

Comparison of Antimicrobial Susceptibility Profiles of Thermotolerant Campylobacter spp. Isolated from Human and Poultry Samples in Georgia (Caucasus)

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Transport
2.2. Detection and Phenotypic Identification of Campylobacter spp.
2.3. Confirmation of Campylobacter Species and Differentiation by Real-Time PCR Analysis
2.4. Antimicrobial Susceptibility Testing
2.5. NGS Methodology
2.6. Statistical Analyses
3. Results
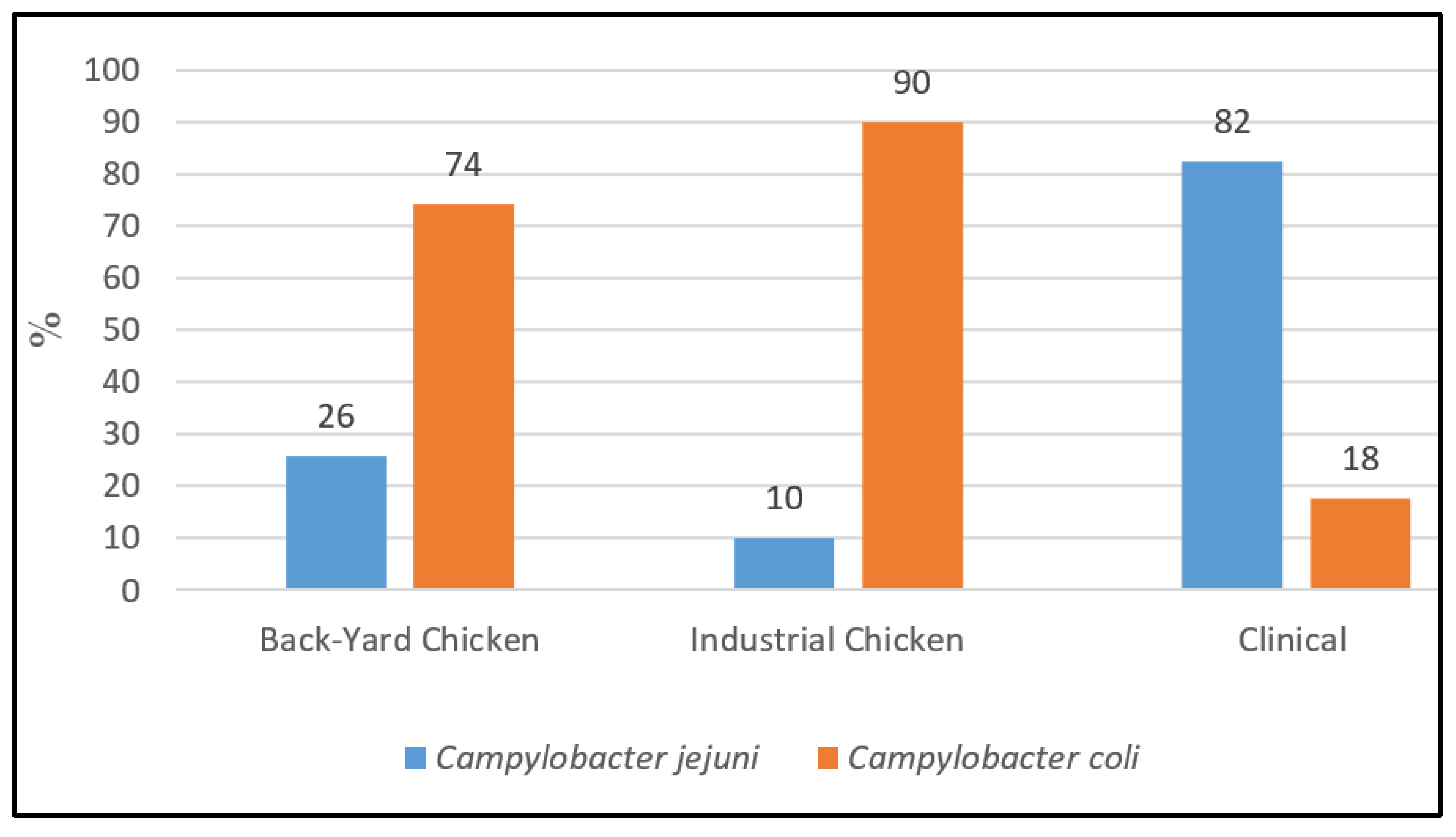
3.1. Collection of Campylobacter spp. Strains and Identification of Species
3.2. Prevalence of Antimicrobial Resistance in Campylobacter Isolates
