
Air Ambulance: Antimicrobial Power of Bacterial Volatiles



Abstract
:1. Antimicrobial Resistance: A Global Crisis
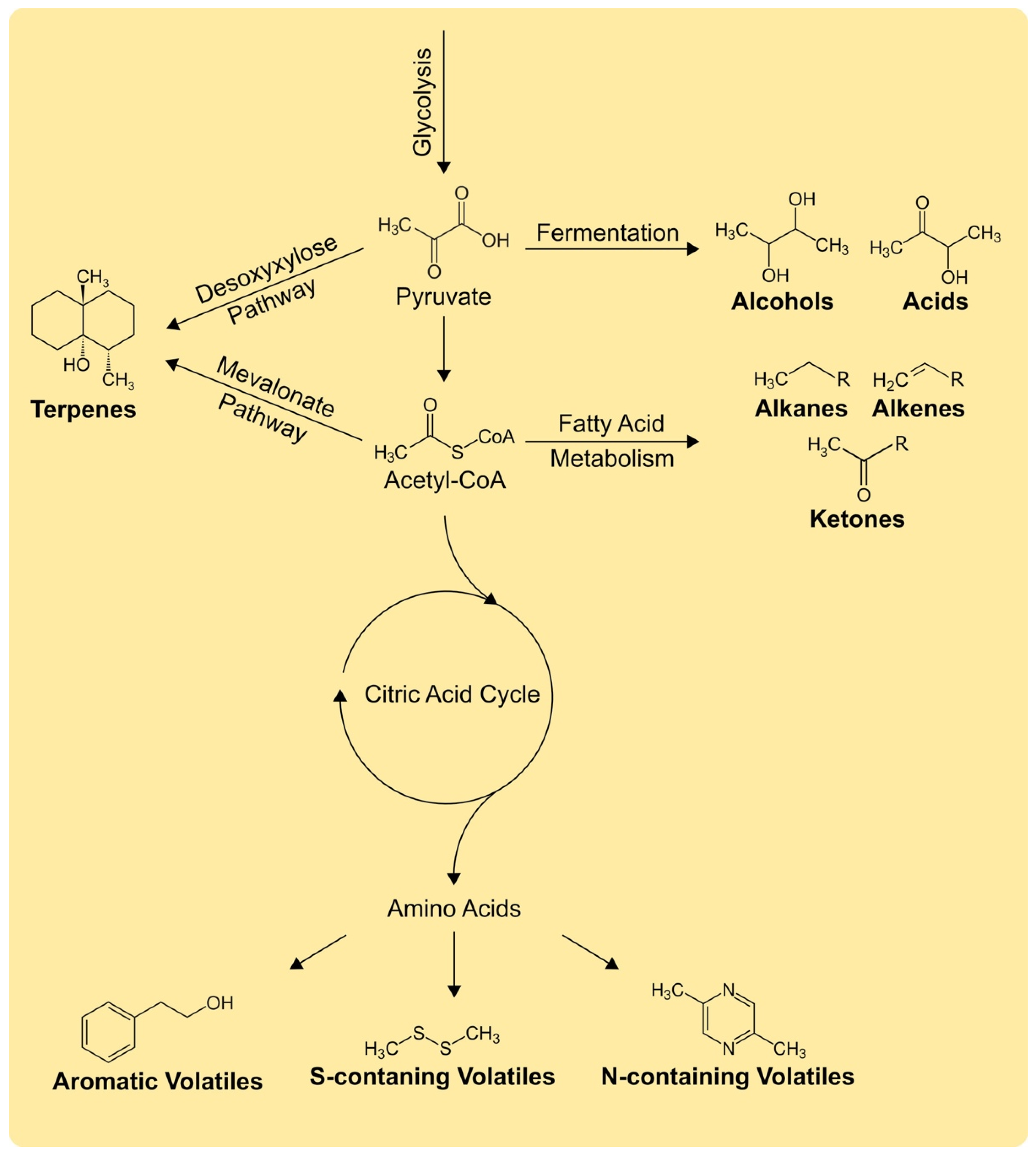
2. Biochemistry of Volatiles: Diverse and Diffusive
3. Bacterial Antimicrobial Volatiles: An Overview
3.1. Sulfur-Containing Volatiles
3.2. Bacillus and Streptomyces as Volatile-Producers
3.3. Co-Cultivation and Volatile Blends
3.4. Modes of Action and Abiotic Factors
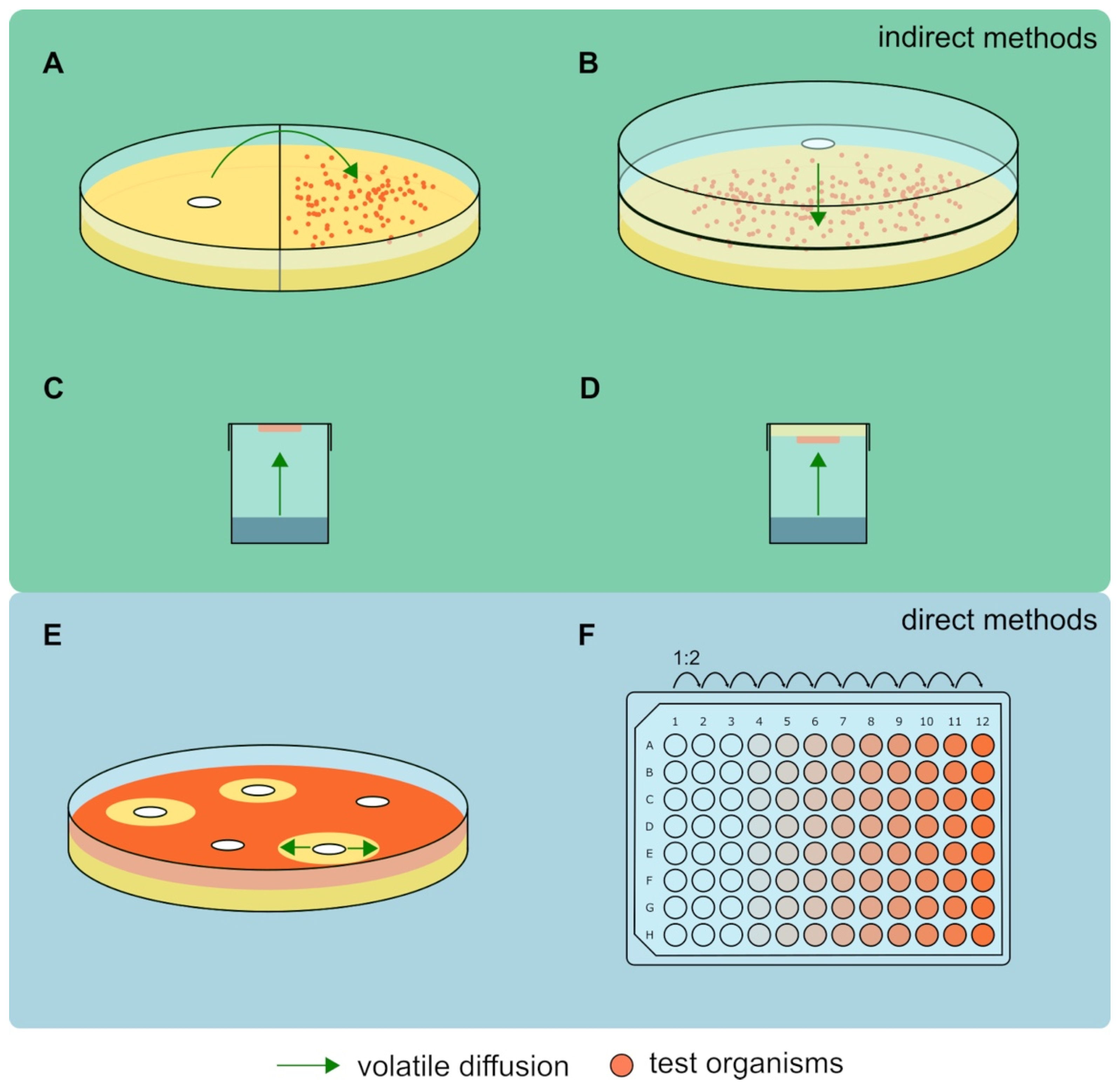
4. Antimicrobial Activity of Volatiles: The Testing Methods
5. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dodds, D.R. Antibiotic Resistance: A Current Epilogue. Biochem. Pharmacol. 2017, 134, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S.; Buss, A.D. Natural Products-The Future Scaffolds for Novel Antibiotics? Biochem. Pharmacol. 2006, 71, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Rolain, J.-M.; Abat, C.; Jimeno, M.-T.; Fournier, P.-E.; Raoult, D. Do We Need New Antibiotics? Clin. Microbiol. Infect. 