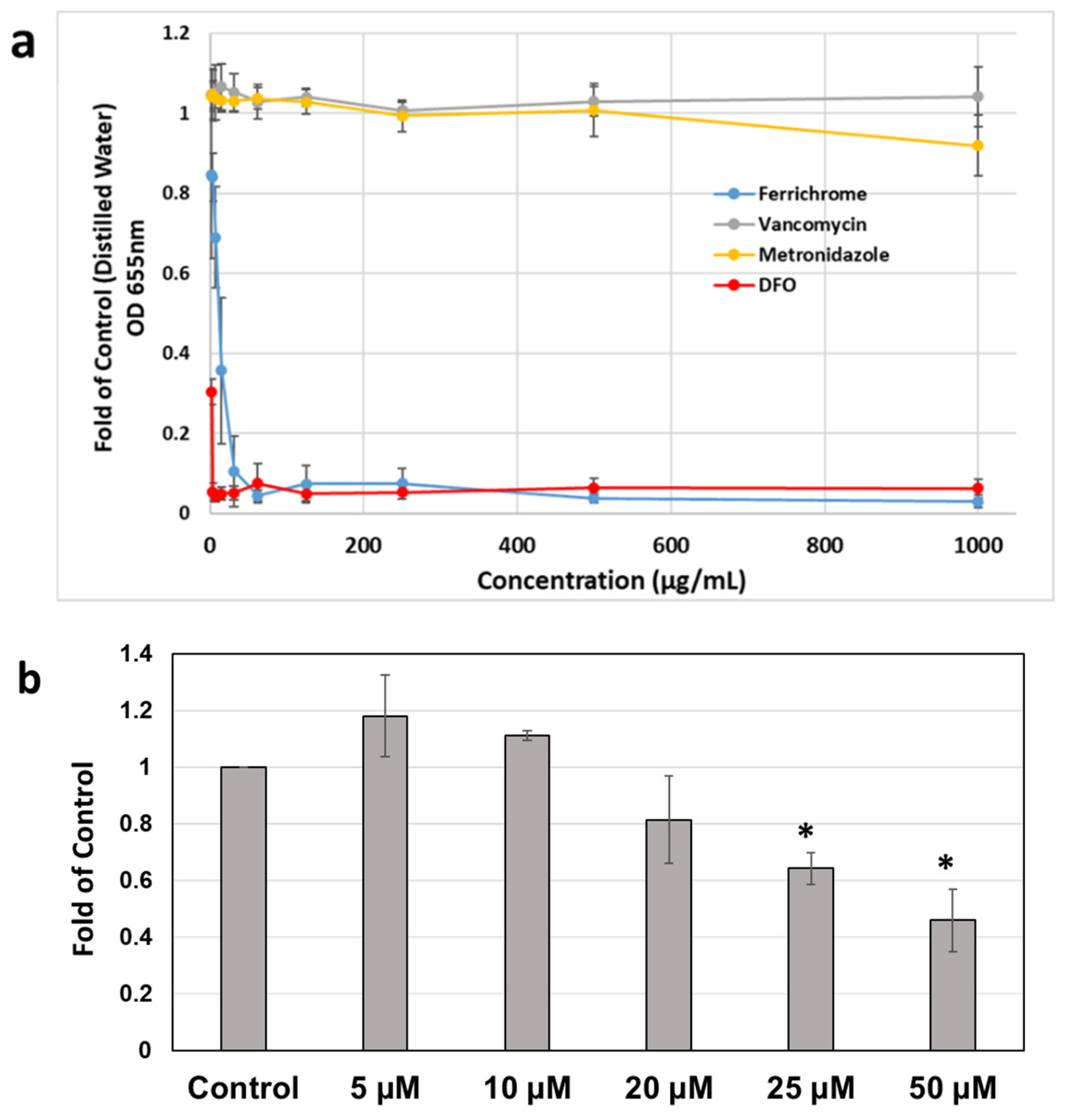
2.2. Effects of Iron Chelation on Metronidazole MIC Profiles of C. difficile Strains
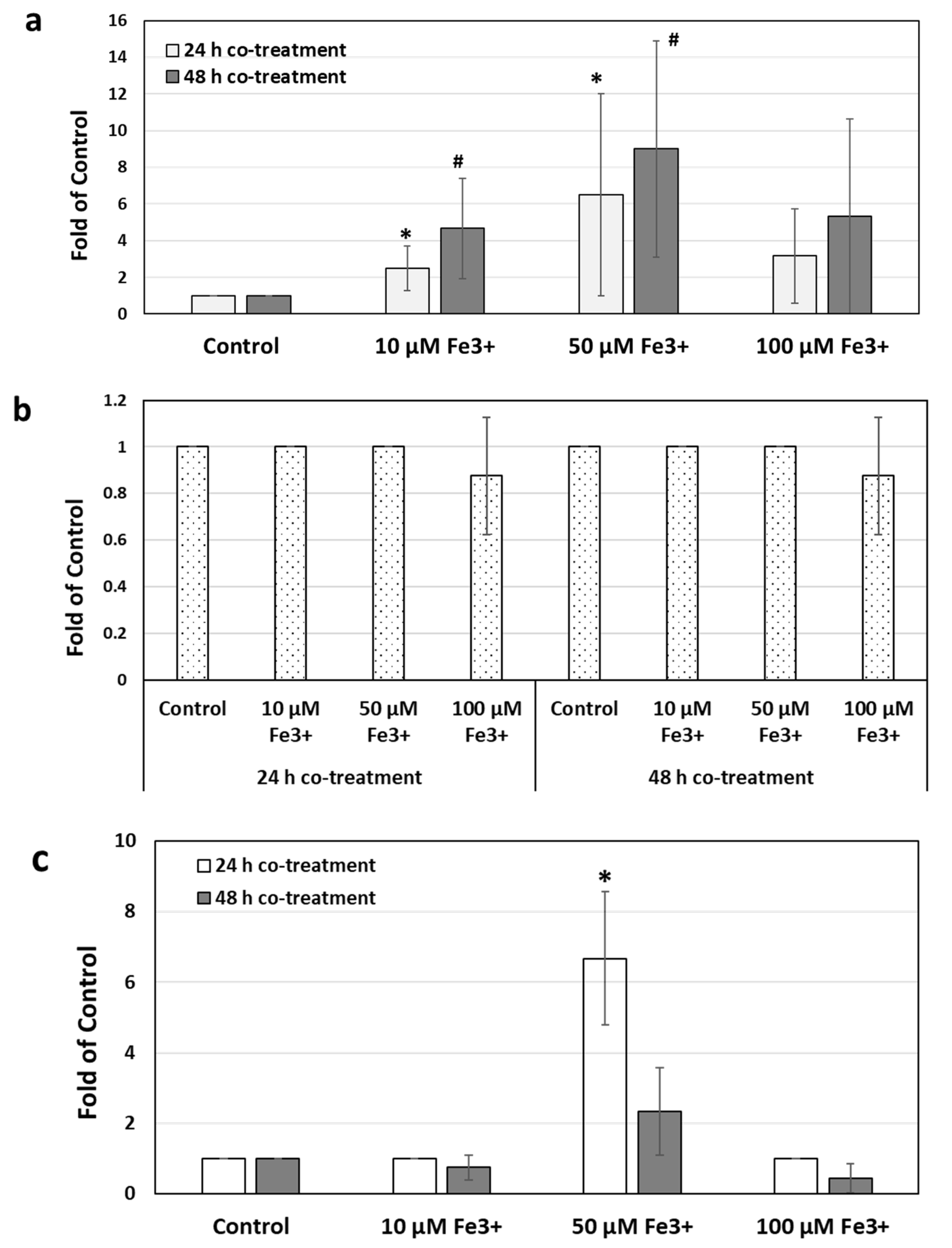
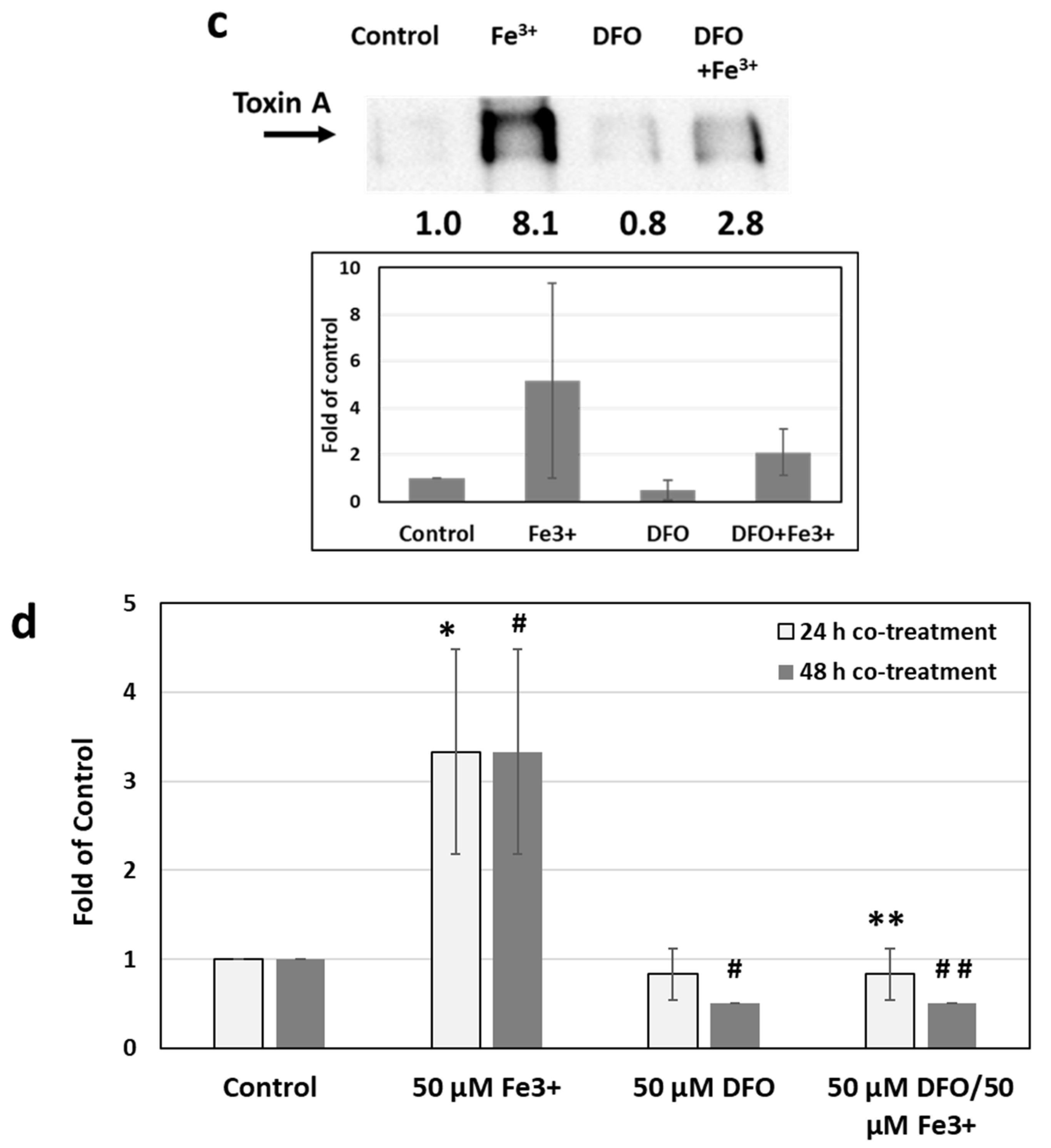
2.3. Effects of Iron Chelation on Iron-Induced Toxin Production and Metronidazole Resistance
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Culture Conditions, and Growth Analysis
4.2. Antibiotics, Iron-Chelators and Iron Supplementation
4.3. Susceptibility Testing
4.4. Polyacrylamide Gel Electrophoresis (PAGE) and Western Blot Analysis
4.5. Chrome Azurol S (CAS) Assay
4.6. Isolation of RNA and qRT-PCR Assay
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lessa, F.C.; Mu, Y.; Bamberg, W.M.; Beldavs, Z.G.; Dumyati, G.K.; Dunn, J.R.; Farley, M.M.; Holzbauer, S.M.; Meek, J.I.; Phipps, E.C.; et al. Burden of Clostridium difficile Infection in the United States. N. Engl. J. Med. 2015, 372, 825–834. [Google Scholar] [CrossRef] [PubMed]
- See, I.; Mu, Y.; Cohen, J.; Beldavs, Z.G.; Winston, L.G.; Dumyati, G.; Holzbauer, S.; Dunn, J.; Farley, M.M.; Lyons, C.; et al. NAP1 strain type predicts outcomes from Clostridium difficile infection. Clin. Infect. Dis. 2014, 58, 1394–1400. [Google Scholar] [CrossRef] [PubMed]
- Curry, S.R.; Marsh, J.W.; Muto, C.A.; O’Leary, M.M.; Pasculle, A.W.; Harrison, L.H. tcdC genotypes associated with severe TcdC truncation in an epidemic clone and other strains of Clostridium difficile. J. Clin. Microbiol. 2007, 45, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Warny, M.; Pepin, J.; Fang, A.; Killgore, G.; Thompson, A.; Brazier, J.; Frost, E.; McDonald, L.C. Toxin production by an emerging strain of Clostridium difficile associated with outbreaks of severe disease in North America and Europe. Lancet 2005, 366, 1079–1084. [Google Scholar] [CrossRef]