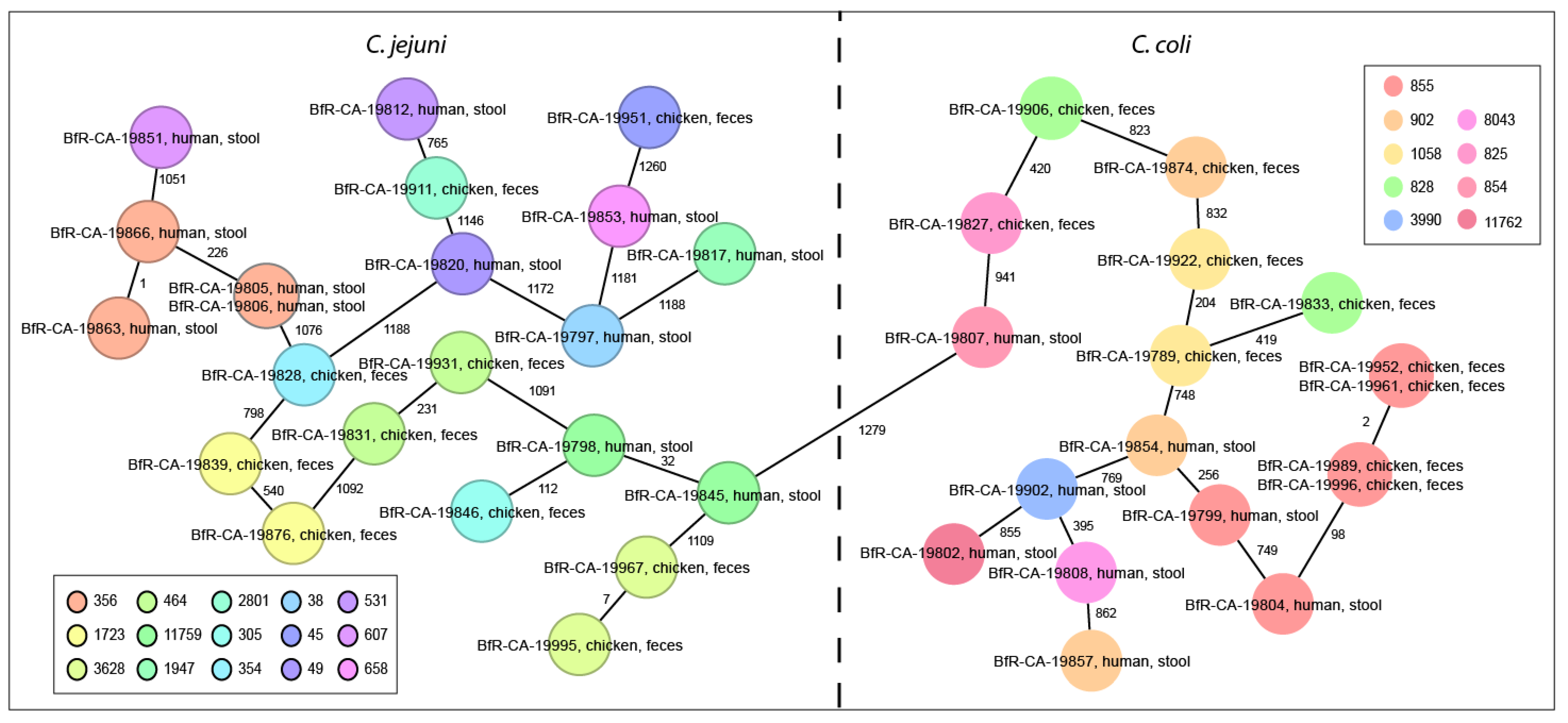
3.3. Campylobacter spp. Isolates Are Phylogenetically Diverse
3.4. Detection of Antimicrobial Resistance Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Competing Interests
References
- WHO Regional Office for Europe/European Centre for Disease Prevention and Control. Antimicrobial Resistance Surveillance in Europe 2022–2020 Data; WHO Regional Office for Europe: Copenhagen, Denmark, 2022. [CrossRef]
- World Health Organization (WHO). Available online: https://www.who.int/news-room/fact-sheets/detail/campylobacter (accessed on 23 July 2022).
