
Bacterial Mixology: Combining Pharmacodynamic Models to Predict In Vitro Competition of MCR-1-Harboring E. coli


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
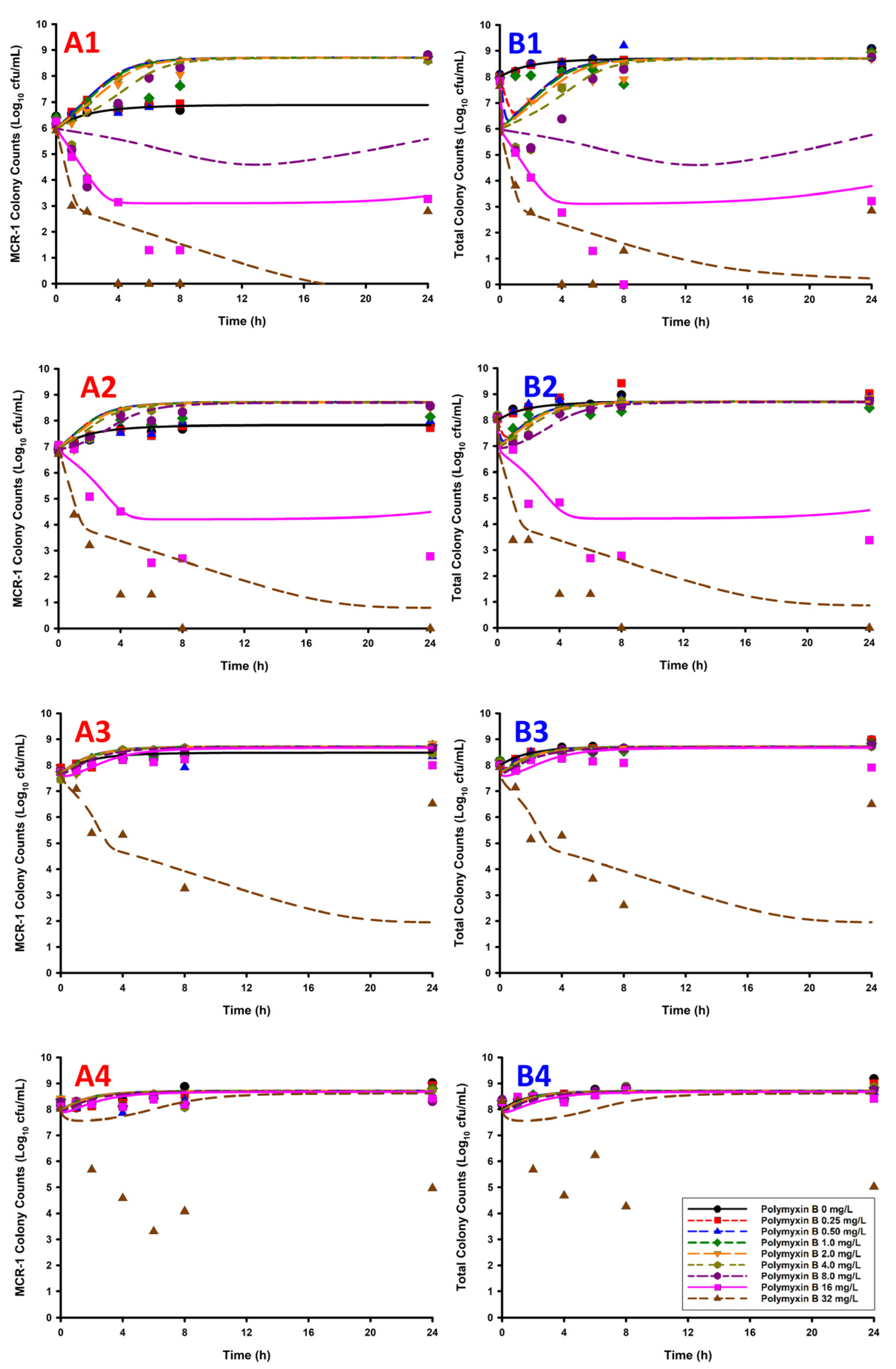
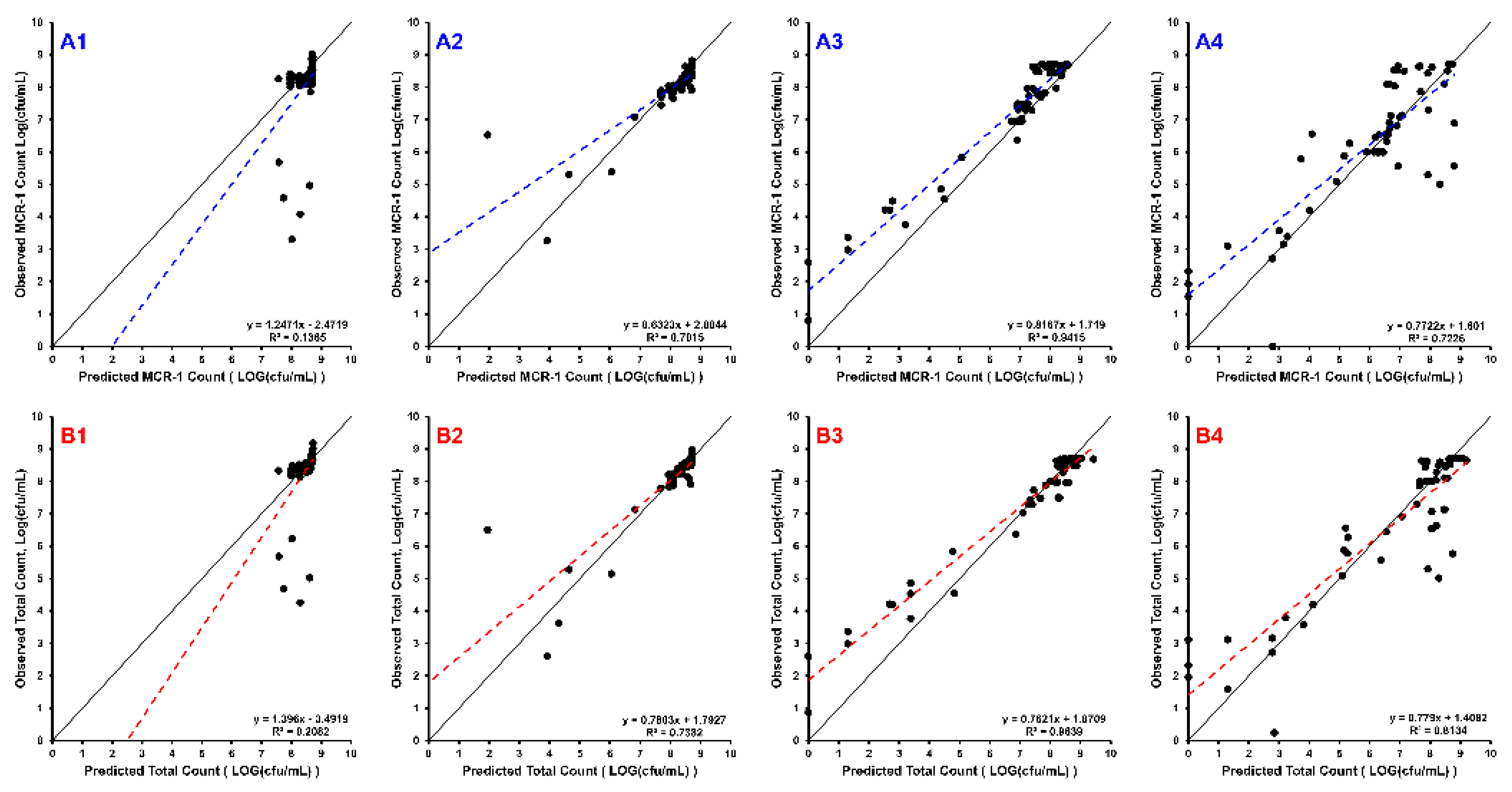
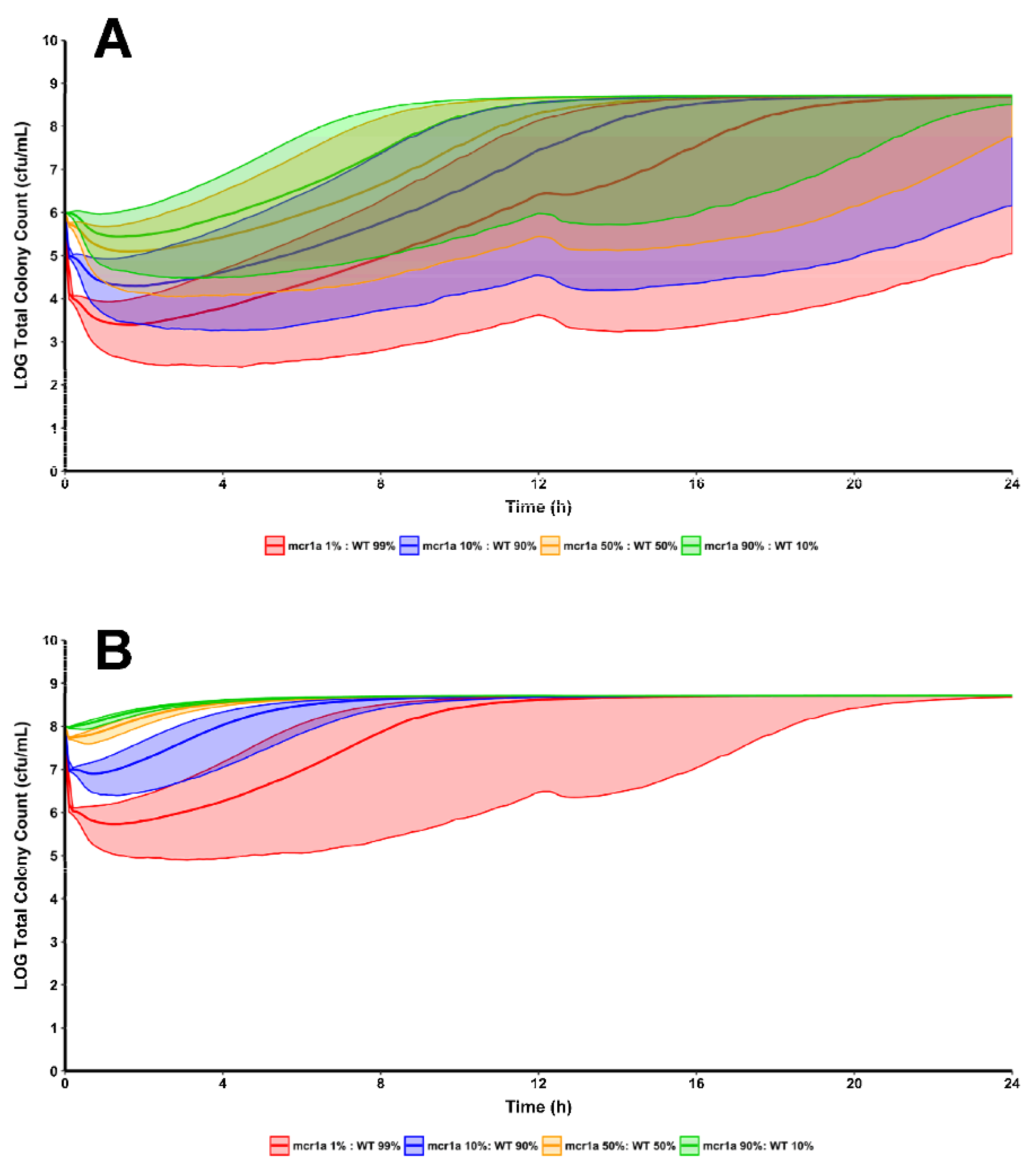
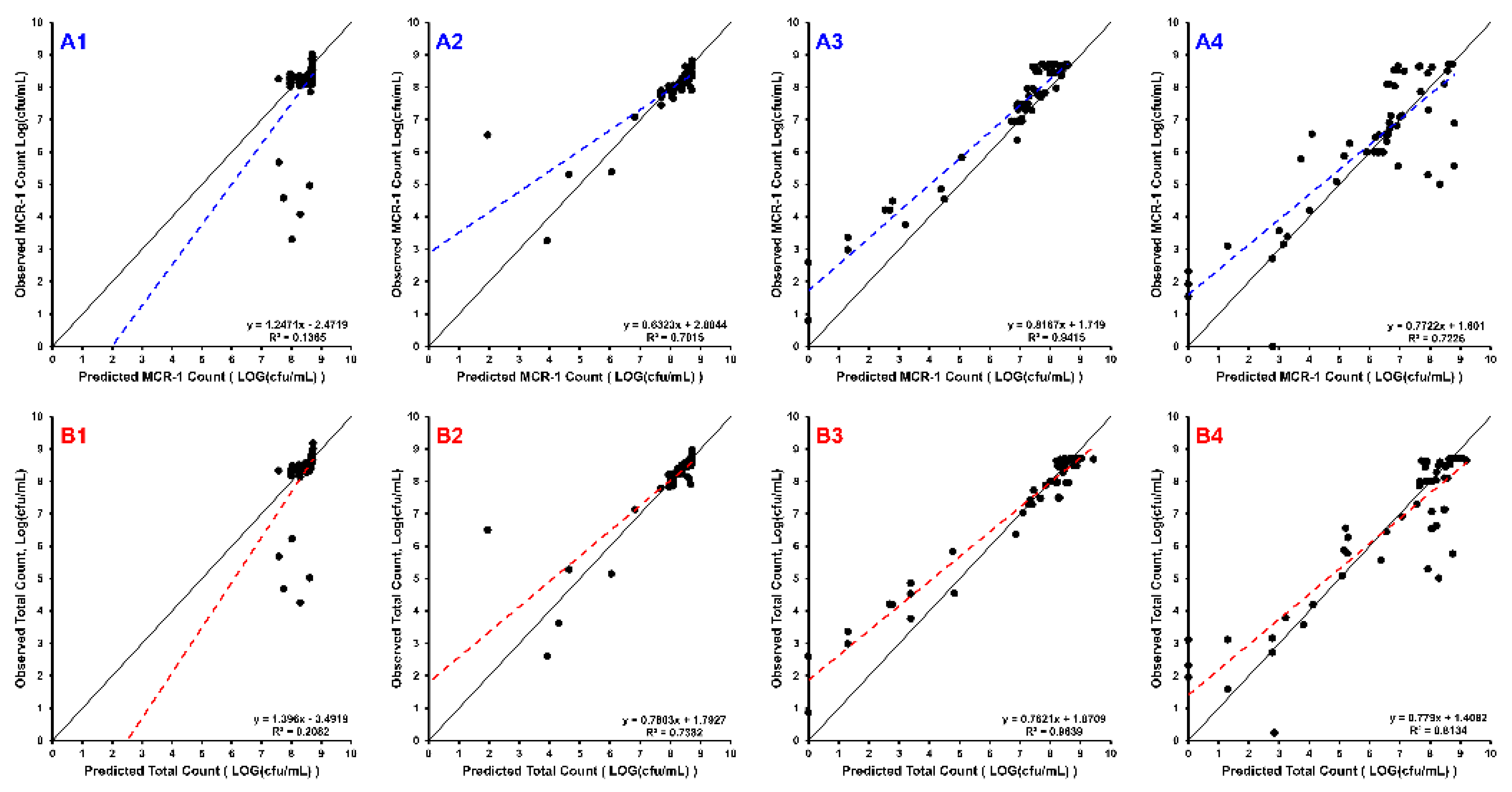
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates, Antibiotics, and Media
4.2. Mechanism-Based Model
4.3. Clinical Trial Simulation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Newly Reported Gene, mcr-1, Threatens Last-Resort Antibiotics. 2016. Available online: https://www.cdc.gov/drugresistance/solutions-initiative/stories/gene-reported-mcr.html (accessed on 20 November 2021).
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet. Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- McGann, P.; Snesrud, E.; Maybank, R.; Corey, B.; Ong, A.C.; Clifford, R.; Hinkle, M.; Whitman, T.; Lesho, E.; Schaecher, K.E. Escherichia coli Harboring mcr-1 and blaCTX-M on a Novel IncF Plasmid: First Report of mcr-1 in the United States. Antimicrob. Agents Chemother. 