

Evaluation of a Sequential Antibiotic Treatment Regimen of Ampicillin, Ciprofloxacin and Fosfomycin against Escherichia coli CFT073 in the Hollow Fiber Infection Model Compared with Simultaneous Combination Treatment

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
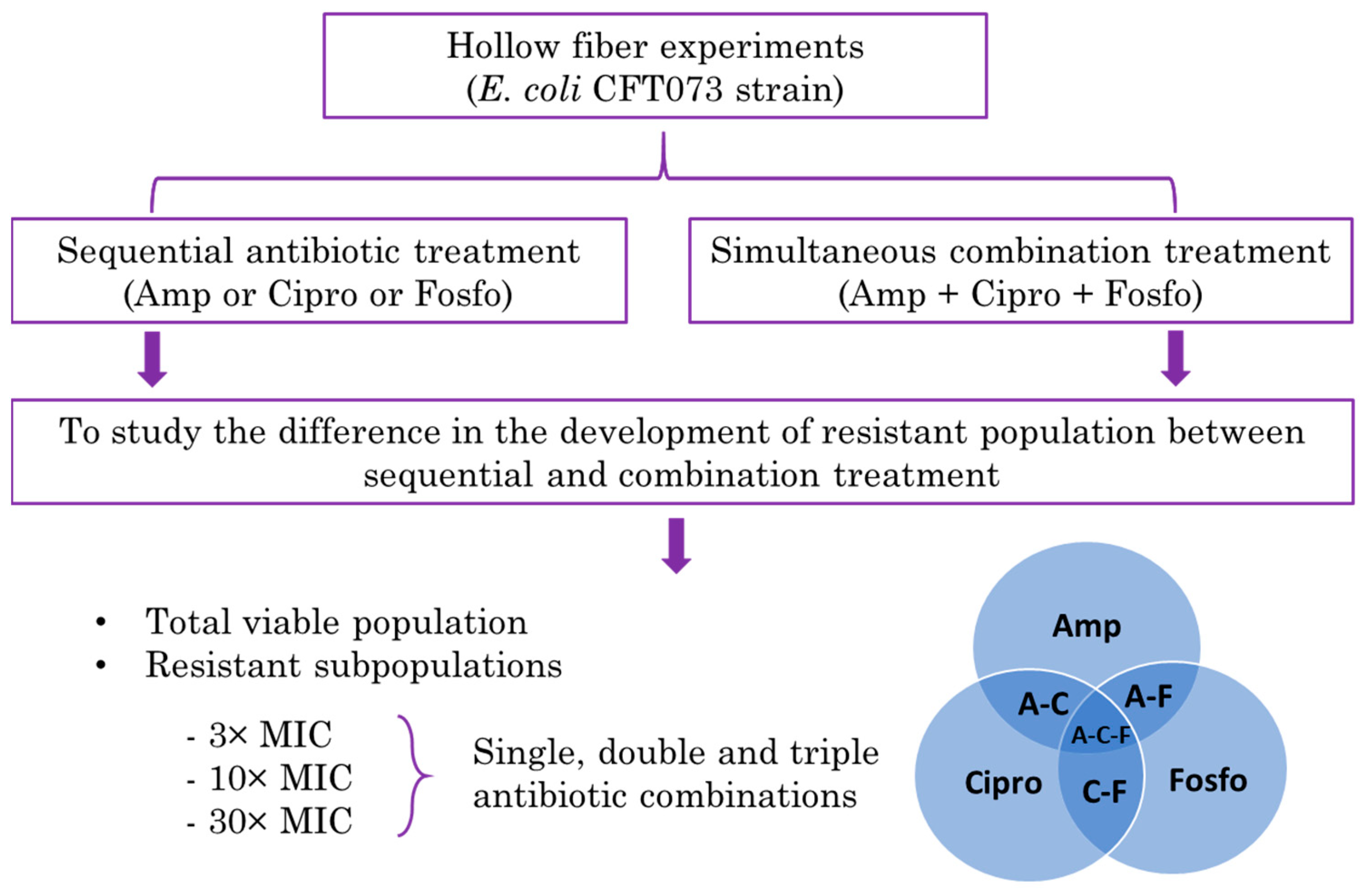
Abstract
1. Introduction
2. Results
2.1. Antibiotic Treatment Effect on Total E. coli Population
2.2. Single Antibiotic-resistant E. coli Subpopulations
2.3. Double Antibiotic-Resistant E. coli Subpopulations
2.4. Triple Antibiotic-resistant E. coli Subpopulations
2.5. Effect of Single Antibiotics on Total and 3× MIC Resistant E. coli Subpopulations
3. Discussion
4. Materials and Methods
4.1. Microorganisms, Antibiotics, Media, and Agar
4.2. In Vitro Hollow Fiber Infection Model
4.3. Assessment of Microbiological Response
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Disclaimer
Abbreviations
References
- Sengupta, S.; Chattopadhyay, M.K.; Grossart, H.P. The multifaceted roles of antibiotics and antibiotic resistance in nature. Front. Microbiol. 2013, 4, 47. [Google Scholar] [CrossRef] [PubMed]
- Exner, M. Antibiotic resistance: What is so special about multidrug-resistant Gram-negative bacteria? GMS Hyg. Infect. Control. 2017, 12, Doc05. [Google Scholar] [PubMed]
- Theuretzbacher, U. Global antimicrobial resistance in Gram-negative pathogens and clinical need. Curr. Opin. Microbiol. 2017, 39, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Karaiskos, I.; Antoniadou, A.; Giamarellou, H. Combination therapy for extensively-drug resistant gram-negative bacteria. Expert Rev. Anti. Infect. Ther. 2017, 15, 1123–1140. [Google Scholar] [CrossRef]
- Ambrose, P.G.; Bhavnani, S.M.; Rubino, C.M.; Louie, A.; Gumbo, T.; Forrest, A.; Drusano, G.L. Pharmacokinetics-pharmacodynamics of antimicrobial therapy: It’s not just for mice anymore. Clin. Infect. Dis. 2007, 44, 79–86. [Google Scholar] [CrossRef]
- Drusano, G.L. Antimicrobial pharmacodynamics: Critical interactions of ‘bug and drug’. Nat. Rev. Microbiol. 2004, 2, 289–300. [Google Scholar] [CrossRef]
- Bulitta, J.B.; Hope, W.W.; Eakin, A.E.; Guina, T.; Tam, V.H.; Louie, A.; Drusano, G.L.; Hoover, J.L. Generating robust and informative nonclinical in vitro and in vivo bacterial infection model efficacy data to support translation to humans. Antimicrob. Agents Chemother. 2019, 63, 1–24. [Google Scholar] [CrossRef]
- VanScoy, B.; McCauley, J.; Bhavnani, S.M.; Ellis-Grosse, E.J.; Ambrose, P.G. Relationship between Fosfomycin Exposure and Amplification of Escherichia coli Subpopulations with Reduced Susceptibility in a Hollow-Fiber Infection Model. Antimicrob. Agents Chemother. 2016, 60, 5141–5145. [Google Scholar] [CrossRef]
- McEntee, L.; Johnson, A.; Farrington, N.; Unsworth, J.; Dane, A.; Jain, A.; Cotroneo, N.; Critchley, I.; Melnick, D.; Parr, T.; et al. Pharmacodynamics of Tebipenem: New Options for Oral Treatment of Multidrug-Resistant Gram-Negative Infections. Antimicrob. Agents Chemother. 2019, 63, e00603-19. [Google Scholar] [CrossRef]