- Rosner, B.M.; Schielke, A.; Didelot, X.; Kops, F.; Breidenbach, J.; Willrich, N.; Gölz, G.; Alter, T.; Stingl, K.; Josenhans, C.; et al. A combined case-control and molecular source attribution study of human Campylobacter infections in Germany, 2011–2014. Sci. Rep. 2017, 7, 5139. [Google Scholar] [CrossRef] [PubMed]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Joint Report by ECDC, EFSA, EMA, and OECD on Antimicrobial Resistance in the EU/EEA and a One Health Response. 2022. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/antimicrobial-resistance-policy-brief-2022.pdf (accessed on 7 September 2022).
- EFSA (European Food Safety Authority) and ECDC (European Centre for Disease Prevention and Control). The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J. 2022, 20, e07209. [Google Scholar] [CrossRef]
- European Medicines Agency (EMA). Categorization of Antibiotics in the European Union. EMA/CVMP/CHMP/682198/2017. 2019. Available online: https://bit.ly/30ZEuRi (accessed on 7 September 2022).
- National Food Safety Agency (NFA) of Georgia. Register of Authorized Medications for Veterinary Use. Available online: https://nfa.gov.ge/Ge/Page/InformationAboutVeterinaryDrugs (accessed on 7 September 2022). (In Georgian)
- ECDC (European Centre for Disease Prevention and Control); EFSA (European Food Safety Authority); EMA (European Medicines Agency). Third Joint INTER-Agency Report on the Integrated Analysis of Consumption of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Humans and Food-Producing Animals in the EU/EEA.; Joint Interagency Antimicrobial Consumption and Resistance Analysis (JIACRA) III Report 2016–2018; EDCD: Stockholm, Sweden; EFSA: Parma, Italy; EMA: Amsterdam, The Netherlands, 2021. [CrossRef]
- World Health Organization (WHO). Central Asian and European Surveillance of Antimicrobial Resistance (CAESAR), Annual Report 2020. Available online: https://www.euro.who.int/__data/assets/pdf_file/0003/469200/Central-Asian-and-European-Surveillance-of-Antimicrobial-Resistance.-Annual-report-2020-eng.pdf (accessed on 7 September 2022).
- European Parliament and Council. Directive 2003/99/EC of the European Parliament and of the Council of 17 November 2003 on the Monitoring of Zoonoses and Zoonotic Agents. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2003:325:0031:0040:EN:PDF (accessed on 23 July 2022).
