
Statins Induce Actin Cytoskeleton Disassembly and an Apoptosis-Like Process in Acanthamoeba spp.

 ,
, 

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. In Vitro Activity of Statin against Acanthamoeba spp.
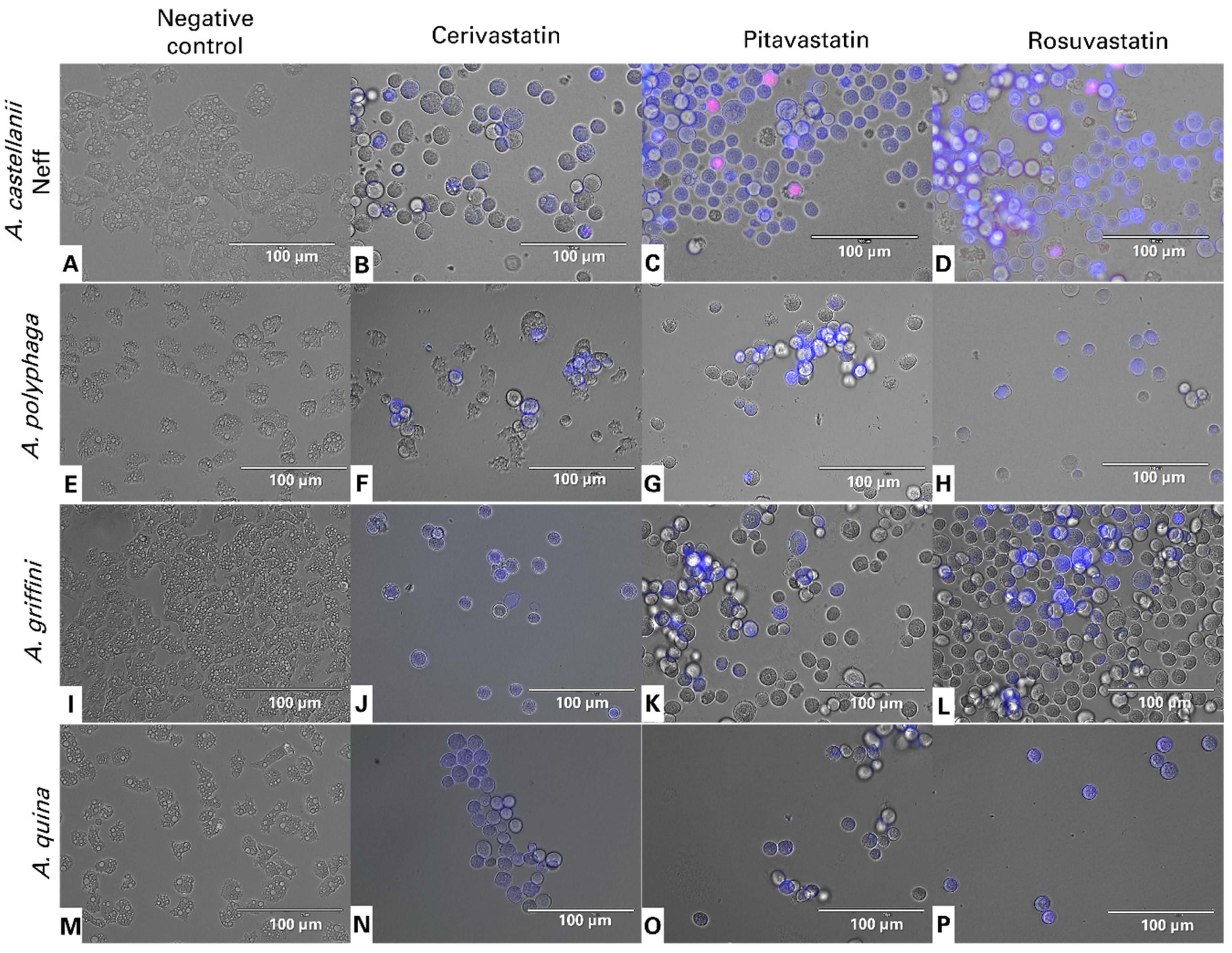
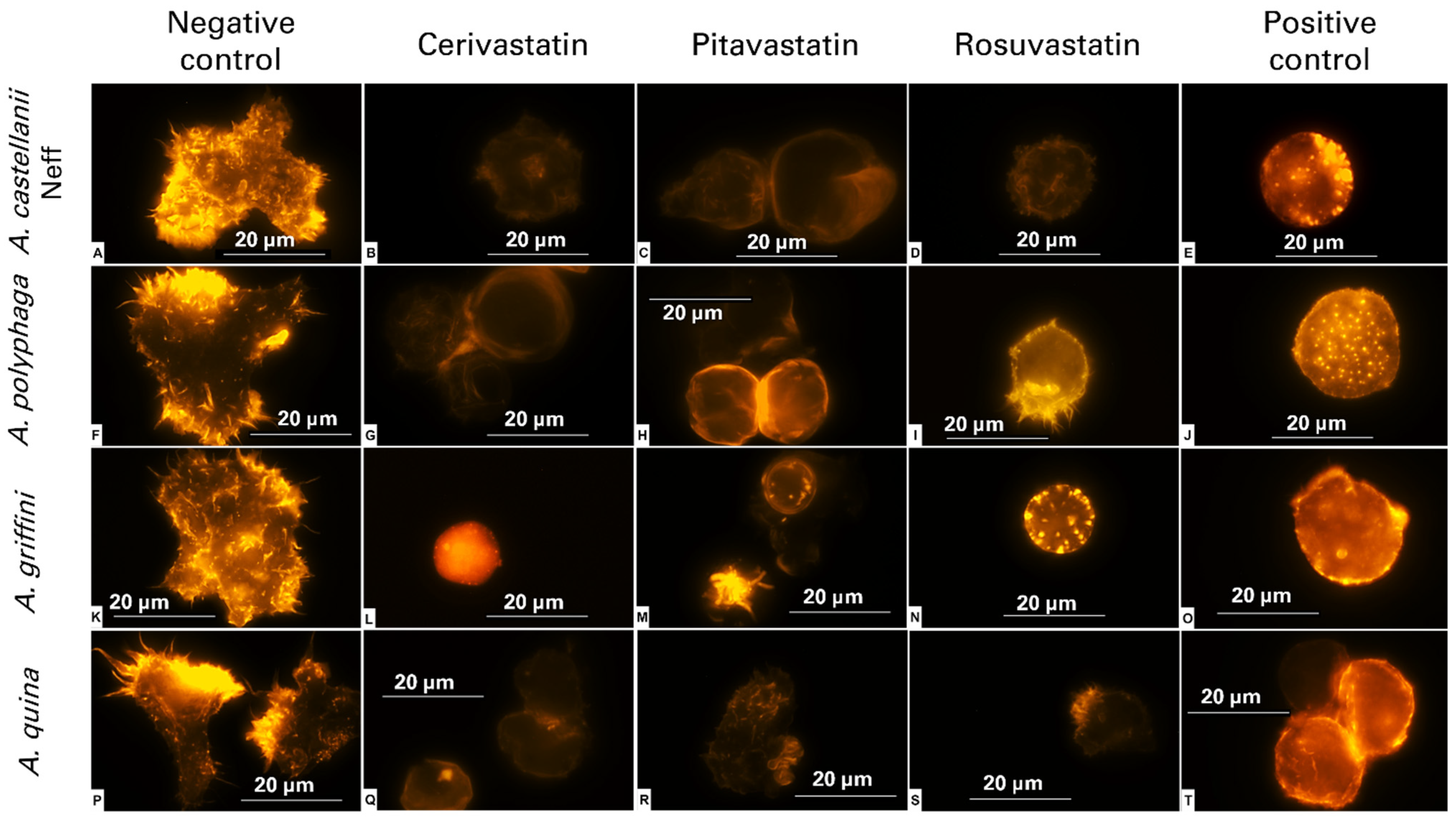
2.2. Statins Induced Actin Cytoskeleton Disassembly
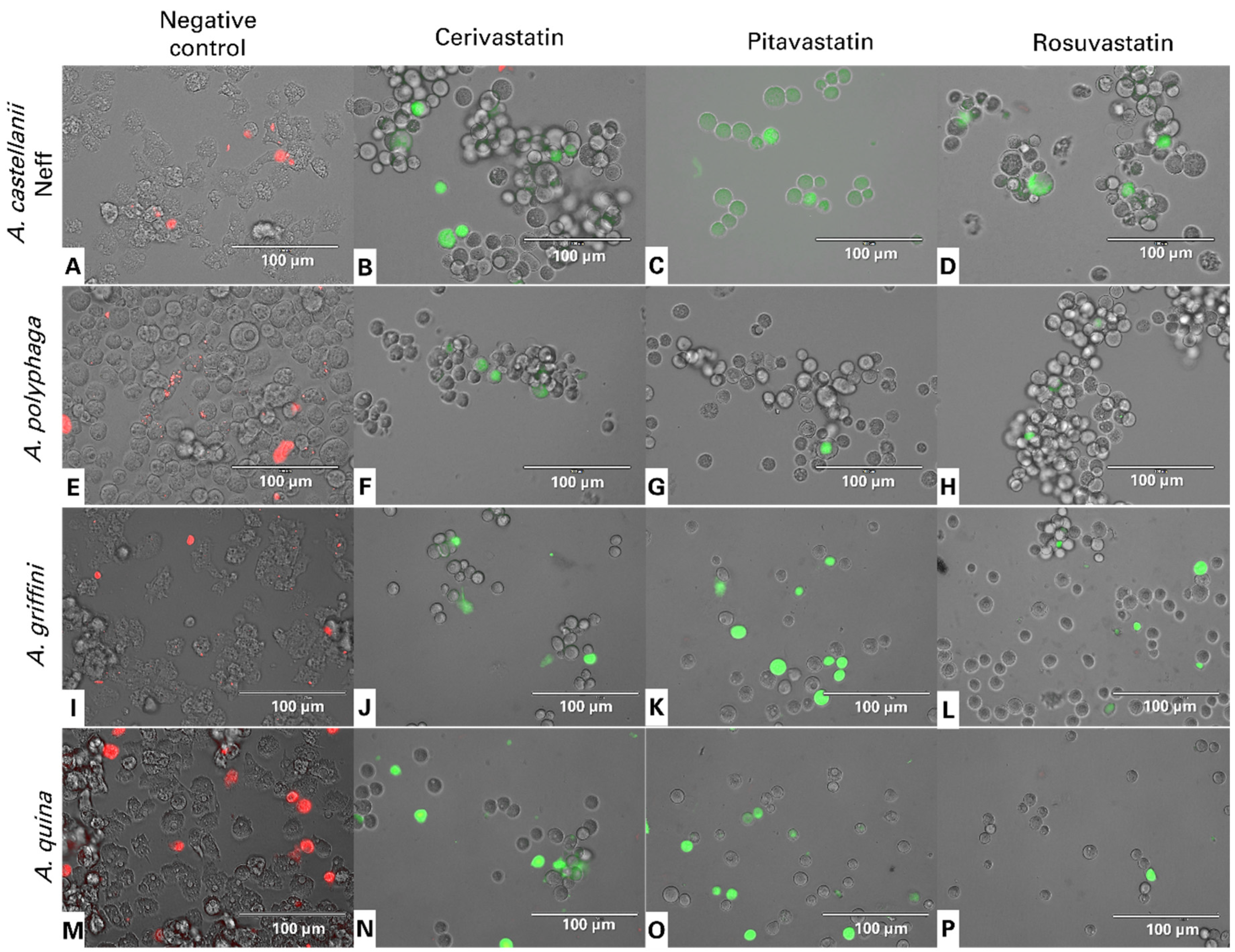
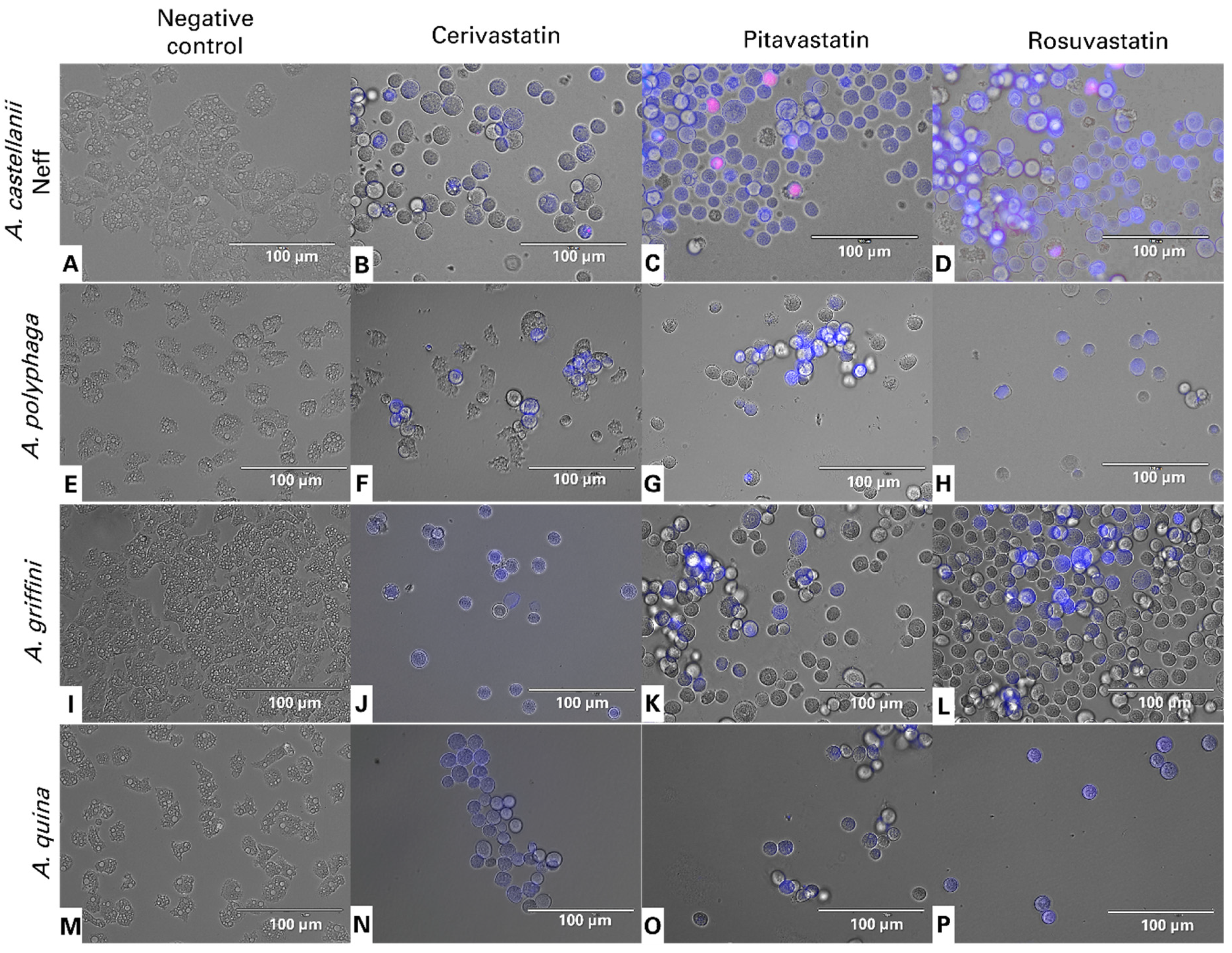
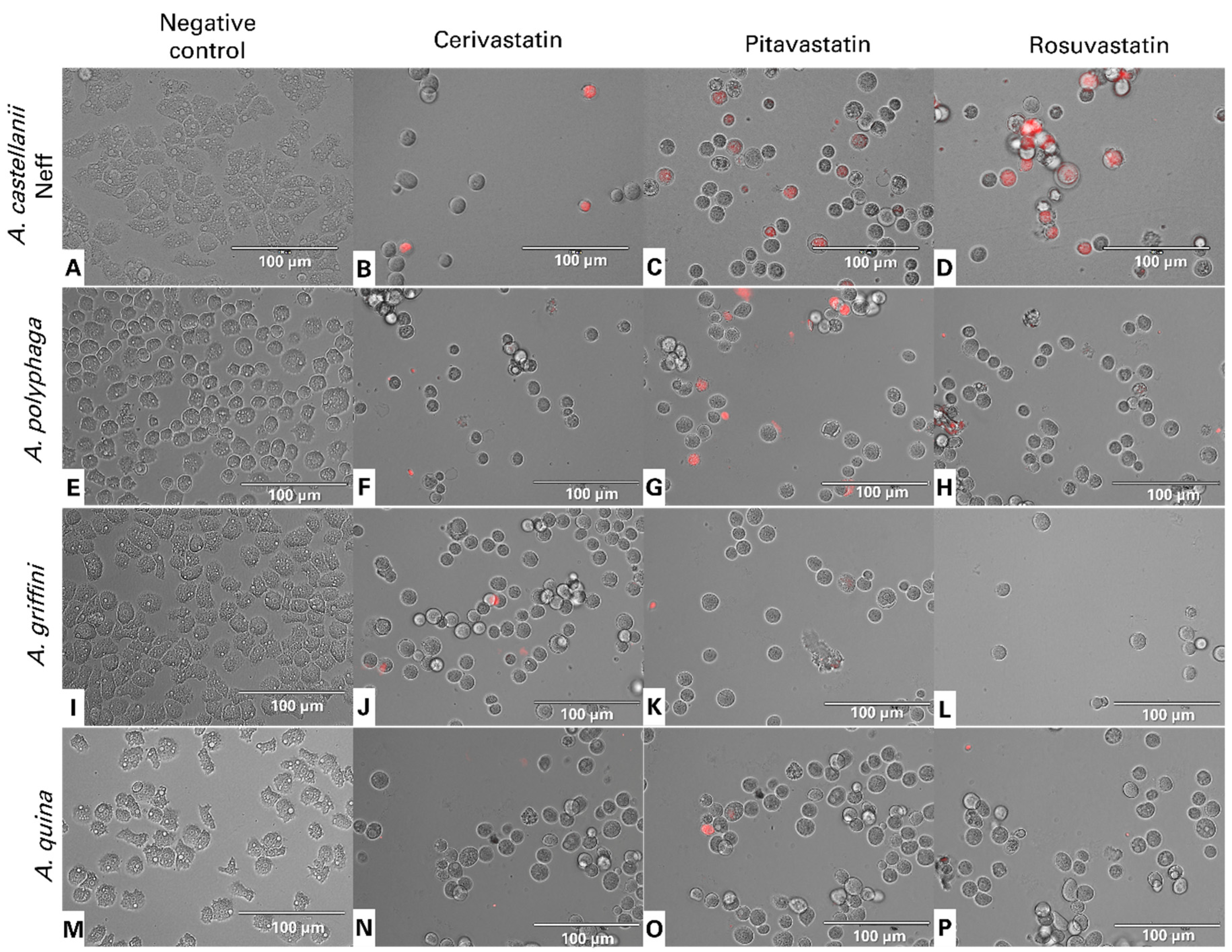
2.3. Statin Treatments Induce Compatible Events Related to Programmed Cell Death of Amoebae
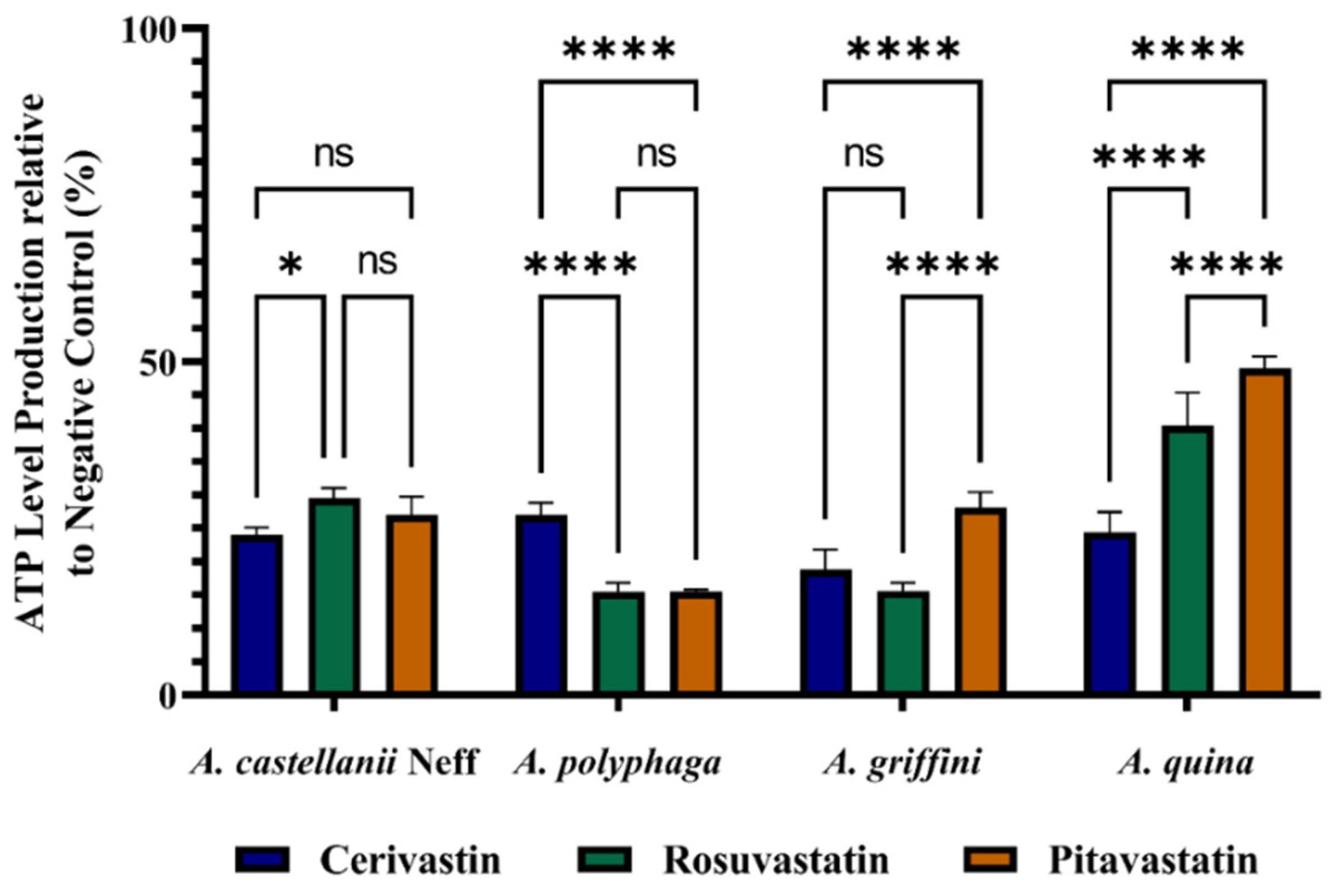
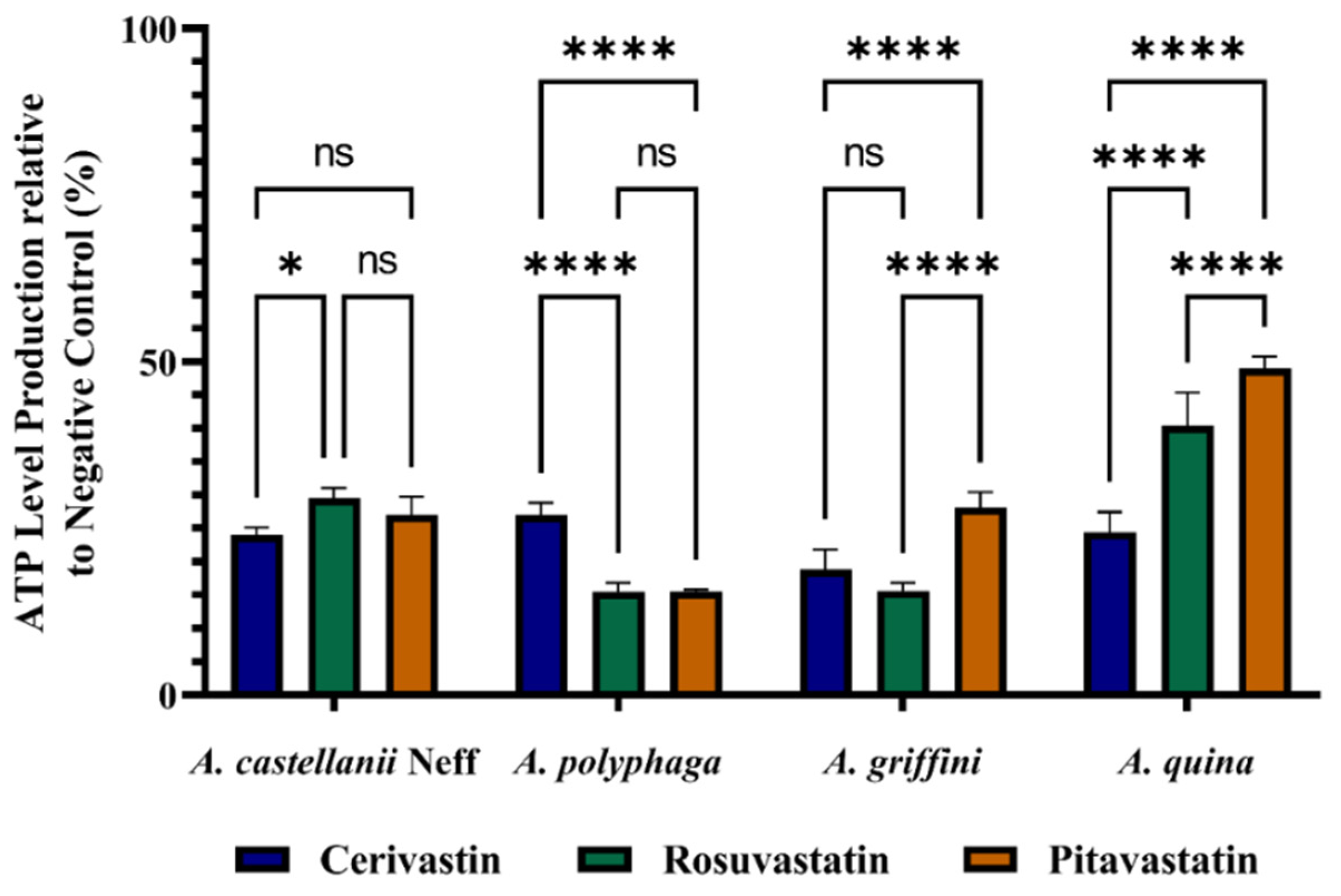
2.4. Statins Decreased ATP Levels during the Time of Incubation
2.4.1. Statins Induced the Collapse of Mitochondrial Membrane Potential (ΔΨm)
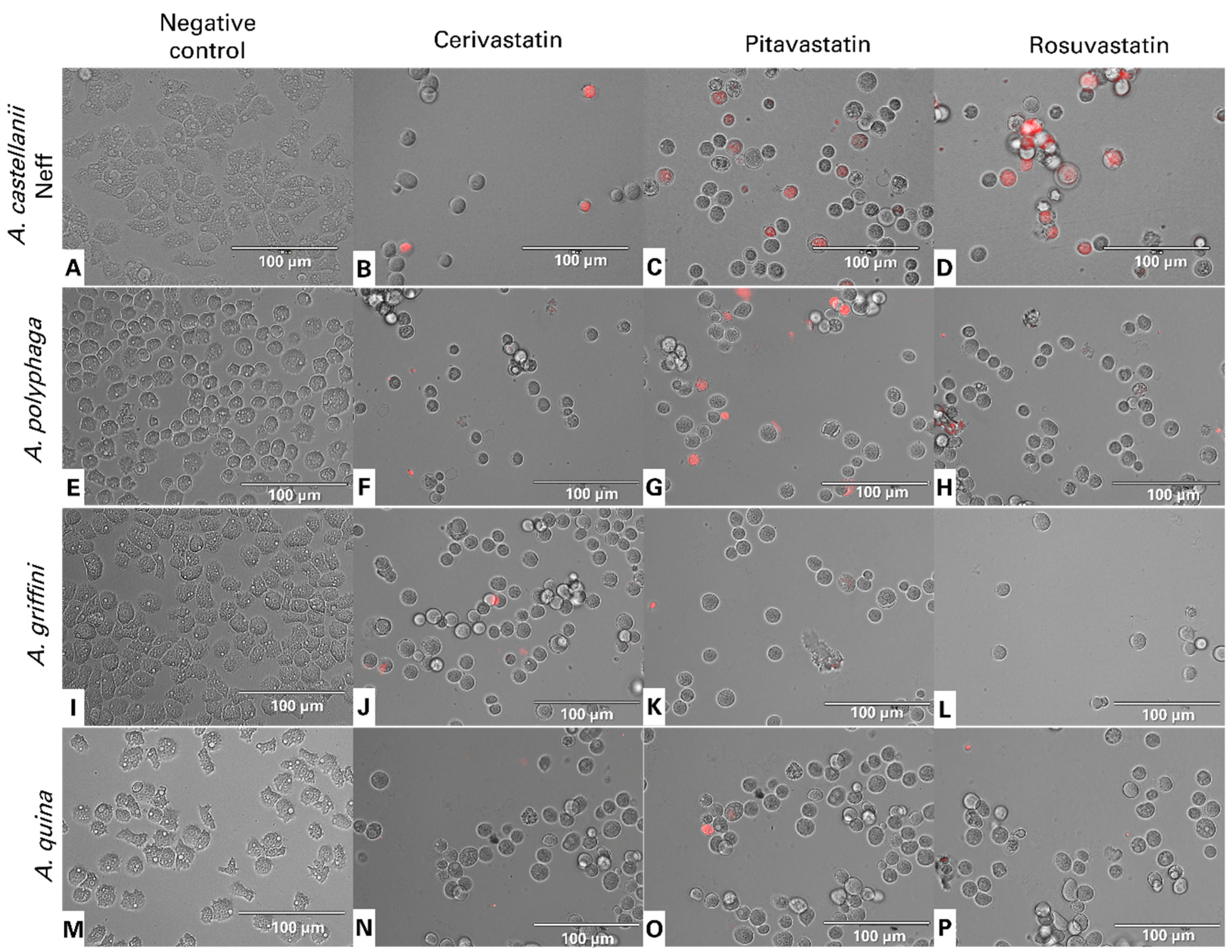
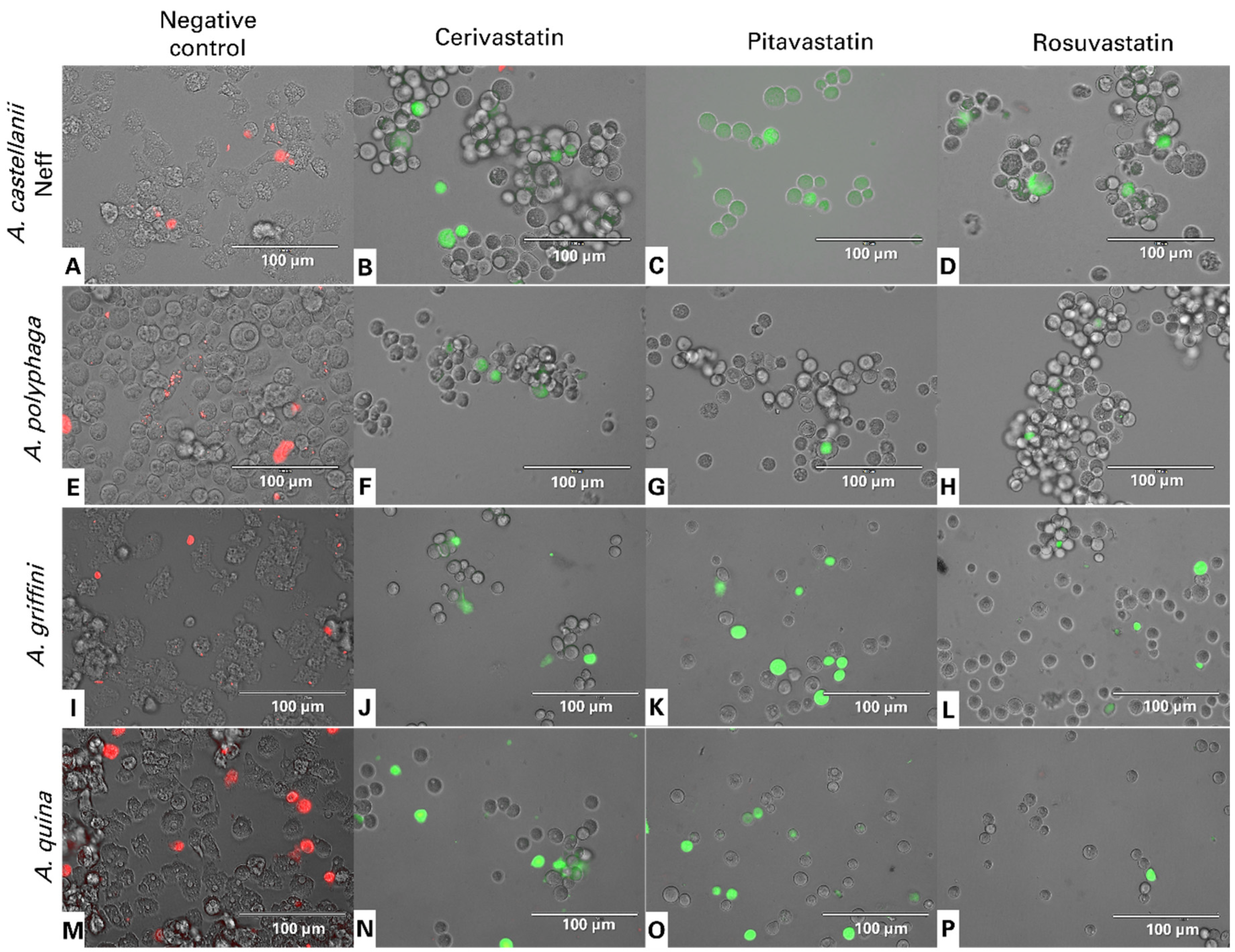
2.4.2. Statins Increase Reactive Oxygen Species (ROS) Levels in Acanthamoeba
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Acanthamoeba Strains
4.3. In Vitro Effect against the Trophozoite Stage of Acanthamoeba
4.4. In Vitro Effect against the Cyst Stage of Acanthamoeba spp.
4.5. Fluorescent Staining of Actin Distribution
4.6. Evaluation of Programmed Cell Death (PDC) Induction