- Hebbard, A.I.; Slavin, M.; Reed, C.; Trubiano, J.; Teh, B.; Haeusler, G.; Thursky, K.; Worth, L.J. Risks factors and outcomes of Clostridium difficile infection in patients with cancer: A matched case-control study. Support. Care Cancer 2017, 25, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; Coffin, S.E.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical Practice Guidelines for Clostridium difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, e1–e48. [Google Scholar] [CrossRef] [PubMed]
- Krishna, S.G.; Zhao, W.; Apewokin, S.K.; Krishna, K.; Chepyala, P.; Anaissie, E.J. Risk factors, preemptive therapy, and antiperistaltic agents for Clostridium difficile infection in cancer patients. Transpl. Infect. Dis. 2013, 15, 493–501. [Google Scholar]
- Shah, D.; Dang, M.D.; Hasbun, R.; Koo, H.L.; Jiang, Z.D.; DuPont, H.L.; Garey, K.W. Clostridium difficile infection: Update on emerging antibiotic treatment options and antibiotic resistance. Expert Rev. Anti-Infect. Ther. 2010, 8, 555–564. [Google Scholar] [CrossRef]
- Schmid, D.; Kuo, H.; Simons, E.; Kanitz, E.; Wenisch, J.; Allerberger, F.; Wenisch, C. All-cause mortality in hospitalized patients with infectious diarrhea: Clostridium difficile versus other enteric pathogens in Austria from 2008 to 2010. J. Infect. Public Health 2014, 7, 133–144. [Google Scholar] [CrossRef]
- Sammons, J.S.; Localio, R.; Xiao, R.; Coffin, S.E.; Zaoutis, T. Clostridium difficile Infection Is Associated with Increased Risk of Death and Prolonged Hospitalization in Children. Clin. Infect. Dis. 2013, 57, 1–8. [Google Scholar] [CrossRef]