- Metreveli, M.; Bulia, S.; Shalamberidze, I.; Tevzadze, L.; Tsanava, S.; Goenaga, J.C.; Stingl, K.; Imnadze, P. Campylobacteriosis, Shigellosis and Salmonellosis in Hospitalized Children with Acute Inflammatory Diarrhea in Georgia. Pathogens 2022, 11, 232. [Google Scholar] [CrossRef]
- Bolton, F.J.; Robertson, L. A selective medium for isolating Campylobacter jejuni/coli. J. Clin. Pathol. 1982, 35, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Mayr, A.M.; Lick, S.; Bauer, J.; Thärigen, D.; Busch, U.; Huber, I. Rapid detection and differentiation of Campylobacter jejuni, Campylobacter coli, and Campylobacter lari in food, using multiplex real-time PCR. J. Food Prot. 2010, 73, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Detection of Campylobacter spp. In Minced Meat-Real-Time PCR Methods; German Food and Feed Code (LFGB) L 06.32-1 (2013-08); Federal Office of Consumer Protection and Food Safety (BVL): Beuth GmbH, Germany, 2013. (In German) [Google Scholar]
- Anderson, A.; Pietsch, K.; Zucker, R.; Mayr, A.; Müller-Hohe, E.; Messelhäusser, U.; Sing, A.; Busch, U.; Huber, I. Validation of a Duplex Real-Time PCR for the Detection of Salmonella spp. in Different Food Products. Food Anal. Methods 2010, 4, 259–267. [Google Scholar] [CrossRef]
- Commission Implementing Decision (EU) 2020/1729 of 17 November 2020 on the Monitoring and Reporting of Antimicrobial Resistance in Zoonotic and Commensal Bacteria and Repealing Implementing Decision 2013/652/EU. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32020D1729&rid=4 (accessed on 7 September 2022).
- CLSI. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015; CLSI guideline M45. [Google Scholar]
- CLSI. Methods for Antimicrobial Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria Isolated from Animals, 1st ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017; CLSI supplement VET06. [Google Scholar]
- Cody, A.J.; Bray, J.E.; Jolley, K.A.; McCarthy, N.D.; Maiden, M. Core Genome Multilocus Sequence Typing Scheme for Stable, Comparative Analyses of Campylobacter jejuni and C. coli Human Disease Isolates. J. Clin. Microbiol. 2017, 55, 2086–2097. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef]
- Schwengers, O.; Barth, P.; Falgenhauer, L.; Hain, T.; Chakraborty, T.; Goesmann, A. Platon: Identification and characterization of bacterial plasmid contigs in short-read draft assemblies exploiting protein sequence-based replicon distribution scores. Microb. Genom. 2020, 6, e000398. [Google Scholar] [CrossRef]
- Hull, D.M.; Harrell, E.; van Vliet, A.H.M.; Correa, M.; Thakur, S. Antimicrobial resistance and interspecies gene transfer in Campylobacter coli and Campylobacter jejuni isolated from food animals, poultry processing, and retail meat in North Carolina, 2018–2019. PLoS ONE 2021, 16, e0246571. [Google Scholar] [CrossRef]
- Tedersoo, T.; Roasto, M.; Mäesaar, M.; Häkkinen, L.; Kisand, V.; Ivanova, M.; Valli, M.H.; Meremäe, K. Antibiotic Resistance in Campylobacter spp. Isolated from Broiler Chicken Meat and Human Patients in Estonia. Microorganisms 2022, 10, 1067. [Google Scholar] [CrossRef]
- Taylor, N.M.; Wales, A.D.; Ridley, A.M.; Davies, R.H. Farm level risk factors for fluoroquinolone resistance in E. coli and thermophilic Campylobacter spp. on poultry farms. Avian Pathol. 2016, 45, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Pereira, S.; Sahin, O.; Lin, J.; Huang, S.; Michel, L.; Zhang, Q. Enhanced in vivo fitness of fluoroquinolone-resistant Campylobacter jejuni in the absence of antibiotic selection pressure. Proc. Natl. Acad. Sci. USA 2005, 102, 541–546. [Google Scholar] [CrossRef]
- Kovač, J.; Čadež, N.; Stessl, B.; Stingl, K.; Gruntar, I.; Ocepek, M.; Etrkov, M.; Ewagner, M.; Možina, S. High genetic similarity of ciprofloxacin-resistant Campylobacter jejuni in central Europe. Front. Microbiol. 2015, 6, 1169. [Google Scholar] [CrossRef]
- Technical University of Denmark—National Food Institute. Comparative Genomics of Quinolone-Resistant and Susceptible Campylobacter jejuni of Poultry Origin from Major Poultry Producing European Countries (GENCAMP); EFSA Supporting Publication: Parma, Italy, 2018; EN-1398; p. 35. [Google Scholar] [CrossRef]
- Tenhagen, B.-A.; Flor, M.; Alt, K.; Knüver, M.-T.; Buhler, C.; Käsbohrer, A.; Stingl, K. Association of Antimicrobial Resistance in Campylobacter spp. in Broilers and Turkeys with Antimicrobial Use. Antibiotics 2021, 10, 673. [Google Scholar] [CrossRef]
- World Health Organization. Critically Important Antimicrobials for Human Medicine; 6th revision; Geneva, Switzerland, 2019; ISBN 978-92-4-151552-8. Available online: https://www.who.int/publications/i/item/9789241515528 (accessed on 7 September 2022).