4.7. Double-Stain Assay for Programmed Cell Death Determination
4.8. Measurement of ATP
4.9. Intracellular ROS Production Using CellROX® Deep Red Staining
4.10. Analysis of Mitochondrial Membrane Potential
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Martin-Navarro, C.M.; Lopez-Arencibia, A.; Sifaoui, I.; Reyes-Batlle, M.; Valladares, B.; Martinez-Carretero, E.; Pinero, J.E.; Maciver, S.K.; Lorenzo-Morales, J. Statins and voriconazole induce programmed cell death in Acanthamoeba castellanii. Antimicrob. Agents Chemother. 2015, 59, 2817–2824. [Google Scholar] [CrossRef] [Green Version]
- Thomson, S.; Rice, C.A.; Zhang, T.; Edrada-Ebel, R.; Henriquez, F.L.; Roberts, C.W. Characterisation of sterol biosynthesis and validation of 14alpha-demethylase as a drug target in Acanthamoeba. Sci. Rep. 2017, 7, 8247-z. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putaporntip, C.; Kuamsab, N.; Nuprasert, W.; Rojrung, R.; Pattanawong, U.; Tia, T.; Yanmanee, S.; Jongwutiwes, S. Analysis of Acanthamoeba genotypes from public freshwater sources in Thailand reveals a new genotype, T23 Acanthamoeba bangkokensis sp. nov. Sci. Rep. 2021, 11, 17290. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.R.; Korn, E.D. 7-Dehydrostigmasterol and ergosterol: The major sterols of an amoeba. J. Lipid Res. 1968, 9, 405–408. [Google Scholar] [CrossRef]