- Sammons, J.S.; Toltzis, P.; Zaoutis, T.E. Clostridium difficile Infection in children. JAMA Pediatr. 2013, 167, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Lynch, T.; Chong, P.; Zhang, J.; Hizon, R.; Du, T.; Graham, M.R.; Beniac, D.R.; Booth, T.F.; Kibsey, P.; Miller, M.; et al. Characterization of a Stable, Metronidazole-Resistant Clostridium difficile Clinical Isolate. PLoS ONE 2013, 8, e53757. [Google Scholar] [CrossRef] [PubMed]
- Abbaspour, N.; Hurrell, R.; Kelishadi, R. Review on iron and its importance for human health. J. Res. Med. Sci. 2014, 19, 164–174. [Google Scholar] [PubMed]
- Fung, E.B.; Harmatz, P.R.; Milet, M.; Balasa, V.; Ballas, S.K.; Casella, J.F.; Hilliard, L.; Kutlar, A.; McClain, K.; Olivieri, N.F.; et al. Disparity in the management of iron overload between patients with sickle cell disease and thalassemia who received transfusions. Transfusion 2008, 48, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Ault, P.; Jones, K. Understanding iron overload: Screening, monitoring, and caring for patients with transfusion-dependent anemias. Clin. J. Oncol. Nurs. 2009, 13, 511–517. [Google Scholar] [CrossRef]
- Hoffbrand, A.V.; Taher, A.; Cappellini, M.D. How I treat transfusional iron overload. Blood 2012, 120, 3657–3669. [Google Scholar] [CrossRef][Green Version]
- Shander, A.; Sazama, K. Clinical consequences of iron overload from chronic red blood cell transfusions, its diagnosis, and its management by chelation therapy. Transfusion 2010, 50, 1144–1155. [Google Scholar] [CrossRef]
- Murray, M.J.; Murray, A.B.; Murray, M.B.; Murray, C.J. The adverse effect of iron repletion on the course of certain infections. BMJ 1978, 2, 1113–1115. [Google Scholar] [CrossRef]