2016, 22, 408–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casewell, M.; Hill, R. In-Vitro Activity of Mupirocin (‘Pseudomonic Acid’) against Clinical Isolates of Staphylococcus aureus. J. Antimicrob. Chemother. 1985, 15, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, R.; Boon, R.; Griffin, K.; Masters, P.; Slocombe, B.; White, A. Antibacterial Activity of Mupirocin (Pseudomonic Acid), a New Antibiotic for Topical Use. Antimicrob. Agents Chemother. 1985, 27, 495–498. [Google Scholar] [CrossRef] [Green Version]
- Clemett, D.; Markham, A. Linezolid. Drugs 2000, 59, 815–827. [Google Scholar] [CrossRef] [PubMed]
- LaPlante, K.L.; Rybak, M.J. Daptomycin–a Novel Antibiotic against Gram-Positive Pathogens. Expert Opin. Pharmacother. 2004, 5, 2321–2331. [Google Scholar] [CrossRef]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schäberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A New Antibiotic Kills Pathogens without Detectable Resistance. Nature 2015, 517, 455–459. [Google Scholar] [CrossRef]
- Fleming, A. Penicillin. Available online: https://www.nobelprize.org/prizes/medicine/1945/fleming/lecture/ (accessed on 23 November 2021).
- Michael, C.A.; Dominey-Howes, D.; Labbate, M. The Antimicrobial Resistance Crisis: Causes, Consequences, and Management. Front. Public Health 2014, 2, 145. [Google Scholar] [CrossRef]
- WHO. No Time to Wait: Securing the Future from Drug-Resistant Infections. Available online: https://www.who.int/publications/i/item/no-time-to-wait-securing-the-future-from-drug-resistant-infections (accessed on 2 November 2021).
- Hwang, A.Y.; Gums, J.G. The Emergence and Evolution of Antimicrobial Resistance: Impact on a Global Scale. Bioorg. Med. Chem. 2016, 24, 6440–6445. [Google Scholar] [CrossRef]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Available online: https://amr-review.org/Publications.html (accessed on 11 October 2021).