- Mäesaar, M.; Praakle, K.; Meremäe, K.; Kramarenko, T.; Sõgel, J.; Viltrop, A.; Muutra, K.; Kovalenko, K.; Matt, D.; Hörman, A.; et al. Prevalence and counts of Campylobacter spp. in poultry meat at retail level in Estonia. Food Control 2014, 44, 72–77. [Google Scholar] [CrossRef]
- Nassiri, D.; Razavilar, V.; Motalebi, A. Detection of Campylobacter Species in Poultry Meat and Edible Offal’s In Western Azerbaijan Province. J. Pure Appl. Microbiol. 2016, 10, 2641–2646. [Google Scholar] [CrossRef]
- Ranjbar, R.; Babazadeh, D.; Jonaidi-Jafari, N. Prevalence of Campylobacter jejuni in adult patients with inflammatory bacterial diarrhea, East Azerbaijan, Iran. Acta Med. Mediterr. 2017, 33, 901–908. [Google Scholar] [CrossRef]
- Pergola, S.; Franciosini, M.; Comitini, F.; Ciani, M.; De Luca, S.; Bellucci, S.; Menchetti, L.; Proietti, P.C. Genetic diversity and antimicrobial resistance profiles of Campylobacter coli and Campylobacter jejuni isolated from broiler chicken in farms and at time of slaughter in central Italy. J. Appl. Microbiol. 2017, 122, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Jiang, Q.; Tang, H.; Wang, Z.; Yin, Y.; Ren, F.; Kong, L.; Jiao, X.; Huang, J. Characterization and Prevalence of Campylobacter spp. From Broiler Chicken Rearing Period to the Slaughtering Process in Eastern China. Front. Vet. Sci. 2020, 7, 227. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, Y.; Deng, F.; Liu, D.; Yao, H.; Zhang, Q.; Shen, J.; Liu, Z.; Gao, Y.; Wu, C.; et al. Species shift and multidrug resistance of Campylobacter from chicken and swine, China, 2008–2014. J. Antimicrob. Chemother. 2015, 71, 666–669. [Google Scholar] [CrossRef] [PubMed]
- Usha, M.R.; Fauziah, M.; Tunung, R.; Chai, L.C.; Cheah, Y.K.; Farinazleen, M.G.; Son, R. Occurrence and antibiotic resistance of Campylobacter jejuni and Campylobacter coli in retail broiler chicken. Int. Food Res. J. 2010, 17, 247–255. [Google Scholar]
- Mourkas, E.; Florez-Cuadrado, D.; Pascoe, B.; Calland, J.K.; Bayliss, S.C.; Mageiros, L.; Méric, G.; Hitchings, M.D.; Quesada, A.; Porrero, C.; et al. Gene pool transmission of multidrug resistance among Campylobacter from livestock, sewage and human disease. Environ. Microbiol. 2019, 21, 4597–4613. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Antimicrobial | ECOFF (μg/mL) (R>) | No. (%) of Resistant Isolates | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Backyard Chicken (n = 97) | Industrial Chicken (n = 50) | Human (n = 57) | Total (n = 204) | |||||||
| C. jejuni | C. coli | C. jejuni (n = 25) | C. coli (n = 72) | C. jejuni (n = 5) | C. coli (n = 45) | C. jejuni (n = 47) | C. coli (n = 10) | C. jejuni (n = 77) | C. coli (n = 127) | |