- Cai, Y.; Lim, T.P.; Teo, J.Q.; Sasikala, S.; Chan, E.C.; Hong, Y.J.; Lee, W.; Tan, T.Y.; Tan, T.T.; Koh, T.H.; et al. Evaluating Polymyxin B-Based Combinations against Carbapenem-Resistant Escherichia coli in Time-Kill Studies and in a Hollow-Fiber Infection Model. Antimicrob. Agents Chemother. 2017, 61, e01509-16. [Google Scholar] [CrossRef]
- Chait, R.; Craney, A.; Kishony, R. Antibiotic interactions that select against resistance. Nature 2007, 446, 668–671. [Google Scholar] [CrossRef] [PubMed]
- Levin-Reisman, I.; Ronin, I.; Gefen, O.; Braniss, I.; Shoresh, N.; Balaban, N.Q. Antibiotic tolerance facilitates the evolution of resistance. Science 2017, 355, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Band, V.I.; Hufnagel, D.A.; Jaggavarapu, S.; Sherman, E.X.; Wozniak, J.E.; Satola, S.W.; Farley, M.M.; Jacob, J.T.; Burd, E.M.; Weiss, D.S. Antibiotic combinations that exploit heteroresistance to multiple drugs effectively control infection. Nat. Microbiol. 2019, 4, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Principe, L.; D’Arezzo, S.; Capone, A.; Petrosillo, N.; Visca, P. In vitro activity of tigecycline in combination with various antimicrobials against multidrug resistant Acinetobacter baumannii. Ann. Clin. Microbiol. Antimicrob. 2009, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Urban, C.; Mariano, N.; Rahal, J.J. In vitro double and triple bactericidal activities of doripenem, polymyxin B, and rifampin against multidrug-resistant Acinetobacter baumannii, Pseudomonas aeruginosa, Klebsiella pneumoniae, and Escherichia coli. Antimicrob. Agents Chemother. 2010, 54, 2732–2734. [Google Scholar] [CrossRef]
- Bergen, P.J.; Tsuji, B.T.; Bulitta, J.B.; Forrest, A.; Jacob, J.; Sidjabat, H.E.; Paterson, D.L.; Nation, R.L.; Li, J. Synergistic killing of multidrug-resistant Pseudomonas aeruginosa at multiple inocula by colistin combined with doripenem in an in vitro pharmacokinetic/pharmacodynamic model. Antimicrob. Agents Chemother. 2011, 55, 5685–5695. [Google Scholar] [CrossRef]
- Bercot, B.; Poirel, L.; Dortet, L.; Nordmann, P. In vitro evaluation of antibiotic synergy for NDM-1-producing Enterobacteriaceae. J. Antimicrob. Chemother. 2011, 66, 2295–2297. [Google Scholar] [CrossRef]
- Tangden, T.; Hickman, R.A.; Forsberg, P.; Lagerback, P.; Giske, C.G.; Cars, O. Evaluation of double- and triple-antibiotic combinations for VIM- and NDM-producing Klebsiella pneumoniae by in vitro time-kill experiments. Antimicrob. Agents Chemother. 2014, 58, 1757–1762. [Google Scholar] [CrossRef]
- Pena-Miller, R.; Laehnemann, D.; Jansen, G.; Fuentes-Hernandez, A.; Rosenstiel, P.; Schulenburg, H.; Beardmore, R. When the most potent combination of antibiotics selects for the greatest bacterial load: The Smile-Frown transition. PLOS Biol. 2013, 11, e1001540. [Google Scholar] [CrossRef]