- Weinberg, E.D. Development of Clinical Methods of Iron Deprivation for Suppression of Neoplastic and Infectious Diseases. Cancer Investig. 1999, 17, 507–513. [Google Scholar] [CrossRef]
- Nairz, M.; Schroll, A.; Sonnweber, T.; Weiss, G. The struggle for iron-a metal at the host-pathogen interface. Cell. Microbiol. 2010, 12, 1691–1702. [Google Scholar] [CrossRef]
- Schaible, M.E.; Kaufmann, S.H.E. Iron and microbial infection. Nat. Rev. Microbiol. 2004, 2, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Pishchany, G.; McCoy, A.L.; Torres, V.; Krause, J.C.; Crowe, J.E.; Fabry, M.E.; Skaar, E.P. Specificity for Human Hemoglobin Enhances Staphylococcus aureus Infection. Cell Host Microbe 2010, 8, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Lowy, F.D. HowStaphylococcus aureusAdapts to Its Host. N. Engl. J. Med. 2011, 364, 1987–1990. [Google Scholar] [CrossRef] [PubMed]
- Bairwa, G.; Jung, W.H.; Kronstad, J.W. Iron acquisition in fungal pathogens of humans. Metallomics 2017, 9, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Iron and infection. Int. J. Hematol. 2018, 107, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Sivgin, S.; Baldane, S.; Kaynar, L.; Kurnaz, F.; Pala, C.; Sivgin, H.; Ozturk, A.; Cetin, M.; Unal, A.; Eser, B. Pretransplant serum ferritin level may be a predictive marker for outcomes in patients having undergone allogeneic hematopoietic stem cell transplantation. Neoplasma 2012, 59, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Xhaard, A.; Lanternier, F.; Porcher, R.; Dannaoui, E.; Bergeron, A.; Clement, L.; Lacroix, C.; Herbrecht, R.; Legrand, F.; Mohty, M.; et al. Mucormycosis after allogeneic haematopoietic stem cell transplantation: A French Multicentre Cohort Study (2003–2008). Clin. Microbiol. Infect. 2012, 18, E396–E400. [Google Scholar] [CrossRef]