- Martin-Navarro, C.M.; Lorenzo-Morales, J.; Machin, R.P.; Lopez-Arencibia, A.; Garcia-Castellano, J.M.; De Fuentes, I.; Loftus, B.; Maciver, S.K.; Valladares, B.; Pinero, J.E. Inhibition of 3-Hydroxy-3-Methylglutaryl–Coenzyme A Reductase and Application of Statins as a Novel Effective Therapeutic Approach against Acanthamoeba Infections. Antimicrob. Agents Chemother. 2013, 57, 375–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudley, R.; Alsam, S.; Khan, N. Cellulose biosynthesis pathway is a potential target in the improved treatment of Acanthamoeba keratitis. Appl. Microbiol. Biotechnol. 2007, 75, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, F.; Song, Z.; Chen, P.; Liu, S.; Ouyang, L. Statin use and the risk of ovarian and endometrial cancers: A meta-analysis. BMC Cancer 2019, 19, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maji, D.; Shaikh, S.; Solanki, D.; Gaurav, K. Safety of statins. Indian J. Endocrinol. Metab. 2013, 17, 636–646. [Google Scholar] [CrossRef]
- Hahn, H.J.; Abagyan, R.; Podust, L.M.; Roy, S.; Ali, I.K.M.; Debnath, A. HMG-CoA Reductase Inhibitors as Drug Leads against Naegleria fowleri. ACS Chem. Neurosci. 2020, 11, 3089–3096. [Google Scholar] [CrossRef]