2016, 60, 4420–4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anyanwu, M.U.; Jaja, I.F.; Nwobi, O.C. Occurrence and Characteristics of Mobile Colistin Resistance (mcr) Gene-Containing Isolates from the Environment: A Review. Int. J. Environ. Res. Public Health 2020, 17, 1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nang, S.C.; Li, J.; Velkov, T. The rise and spread of mcr plasmid-mediated polymyxin resistance. Crit. Rev. Microbiol. 2019, 45, 131–161. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, H.; Liu, Y.H.; Feng, Y. Towards Understanding MCR-like Colistin Resistance. Trends Microbiol. 2018, 26, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Chen, L.; Tang, Y.W.; Kreiswirth, B.N. Emergence of the mcr-1 colistin resistance gene in carbapenem-resistant Enterobacteriaceae. Lancet Infect. Dis. 2016, 16, 287–288. [Google Scholar] [CrossRef] [Green Version]
- Olaitan, A.O.; Chabou, S.; Okdah, L.; Morand, S.; Rolain, J.M. Dissemination of the mcr-1 colistin resistance gene. Lancet Infect. Dis. 2016, 16, 147. [Google Scholar] [CrossRef] [Green Version]
- Olaitan, A.O.; Morand, S.; Rolain, J.M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meletis, G.; Skoura, L. Polymyxin Resistance Mechanisms: From Intrinsic Resistance to Mcr Genes. Recent Pat. Antiinfect. Drug Discov. 