- Banin, E.; Hughes, D.; Kuipers, O.P. Editorial: Bacterial Pathogens, Antibiotics and Antibiotic Resistance. FEMS Microbiol. Rev. 2017, 41, 450–452. [Google Scholar] [CrossRef] [Green Version]
- Getahun, H.; Smith, I.; Trivedi, K.; Paulin, S.; Balkhy, H.H. Tackling Antimicrobial Resistance in the COVID-19 Pandemic. Bull. World Health Organ. 2020, 98, 442. [Google Scholar] [CrossRef]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.W.; Harper, D.; et al. Alternatives to Antibiotics—A Pipeline Portfolio Review. Lancet Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Coates, A.R.; Halls, G.; Hu, Y. Novel Classes of Antibiotics or More of the Same? Br. J. Pharmacol. 2011, 163, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Avalos, M.; van Wezel, G.P.; Raaijmakers, J.M.; Garbeva, P. Healthy Scents: Microbial Volatiles as New Frontier in Antibiotic Research? Curr. Opin. Microbiol. 2018, 45, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, H.; Hassane, A.; Rawway, M.; El-Sayed, M.; Gomaa, A.E.-R.; Abdul-Raouf, U.; Shah, A.M.; Abdelmotaal, H.; Song, Y. Antibacterial and Cytotoxic Potency of Thermophilic Streptomyces Werraensis MI-S.24-3 Isolated from an Egyptian Extreme Environment. Arch. Microbiol. 2021, 203, 4961–4972. [Google Scholar] [CrossRef] [PubMed]
- Nas, F.; Aissaoui, N.; Mahjoubi, M.; Mosbah, A.; Arab, M.; Abdelwahed, S.; Khrouf, R.; Masmoudi, A.-S.; Cherif, A.; Klouche-Khelil, N. A Comparative GC-MS Analysis of Bioactive Secondary Metabolites Produced by Halotolerant Bacillus spp. Isolated from the Great Sebkha of Oran. Int. Microbiol. 2021, 24, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Tyc, O.; de Jager, V.C.L.; van den Berg, M.; Gerards, S.; Janssens, T.K.S.; Zaagman, N.; Kai, M.; Svatos, A.; Zweers, H.; Hordijk, C.; et al. Exploring Bacterial Interspecific Interactions for Discovery of Novel Antimicrobial Compounds. Microb. Biotechnol. 2017, 10, 910–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, B.; Lin, X.; Lei, L.; Lamb, D.C.; Kelly, S.L.; Waterman, M.R.; Cane, D.E. Biosynthesis of the Sesquiterpene Antibiotic Albaflavenone in Streptomyces Coelicolor A3(2). J. Biol. Chem. 2008, 283, 8183–8189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowan, D.D. Volatile Metabolites. Metabolites 2011, 1, 41–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, R.; Cordovez, V.; de Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile Affairs in Microbial Interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [Green Version]
- Schulz-Bohm, K.; Martin-Sanchez, L.; Garbeva, P. Microbial Volatiles: Small Molecules with an Important Role in Intra- and Inter-Kingdom Interactions. Front. Microbiol. 2017, 8, 2484. [Google Scholar] [CrossRef]
- Weisskopf, L.; Schulz, S.; Garbeva, P. Microbial Volatile Organic Compounds in Intra-Kingdom and Inter-Kingdom Interactions. Nat. Rev. Microbiol. 2021, 19, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Ossowicki, A.; Jafra, S.; Garbeva, P. The Antimicrobial Volatile Power of the Rhizospheric Isolate Pseudomonas donghuensis P482. PLoS ONE 2017, 12, e0174362. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Henderson, G.; Grimm, C.C.; Lloyd, S.W.; Laine, R.A. Termites Fumigate Their Nests with Naphthalene. Nature 1998, 392, 558–559. [Google Scholar] [CrossRef]
- Lammers, A.; Zweers, H.; Sandfeld, T.; Bilde, T.; Garbeva, P.; Schramm, A.; Lalk, M. Antimicrobial Compounds in the Volatilome of Social Spider Communities. Front. Microbiol. 2021, 12, 2434. [Google Scholar] [CrossRef]