- Aldape, M.J.; Heeney, D.D.; Bryant, A.E.; Stevens, D.L. Tigecycline suppresses toxin A and B production and sporulation in Clostridium difficile. J. Antimicrob. Chemother. 2014, 70, 153–159. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Principles and Procedures for Dection of Anaerobes in Clinical Specimen, Approved Guideline; CLSI document M56-A; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2014. [Google Scholar]
- Freeman, J.; Vernon, J.; Vickers, R.; Wilcox, M.H. Susceptibility of Clostridium difficile Isolates of Varying Antimicrobial Resistance Phenotypes to SMT19969 and 11 Comparators. Antimicrob. Agents Chemother. 2016, 60, 689–692. [Google Scholar] [CrossRef]
- Peng, Z.; Addisu, A.; Alrabaa, S.; Sun, X. Antibiotic Resistance and Toxin Production of Clostridium difficile Isolates from the Hospitalized Patients in a Large Hospital in Florida. Front. Microbiol. 2017, 8, 2584. [Google Scholar] [CrossRef]
- Grenier, D.; Huot, M.-P.; Mayrand, D. Iron-Chelating Activity of Tetracyclines and Its Impact on the Susceptibility of Actinobacillus actinomycetemcomitans to These Antibiotics. Antimicrob. Agents Chemother. 2000, 44, 763–766. [Google Scholar] [CrossRef] [PubMed]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- National Comprehensive Cancer Network. NCCN Clinical Practice Guidelines in Oncology: Hematopoietic Growth Factors. Version 1. 2022. 22 December 2021. Available online: https://www.nccn.org/professionals/physician_gls/pdf/growthfactors.pdf (accessed on 18 April 2022).