- Batra, A.; Roemhild, R.; Rousseau, E.; Franzenburg, S.; Niemann, S.; Schulenburg, H. High potency of sequential therapy with only β-lactam antibiotics. eLife 2021, 10, e68876. [Google Scholar] [CrossRef]
- Roemhild, R.; Gokhale, C.S.; Dirksen, P.; Blake, C.; Rosenstiel, P.; Traulsen, A.; Andersson, D.I.; Schulenburg, H. Cellular hysteresis as a principle to maximize the efficacy of antibiotic therapy. Proc. Natl. Acad. Sci. USA 2018, 115, 9767–9772. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, C.; Römhild, R.; Rosenstiel, P.; Schulenburg, H. Evolutionary stability of collateral sensitivity to antibiotics in the model pathogen Pseudomonas aeruginosa. eLife 2019, 8, e51481. [Google Scholar] [CrossRef] [PubMed]
- Garimella, N.; Zere, T.; Hartman, N.; Gandhi, A.; Bekele, A.; Li, X.; Stone, H.; Sacks, L.; Weaver, J.L. Effect of drug combinations on the kinetics of antibiotic resistance emergence in Escherichia coli CFT073 using an in vitro hollow-fibre infection model. Int. J. Antimicrob. Agents 2020, 55, 105861. [Google Scholar] [CrossRef] [PubMed]
- Breijyeh, Z.; Jubeh, B.; Karaman, R. Resistance of Gram-Negative Bacteria to Current Antibacterial Agents and Approaches to Resolve It. Molecules 2020, 25, 1340. [Google Scholar] [CrossRef] [PubMed]
- Tyers, M.; Wright, G.D. Drug combinations: A strategy to extend the life of antibiotics in the 21st century. Nat. Rev. Microbiol. 2019, 17, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Sanders, C.C.; Sanders, W.E.J.; Goering, R.V.; Werner, V. Selection of multiple antibiotic resistance by quinolones, beta-lactams, and aminoglycosides with special reference to cross-resistance between unrelated drug classes. Antimicrob. Agents Chemother. 1984, 26, 797–801. [Google Scholar] [CrossRef] [PubMed]
- van Loon, H.J.; Vriens, M.R.; Fluit, A.C.; Troelstra, A.; van der Werken, C.; Verhoef, J.; Bonten, M.J. Antibiotic rotation and development of gram-negative antibiotic resistance. Am. J. Respir. Crit. Care Med. 2005, 171, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Beardmore, R.E.; Pena-Miller, R.; Gori, F.; Iredell, J. Antibiotic Cycling and Antibiotic Mixing: Which One Best Mitigates Antibiotic Resistance? Mol. Biol. Evol. 2017, 34, 802–817. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Hernandez, A.; Plucain, J.; Gori, F.; Pena-Miller, R.; Reding, C.; Jansen, G.; Schulenburg, H.; Gudelj, I.; Beardmore, R. Using a sequential regimen to eliminate bacteria at sublethal antibiotic dosages. PLoS Biol. 2015, 13, e1002104. [Google Scholar] [CrossRef] [PubMed]
- Roemhild, R.; Barbosa, C.; Beardmore, R.E.; Jansen, G.; Schulenburg, H. Temporal variation in antibiotic environments slows down resistance evolution in pathogenic Pseudomonas aeruginosa. Evol. Appl. 2015, 8, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Maltas, J.; Wood, K.B. Pervasive and diverse collateral sensitivity profiles inform optimal strategies to limit antibiotic resistance. PLOS Biol. 2019, 17, e3000515. [Google Scholar] [CrossRef]