- Garbeva, P.; Hordijk, C.; Gerards, S.; de Boer, W. Volatile-Mediated Interactions between Phylogenetically Different Soil Bacteria. Front. Microbiol. 2014, 5, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Garbeva, P.; Liu, X.; Gunnewiek, P.J.A.K.; Clocchiatti, A.; Hundscheid, M.P.J.; Wang, X.; de Boer, W. Volatile-Mediated Antagonism of Soil Bacterial Communities against Fungi. Environ. Microbiol. 2020, 22, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Schulz, S.; Dickschat, J.S. Bacterial Volatiles: The Smell of Small Organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef]
- Peñuelas, J.; Asensio, D.; Tholl, D.; Wenke, K.; Rosenkranz, M.; Piechulla, B.; Schnitzler, J.p. Biogenic Volatile Emissions from the Soil. Plant Cell Environ. 2014, 37, 1866–1891. [Google Scholar] [CrossRef]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Wei, H.X.; Pare, P.W.; Kloepper, J.W. Bacterial Volatiles Promote Growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [Green Version]
- Hammerbacher, A.; Coutinho, T.A.; Gershenzon, J. Roles of Plant Volatiles in Defence against Microbial Pathogens and Microbial Exploitation of Volatiles. Plant Cell Environ. 2019, 42, 2827–2843. [Google Scholar] [CrossRef] [Green Version]
- Veselova, M.A.; Plyuta, V.A.; Khmel, I.A. Volatile Compounds of Bacterial Origin: Structure, Biosynthesis, and Biological Activity. Microbiology 2019, 88, 261–274. [Google Scholar] [CrossRef]
- Buhaescu, I.; Izzedine, H. Mevalonate Pathway: A Review of Clinical and Therapeutical Implications. Clin. Biochem. 2007, 40, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Rohdich, F.; Zepeck, F.; Adam, P.; Hecht, S.; Kaiser, J.; Laupitz, R.; Gräwert, T.; Amslinger, S.; Eisenreich, W.; Bacher, A.; et al. The Deoxyxylulose Phosphate Pathway of Isoprenoid Biosynthesis: Studies on the Mechanisms of the Reactions Catalyzed by IspG and IspH Protein. Proc. Natl. Acad. Sci. USA 2003, 100, 1586–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avalos, M.; Garbeva, P.; Raaijmakers, J.M.; van Wezel, G.P. Production of Ammonia as a Low-Cost and Long-Distance Antibiotic Strategy by Streptomyces Species. ISME J. 2019, 14, 569–583. [Google Scholar] [CrossRef]
- Bernier, S.P.; Letoffe, S.; Delepierre, M.; Ghigo, J.-M. Biogenic Ammonia Modifies Antibiotic Resistance at a Distance in Physically Separated Bacteria. Mol. Microbiol. 2011, 81, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Calvo, H.; Mendiara, I.; Arias, E.; Gracia, A.P.; Blanco, D.; Eugenia Venturini, M. Antifungal Activity of the Volatile Organic Compounds Produced by Bacillus velezensis Strains against Postharvest Fungal Pathogens. Postharvest Biol. Technol. 2020, 166, 111208. [Google Scholar] [CrossRef]
- Rybakova, D.; Rack-Wetzlinger, U.; Cernava, T.; Schaefer, A.; Schmuck, M.; Berg, G. Aerial Warfare: A Volatile Dialogue between the Plant Pathogen Verticillium longisporum and Its Antagonist Paenibacillus polymyxa. Front. Plant Sci. 2017, 8, 1294. [Google Scholar] [CrossRef] [Green Version]
- Garrido, A.; Atencio, L.A.; Bethancourt, R.; Bethancourt, A.; Guzmán, H.; Gutiérrez, M.; Durant-Archibold, A.A. Antibacterial Activity of Volatile Organic Compounds Produced by the Octocoral-Associated Bacteria Bacillus sp. BO53 and Pseudoalteromonas sp. GA327. Antibiotics 2020, 9, 923. [Google Scholar] [CrossRef]
- Bruisson, S.; Zufferey, M.; L’Haridon, F.; Trutmann, E.; Anand, A.; Dutartre, A.; De Vrieze, M.; Weisskopf, L. Endophytes and Epiphytes From the Grapevine Leaf Microbiome as Potential Biocontrol Agents against Phytopathogens. Front. Microbiol. 2019, 10, 2726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssens, T.K.S.; Tyc, O.; Besselink, H.; de Boer, W.; Garbeva, P. Biological Activities Associated with the Volatile Compound 2,5-bis(1-Methylethyl)-pyrazine. Fems Microbiol. Lett. 2019, 366, fnz023. [Google Scholar] [CrossRef] [Green Version]