2018, 13, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.M.; Bulman, Z.P.; Sieron, A.O.; Bulitta, J.B.; Holden, P.N.; Nation, R.L.; Li, J.; Wright, G.D.; Tsuji, B.T. Pharmacodynamics of dose-escalated ‘front-loading’ polymyxin B regimens against polymyxin-resistant mcr-1-harbouring Escherichia coli. J. Antimicrob. Chemother. 2017, 72, 2297–2303. [Google Scholar] [CrossRef] [PubMed]
- Bulman, Z.P.; Satlin, M.J.; Chen, L.; Kreiswirth, B.N.; Shin, B.S.; Walsh, T.J.; Holden, P.N.; Forrest, A.; Nation, R.L.; Li, J.; et al. New Polymyxin B Dosing Strategies To Fortify Old Allies in the War against KPC-2-Producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2017, 61, e02023-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandri, A.M.; Landersdorfer, C.B.; Jacob, J.; Boniatti, M.M.; Dalarosa, M.G.; Falci, D.R.; Behle, T.F.; Bordinhao, R.C.; Wang, J.; Forrest, A.; et al. Population pharmacokinetics of intravenous polymyxin B in critically ill patients: Implications for selection of dosage regimens. Clin. Infect. Dis. 2013, 57, 524–531. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, B.T.; Landersdorfer, C.B.; Lenhard, J.R.; Cheah, S.E.; Thamlikitkul, V.; Rao, G.G.; Holden, P.N.; Forrest, A.; Bulitta, J.B.; Nation, R.L.; et al. Paradoxical Effect of Polymyxin B: High Drug Exposure Amplifies Resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 3913–3920. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; van Dorp, L.; Shaw, L.P.; Bradley, P.; Wang, Q.; Wang, X.; Jin, L.; Zhang, Q.; Liu, Y.; Rieux, A.; et al. The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nat. Commun. 2018, 9, 1179. [Google Scholar] [CrossRef] [Green Version]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilbäck, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, B.T.; Pogue, J.M.; Zavascki, A.P.; Paul, M.; Daikos, G.L.; Forrest, A.; Giacobbe, D.R.; Viscoli, C.; Giamarellou, H.; Karaiskos, I.; et al. International Consensus Guidelines for the Optimal Use of the Polymyxins: Endorsed by the American College of Clinical Pharmacy (ACCP), European Society of Clinical Microbiology and Infectious Diseases (ESCMID), Infectious Diseases Society of America (IDSA), International Society for Anti-infective Pharmacology (ISAP), Society of Critical Care Medicine (SCCM), and Society of Infectious Diseases Pharmacists (SIDP). Pharmacotherapy 2019, 39, 10–39. [Google Scholar]
- Feng, Y. Transferability of MCR-1/2 Polymyxin Resistance: Complex Dissemination and Genetic Mechanism. ACS Infect. Dis. 2018, 4, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.M.; Lenhard, J.R.; Boissonneault, K.R.; Landersdorfer, C.B.; Bulitta, J.B.; Holden, P.N.; Forrest, A.; Nation, R.L.; Li, J.; Tsuji, B.T. Using