- Pozo, M.; de Nicolas, R.; Egido, J.; Gonzalez-Cabrero, J. Simvastatin inhibits the migration and adhesion of monocytic cells and disorganizes the cytoskeleton of activated endothelial cells. Eur. J. Pharmacol. 2006, 548, 53–63. [Google Scholar] [CrossRef]
- Koyuturk, M.; Ersoz, M.; Altiok, N. Simvastatin induces proliferation inhibition and apoptosis in C6 glioma cells via c-jun N-terminal kinase. Neurosci. Lett. 2004, 370, 212–217. [Google Scholar] [CrossRef]
- Rizo-Liendo, A.; Sifaoui, I.; Arberas-Jimenez, I.; Reyes-Batlle, M.; Pinero, J.E.; Lorenzo-Morales, J. Fluvastatin and atorvastatin induce programmed cell death in the brain eating amoeba Naegleria fowleri. Biomed. Pharmacother. 2020, 130, 110583. [Google Scholar] [CrossRef]
- de Souza, W.; Rodrigues, J.C. Sterol Biosynthesis Pathway as Target for Anti-trypanosomatid Drugs. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 642502. [Google Scholar] [CrossRef] [Green Version]
- Kuipers, H.F.; Rappert, A.A.; Mommaas, A.M.; van Haastert, E.S.; van der Valk, P.; Boddeke, H.W.; Biber, K.P.; van den Elsen, P.J. Simvastatin affects cell motility and actin cytoskeleton distribution of microglia. Glia 2006, 53, 115–123. [Google Scholar] [CrossRef]