| Chloramphenicol | 16 | 16 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ciprofloxacin | 0.5 | 0.5 | 20 (80%) | 69 (96%) | 5 (100%) | 45 (100%) | 41 (87%) | 9 (90%) | 66 (86%) | 123 (97%) |
| Erythromycin | 4 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ertapenem | 0.5 | 0.5 | 0 | 27 (37%) | 0 | 37 (82%) | 0 | 6 (60%) | 0 | 70 |
| Gentamicin | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Tetracycline | 1 | 2 | 18 (72%) | 52 (72%) | 4 (80%) | 45 (100%) | 17 (36%) | 8 (80%) | 39 (51%) | 105 (83%) |
| Anti-Microbial | Covariate | Coefficient of Regression | Standard Error | Wald | Degrees of Freedom | p-Value | Odds Ratio | 95% Confidence Interval of Odds Ratio | Nagelkerke Pseudo R Squared | |
|---|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||||
| TET | Chicken vs. human | 1.153 | 0.403 | 8.203 | 1 | 0.004 | 3.167 | 1.439 | 6.971 | |
| C. coli vs. C. jejuni | 0.947 | 0.391 | 5.858 | 1 | 0.016 | 2.577 | 1.197 | 5.547 | 0.204 | |
| 2-3-fold resistance | Chicken vs. human | 1.361 | 0.442 | 9.487 | 1 | 0.002 | 3.901 | 1.641 | 9.276 | |
| C. coli vs. C. jejuni | 2.271 | 0.496 | 21.000 | 1 | <0.001 | 9.693 | 3.669 | 25.607 | 0.435 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metreveli, M.; Bulia, S.; Tevzadze, L.; Tsanava, S.; Zarske, M.; Goenaga, J.C.; Preuß, S.; Lomidze, G.; Koulouris, S.; Imnadze, P.; et al. Comparison of Antimicrobial Susceptibility Profiles of Thermotolerant Campylobacter spp. Isolated from Human and Poultry Samples in Georgia (Caucasus). Antibiotics 2022, 11, 1419. https://doi.org/10.3390/antibiotics11101419
Metreveli M, Bulia S, Tevzadze L, Tsanava S, Zarske M, Goenaga JC, Preuß S, Lomidze G, Koulouris S, Imnadze P, et al. Comparison of Antimicrobial Susceptibility Profiles of Thermotolerant Campylobacter spp. Isolated from Human and Poultry Samples in Georgia (Caucasus). Antibiotics. 2022; 11(10):1419. https://doi.org/10.3390/antibiotics11101419
Chicago/Turabian StyleMetreveli, Maia, Salome Bulia, Liana Tevzadze, Shota Tsanava, Michael Zarske, Juan Cruz Goenaga, Sandra Preuß, Giorgi Lomidze, Stylianos Koulouris, Paata Imnadze, and et al. 2022. "Comparison of Antimicrobial Susceptibility Profiles of Thermotolerant Campylobacter spp. Isolated from Human and Poultry Samples in Georgia (Caucasus)" Antibiotics 11, no. 10: 1419. https://doi.org/10.3390/antibiotics11101419
APA StyleMetreveli, M., Bulia, S., Tevzadze, L., Tsanava, S., Zarske, M., Goenaga, J. C., Preuß, S., Lomidze, G., Koulouris, S., Imnadze, P., & Stingl, K. (2022). Comparison of Antimicrobial Susceptibility Profiles of Thermotolerant Campylobacter spp. Isolated from Human and Poultry Samples in Georgia (Caucasus). Antibiotics, 11(10), 1419. https://doi.org/10.3390/antibiotics11101419