machine learning to optimize antibiotic combinations: Dosing strategies for meropenem and polymyxin B against carbapenem-resistant Acinetobacter baumannii. Clin. Microbiol. Infect. 2020, 26, 1207–1213. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, N.M.; Chan, A.; Nguyen, T.D.; Dumbleton, J.T. Bacterial Mixology: Combining Pharmacodynamic Models to Predict In Vitro Competition of MCR-1-Harboring E. coli. Antibiotics 2022, 11, 34. https://doi.org/10.3390/antibiotics11010034
Smith NM, Chan A, Nguyen TD, Dumbleton JT. Bacterial Mixology: Combining Pharmacodynamic Models to Predict In Vitro Competition of MCR-1-Harboring E. coli. Antibiotics. 2022; 11(1):34. https://doi.org/10.3390/antibiotics11010034
Chicago/Turabian StyleSmith, Nicholas M., Arthur Chan, Thomas D. Nguyen, and Jacob T. Dumbleton. 2022. "Bacterial Mixology: Combining Pharmacodynamic Models to Predict In Vitro Competition of MCR-1-Harboring E. coli" Antibiotics 11, no. 1: 34. https://doi.org/10.3390/antibiotics11010034
APA StyleSmith, N. M., Chan, A., Nguyen, T. D., & Dumbleton, J. T. (2022). Bacterial Mixology: Combining Pharmacodynamic Models to Predict In Vitro Competition of MCR-1-Harboring E. coli. Antibiotics, 11(1), 34. https://doi.org/10.3390/antibiotics11010034