- Falagas, M.E.; Tansarli, G.S.; Karageorgopoulos, D.E.; Vardakas, K.Z. Deaths attributable to carbapenem-resistant Enterobacteriaceae infections. Emerg. Infect. Dis. 2014, 20, 1170–1175. [Google Scholar] [CrossRef]
- Taggar, G.; Attiq Rheman, M.; Boerlin, P.; Diarra, M.S. Molecular Epidemiology of Carbapenemases in Enterobacteriales from Humans, Animals, Food and the Environment. Antibiotics 2020, 9, 693. [Google Scholar] [CrossRef] [PubMed]
- Salabi, A.E.; Walsh, T.R.; Chouchani, C. Extended spectrum β-lactamases, carbapenemases and mobile genetic elements responsible for antibiotics resistance in Gram-negative bacteria. Crit. Rev. Microbiol. 2013, 39, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P., III; Clark, N.M.; Zhanel, G.G. Escalating antimicrobial resistance among Enterobacteriaceae: Focus on carbapenemases. Expert Opin. Pharmacother. 2021, 22, 1455–1474. [Google Scholar] [CrossRef]
- Li, X.-Z.; Plesiat, P.; Nikaido, H. The Challenge of Efflux-Mediated Antibiotic Resistance in GramNegative Bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed]
- Brochado, A.R.; Telzerow, A.; Bobonis, J.; Banzhaf, M.; Mateus, A.; Selkrig, J.; Huth, E.; Bassler, S.; Beas, J.Z.; Zietek, M.; et al. Species-specific activity of antibacterial drug combinations. Nature 2018, 559, 259–263. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krishna, A.; Zere, T.; Mistry, S.; Ismaiel, O.; Stone, H.; Sacks, L.V.; Weaver, J.L. Evaluation of a Sequential Antibiotic Treatment Regimen of Ampicillin, Ciprofloxacin and Fosfomycin against Escherichia coli CFT073 in the Hollow Fiber Infection Model Compared with Simultaneous Combination Treatment. Antibiotics 2022, 11, 1705. https://doi.org/10.3390/antibiotics11121705
Krishna A, Zere T, Mistry S, Ismaiel O, Stone H, Sacks LV, Weaver JL. Evaluation of a Sequential Antibiotic Treatment Regimen of Ampicillin, Ciprofloxacin and Fosfomycin against Escherichia coli CFT073 in the Hollow Fiber Infection Model Compared with Simultaneous Combination Treatment. Antibiotics. 2022; 11(12):1705. https://doi.org/10.3390/antibiotics11121705
Chicago/Turabian StyleKrishna, Ashok, Tesfalem Zere, Sabyasachy Mistry, Omnia Ismaiel, Heather Stone, Leonard V. Sacks, and James L. Weaver. 2022. "Evaluation of a Sequential Antibiotic Treatment Regimen of Ampicillin, Ciprofloxacin and Fosfomycin against Escherichia coli CFT073 in the Hollow Fiber Infection Model Compared with Simultaneous Combination Treatment" Antibiotics 11, no. 12: 1705. https://doi.org/10.3390/antibiotics11121705
APA StyleKrishna, A., Zere, T., Mistry, S., Ismaiel, O., Stone, H., Sacks, L. V., & Weaver, J. L. (2022). Evaluation of a Sequential Antibiotic Treatment Regimen of Ampicillin, Ciprofloxacin and Fosfomycin against Escherichia coli CFT073 in the Hollow Fiber Infection Model Compared with Simultaneous Combination Treatment. Antibiotics, 11(12), 1705. https://doi.org/10.3390/antibiotics11121705