- Chappell, J.; Wolf, F.; Proulx, J.; Cuellar, R.; Saunders, C. Is the Reaction Catalyzed by 3-Hydroxy-3-Methylglutaryl Coenzyme A Reductase a Rate-Limiting Step for Isoprenoid Biosynthesis in Plants? Plant Physiol. 1995, 109, 1337–1343. [Google Scholar] [CrossRef] [Green Version]
- Copaja, M.; Venegas, D.; Aranguiz, P.; Canales, J.; Vivar, R.; Avalos, Y.; Garcia, L.; Chiong, M.; Olmedo, I.; Catalan, M.; et al. Simvastatin disrupts cytoskeleton and decreases cardiac fibroblast adhesion, migration and viability. Toxicology 2012, 294, 42–49. [Google Scholar] [CrossRef]
- Sokalska, A.; Piotrowski, P.C.; Rzepczynska, I.J.; Cress, A.; Duleba, A.J. Statins inhibit growth of human theca-interstitial cells in PCOS and non-PCOS tissues independently of cholesterol availability. J. Clin. Endocrinol. Metab. 2010, 95, 5390–5394. [Google Scholar] [CrossRef] [Green Version]
- Tsubaki, M.; Takeda, T.; Kino, T.; Obata, N.; Itoh, T.; Imano, M.; Mashimo, K.; Fujiwara, D.; Sakaguchi, K.; Satou, T.; et al. Statins improve survival by inhibiting spontaneous metastasis and tumor growth in a mouse melanoma model. Am. J. Cancer Res. 2015, 5, 3186–3197. [Google Scholar]
- Lee, N.; Tilija Pun, N.; Jang, W.J.; Bae, J.W.; Jeong, C.H. Pitavastatin induces apoptosis in oral squamous cell carcinoma through activation of FOXO3a. J. Cell Mol. Med. 2020, 24, 7055–7066. [Google Scholar] [CrossRef]