- Troxell, B.; Hassan, H.M. Transcriptional regulation by Ferric Uptake Regulator (Fur) in pathogenic bacteria. Front. Cell. Infect. Microbiol. 2013, 3, 59. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.D.; Ellermeier, C.D. Ferric Uptake Regulator Fur Control of Putative Iron Acquisition Systems in Clostridium difficile. J. Bacteriol. 2015, 197, 2930–2940. [Google Scholar] [CrossRef]
- Hastie, J.L.; Hanna, P.C.; Carlson, P.E. Transcriptional response of Clostridium difficile to low iron conditions. Pathog. Dis. 2018, 76, fty009. [Google Scholar] [CrossRef]
- Matthaiou, E.I.; Sass, G.; Stevens, D.A.; Hsu, J.L. Iron: An essential nutrient for Aspergillus fumigatus and a fulcrum for pathogenesis. Curr. Opin. Infect. Dis. 2018, 31, 506–511. [Google Scholar] [CrossRef]
- Chan, G.C.; Chan, S.; Ho, P.L.; Ha, S.Y. Effects of chelators (deferoxamine, deferiprone and deferasirox) on the growth of Klebsiella pneumoniae and Aeromonas hydrophila isolated from transfusion-dependent thalassemia patients. Hemoglobin 2009, 33, 352–360. [Google Scholar] [CrossRef]
- Chong, P.M.; Lynch, T.; McCorrister, S.; Kibsey, P.; Miller, M.; Gravel, D.; Westmacott, G.R.; Mulvey, M.R.; The Canadian Nosocomial Infection Surveillance Program (CNISP). Proteomic Analysis of a NAP1 Clostridium difficile Clinical Isolate Resistant to Metronidazole. PLoS ONE 2014, 9, e82622. [Google Scholar] [CrossRef]
- Boekhoud, I.M.; Hornung, B.V.H.; Sevilla, E.; Harmanus, C.; Bos-Sanders, I.M.J.G.; Terveer, E.M.; Bolea, R.; Corver, J.; Kuijper, E.J.; Smits, W.K. Plasmid-mediated metronidazole resistance in Clostridioides difficile. Nat. Commun. 2020, 11, 598. [Google Scholar] [CrossRef]
- Gonzales-Luna, A.J.; Olaitan, A.O.; Shen, W.-J.; Deshpande, A.; Carlson, T.J.; Dotson, K.M.; Lancaster, C.; Begum, K.; Alam, M.J.; Hurdle, J.G.; et al. Reduced Susceptibility to Metronidazole Is Associated With Initial Clinical Failure in Clostridioides difficile Infection. Open Forum Infect. Dis. 2021, 8, ofab365. [Google Scholar] [CrossRef]
- Boekhoud, I.M.; Sidorov, I.; Nooij, S.; Harmanus, C.; Bos-Sanders, I.M.J.G.; Viprey, V.; Spittal, W.; Clark, E.; Davies, K.; Freeman, J.; et al. Haem is crucial for medium-dependent metronidazole resistance in clinical isolates of Clostridioides difficile. J. Antimicrob. Chemother. 2021, 76, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Darkoh, C.; DuPont, H.L.; Norris, S.J.; Kaplan, H.B. Toxin Synthesis by Clostridium difficile Is Regulated through Quorum Signaling. mBio 2015, 6, e02569-14. [Google Scholar] [CrossRef] [PubMed]
- Neemann, K.; Freifeld, A. Clostridium difficile–Associated Diarrhea in the Oncology Patient. J. Oncol. Pract. 2017, 13, 25–30. [Google Scholar] [CrossRef]
- Dubberke, E.R.; Reske, K.A.; Olsen, M.A.; McMullen, K.M.; Mayfield, J.L.; McDonald, L.C.; Fraser, V.J. Evaluation of Clostridium difficile–Associated Disease Pressure as a Risk Factor for C difficile–Associated Disease. Arch. Intern. Med. 2007, 167, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Dubberke, E.R.; Sadhu, J.; Gatti, R.; Reske, K.A.; DiPersio, J.F.; Devine, S.M.; Fraser, V. Severity of Clostridium difficile–Associated Disease (CDAD) in Allogeneic Stem Cell Transplant Recipients: Evaluation of a CDAD Severity Grading System. Infect. Control Hosp. Epidemiol. 2007, 28, 208–211. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bauer, M.; Veenendaal, D.; Verhoef, L.; Bloembergen, P.; van Dissel, J.; Kuijper, E. Clinical and microbiological characteristics of community-onset Clostridium difficile infection in The Netherlands. Clin. Microbiol. Infect. 2009, 15, 1087–1092. [Google Scholar] [CrossRef][Green Version]