- Martin-Navarro, C.M.; Lorenzo-Morales, J.; Cabrera-Serra, M.G.; Rancel, F.; Coronado-Alvarez, N.M.; Pinero, J.E.; Valladares, B. The potential pathogenicity of chlorhexidine-sensitive Acanthamoeba strains isolated from contact lens cases from asymptomatic individuals in Tenerife, Canary Islands, Spain. J. Med. Microbiol. 2008, 57, 1399–1404. [Google Scholar] [CrossRef]
- Lorenzo-Morales, J.; Kliescikova, J.; Martinez-Carretero, E.; De Pablos, L.M.; Profotova, B.; Nohynkova, E.; Osuna, A.; Valladares, B. Glycogen phosphorylase in Acanthamoeba spp.: Determining the role of the enzyme during the encystment process using RNA interference. Eukaryot. Cell 2008, 7, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Zidovetzki, R.; Levitan, I. Use of cyclodextrins to manipulate plasma membrane cholesterol content: Evidence, misconceptions and control strategies. Biochim. Biophys Acta 2007, 1768, 1311–1324. [Google Scholar] [CrossRef] [Green Version]
- Sifaoui, I.; Reyes-Batlle, M.; Lopez-Arencibia, A.; Chiboub, O.; Rodriguez-Martin, J.; Rocha-Cabrera, P.; Valladares, B.; Pinero, J.E.; Lorenzo-Morales, J. Toxic effects of selected proprietary dry eye drops on Acanthamoeba. Sci. Rep. 2018, 8, 8520. [Google Scholar] [CrossRef]