- Hershko, C. Pathogenesis and management of iron toxicity in thalassemia. Ann. N. Y. Acad. Sci. 2010, 1202, 1–9. [Google Scholar] [CrossRef]
- Zhou, T.; Ma, Y.; Kong, X.; Hider, R.C. Design of iron chelators with therapeutic application. Dalton Trans. 2012, 41, 6371–6389. [Google Scholar] [CrossRef]
- Kovacevic, Z.; Kalinowski, D.S.; Lovejoy, D.B.; Quach, P.; Wong, J.; Richardson, D.R. Iron Chelators: Development of Novel Compounds with High and Selective Anti-Tumour Activity. Curr. Drug Deliv. 2010, 7, 194–207. [Google Scholar] [CrossRef]
- Liu, D.Y.; Liu, Z.D.; Hider, R.C. Oral iron chelators—Development and application. Best Pract. Res. Clin. Haematol. 2002, 15, 369–384. [Google Scholar] [CrossRef]
- Richardson, D.R.; Kalinowski, D.S.; Lau, S.; Jansson, P.J.; Lovejoy, D.B. Cancer cell iron metabolism and the development of potent iron chelators as anti-tumour agents. Biochim. Biophys. Acta Gen. Subj. 2009, 1790, 702–717. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Pattichis, K.; Neocleous, K.; Kolnagou, A. The Design and Development of Deferiprone (L1) and Other Iron Chelators for Clinical Use: Targeting Methods and Application Prospects. Curr. Med. Chem. 2004, 11, 2161–2183. [Google Scholar] [CrossRef] [PubMed]
- Nick, H.; Acklin, P.; Lattmann, R.; Buehlmayer, P.; Hauffe, S.; Schupp, J.; Alberti, D. Development of Tridentate Iron Chelators: From Desferrithiocin to ICL670. Curr. Med. Chem. 2003, 10, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Kamchonwongpaisan, S.; Paitayatat, S.; Thebtaranonth, Y.; Wilairat, P.; Yuthavong, Y. Mechanism-Based Development of New Antimalarials: Synthesis of Derivatives of Artemisinin Attached to Iron Chelators. J. Med. Chem. 1995, 38, 2311–2316. [Google Scholar] [CrossRef]
- Cappellini, M.D.; Taher, A. Deferasirox (Exjade®) for the Treatment of Iron Overload. Acta Haematol. 2009, 122, 165–173. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamaki, J.; Chawla, S.; Tong, S.; Lozada, K.A.; Yang, S. Iron Effects on Clostridioides difficile Toxin Production and Antimicrobial Susceptibilities. Antibiotics 2022, 11, 537. https://doi.org/10.3390/antibiotics11050537
Yamaki J, Chawla S, Tong S, Lozada KA, Yang S. Iron Effects on Clostridioides difficile Toxin Production and Antimicrobial Susceptibilities. Antibiotics. 2022; 11(5):537. https://doi.org/10.3390/antibiotics11050537
Chicago/Turabian StyleYamaki, Jason, Swati Chawla, Shirley Tong, Kate Alison Lozada, and Sun Yang. 2022. "Iron Effects on Clostridioides difficile Toxin Production and Antimicrobial Susceptibilities" Antibiotics 11, no. 5: 537. https://doi.org/10.3390/antibiotics11050537
APA StyleYamaki, J., Chawla, S., Tong, S., Lozada, K. A., & Yang, S. (2022). Iron Effects on Clostridioides difficile Toxin Production and Antimicrobial Susceptibilities. Antibiotics, 11(5), 537. https://doi.org/10.3390/antibiotics11050537