- Rodriguez-Exposito, R.L.; Nocchi, N.; Reyes-Batlle, M.; Sifaoui, I.; Suarez-Gomez, B.; Diaz-Marrero, A.R.; Souto, M.L.; Pinero, J.E.; Fernandez, J.J.; Lorenzo-Morales, J. Antiamoebic effects of sesquiterpene lactones isolated from the zoanthid Palythoa aff. clavata. Bioorg. Chem. 2021, 108, 104682. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. castellanii Neff | A. polyphaga | A. griffini | A. quina | |||||
|---|---|---|---|---|---|---|---|---|
| Trophozoite | Cyst | Trophozoite | Cyst | Trophozoite | Cyst | Trophozoite | Cyst | |
| Cerivastatin | 0.114 ± 0.050 Aa | 0.704 ± 0.129 Aa | 0.939 ± 0.102 Ca | 20.037 ± 0.042 Da | 0.384 ± 0.064 Ba | 2.108 ± 0.046 Ba | 0.274 ± 0.046 ABa | 5.531 ± 0.941 Ca |
| Rosuvastatin | 5.583 ± 0.750 Ab | 18.168 ± 2.064 Ab | 16.463 ± 0.939 Cb | >100 Cc | 8.582 ± 1.200 Bb | 23.738 ± 3.853 Ab | 5.659 ± 1.117 Ab | 81.620 ± 4.964 Bb |
| Pitavastatin | 0.389 ± 0.047 ABa | 3.040 ± 0.249 Aa | 1.907 ± 0.364 Ca | 48.774 ± 3.031 Bb | 0.859 ± 0.093 Ba | 5.306 ± 0.068 Aa | 0.229 ± 0.035 Aa | 5.921 ± 0.159 Aa |
| A. castellanii Neff | A. polyphaga | A. griffini | A. quina | |
|---|---|---|---|---|
| Cerivastatin | 0.305 ± 0.166 | 2.010 ± 0.010 | 1.850 ± 0.280 | 0.901 ± 0.143 |
| Rosuvastatin | 12.806 ± 2.081 | 47.728 ± 5.362 | 26.552 ± 5.583 | 14.010 ± 2.025 |
| Pitavastatin | 0.859 ± 0.163 | 5.208 ± 1.106 | 6.817 ± 4.728 | 0.695 ± 0.075 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Expósito, R.L.; Sifaoui, I.; Reyes-Batlle, M.; Maciver, S.K.; Piñero, J.E.; Lorenzo-Morales, J. Statins Induce Actin Cytoskeleton Disassembly and an Apoptosis-Like Process in Acanthamoeba spp. Antibiotics 2022, 11, 280. https://doi.org/10.3390/antibiotics11020280
Rodríguez-Expósito RL, Sifaoui I, Reyes-Batlle M, Maciver SK, Piñero JE, Lorenzo-Morales J. Statins Induce Actin Cytoskeleton Disassembly and an Apoptosis-Like Process in Acanthamoeba spp. Antibiotics. 2022; 11(2):280. https://doi.org/10.3390/antibiotics11020280
Chicago/Turabian StyleRodríguez-Expósito, Rubén L., Ines Sifaoui, María Reyes-Batlle, Sutherland K. Maciver, José E. Piñero, and Jacob Lorenzo-Morales. 2022. "Statins Induce Actin Cytoskeleton Disassembly and an Apoptosis-Like Process in Acanthamoeba spp." Antibiotics 11, no. 2: 280. https://doi.org/10.3390/antibiotics11020280
APA StyleRodríguez-Expósito, R. L., Sifaoui, I., Reyes-Batlle, M., Maciver, S. K., Piñero, J. E., & Lorenzo-Morales, J. (2022). Statins Induce Actin Cytoskeleton Disassembly and an Apoptosis-Like Process in Acanthamoeba spp. Antibiotics, 11(2), 280. https://doi.org/10.3390/antibiotics11020280