- Schulz, S.; Dickschat, J.S.; Kunze, B.; Wagner-Dobler, I.; Diestel, R.; Sasse, F. Biological Activity of Volatiles from Marine and Terrestrial Bacteria. Mar. Drugs 2010, 8, 2976–2987. [Google Scholar] [CrossRef] [PubMed]
- Gotor-Vila, A.; Teixidó, N.; Di Francesco, A.; Usall, J.; Ugolini, L.; Torres, R.; Mari, M. Antifungal Effect of Volatile Organic Compounds Produced by Bacillus amyloliquefaciens CPA-8 against Fruit Pathogen Decays of Cherry. Food Microbiol. 2017, 64, 219–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.; Park, D.; Kim, K.; Lim, S.M.; Yu, N.H.; Kim, S.; Kim, H.-Y.; Jung, K.S.; Jang, J.Y.; Park, J.-C.; et al. Characterization of Bacillus amyloliquefaciens DA12 Showing Potent Antifungal Activity against Mycotoxigenic Fusarium Species. Plant Pathol. J. 2017, 33, 499–507. [Google Scholar] [CrossRef] [Green Version]
- Tahir, H.A.S.; Gu, Q.; Wu, H.; Niu, Y.; Huo, R.; Gao, X. Bacillus Volatiles Adversely Affect the Physiology and Ultra-Structure of Ralstonia Solanacearum and Induce Systemic Resistance in Tobacco against Bacterial Wilt. Sci. Rep. 2017, 7, 40481. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, J.; Li, C.; Ma, Y. Antifungal and Plant Growth Promotion Activity of Volatile Organic Compounds Produced by Bacillus amyloliquefaciens. MicrobiologyOpen 2019, 8, e00813. [Google Scholar] [CrossRef] [Green Version]
- Mannaa, M.; Kim, K.D. Biocontrol Activity of Volatile-Producing Bacillus megaterium and Pseudomonas protegens against Aspergillus and Penicillium spp. Predominant in Stored Rice Grains: Study II. Mycobiology 2018, 46, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Morita, T.; Tanaka, I.; Ryuda, N.; Ikari, M.; Ueno, D.; Someya, T. Antifungal Spectrum Characterization and Identification of Strong Volatile Organic Compounds Produced by Bacillus pumilus TM-R. Heliyon 2019, 5, e01817. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Li, M.; Wang, D.; Wang, F.; Shen, H.; Sun, G.; Feng, C.; Wang, X.; Chen, D.; Sun, X. Biocontrol Efficacy of Bacillus siamensis LZ88 against Brown Spot Disease of Tobacco Caused by Alternaria alternata. Biol. Control 2021, 154, 104508. [Google Scholar] [CrossRef]
- Ezrari, S.; Mhidra, O.; Radouane, N.; Tahiri, A.; Polizzi, G.; Lazraq, A.; Lahlali, R. Potential Role of Rhizobacteria Isolated from Citrus Rhizosphere for Biological Control of Citrus Dry Root Rot. Plants 2021, 10, 872. [Google Scholar] [CrossRef]
- Massawe, V.C.; Hanif, A.; Farzand, A.; Mburu, D.K.; Ochola, S.O.; Wu, L.; Tahir, H.A.S.; Gu, Q.; Wu, H.; Gao, X. Volatile Compounds of Endophytic Bacillus spp. Have Biocontrol Activity against Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1373–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.; Zang, H.; Wu, H.; Rajer, F.U.; Gao, X. Antibacterial Effects of Volatiles Produced by Bacillus Strain D13 against Xanthomonas Oryzae Pv. Oryzae. Mol. Plant Pathol. 2018, 19, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Osaki, C.; Yamaguchi, K.; Urakawa, S.; Nakashima, Y.; Sugita, K.; Nagaishi, M.; Mitsuiki, S.; Kuraoka, T.; Ogawa, Y.; Sato, H. The Bacteriological Properties of Bacillus Strain TM-I-3 and Analysis of the Volatile Antifungal Compounds Emitted by This Bacteria. Biocontrol Sci. 2019, 24, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajer, F.U.; Wu, H.; Xie, Y.; Xie, S.; Raza, W.; Tahir, H.A.S.; Gao, X. Volatile Organic Compounds Produced by a Soil-Isolate, Bacillus subtilis FA26 Induce Adverse Ultra-Structural Changes to the Cells of Clavibacter michiganensis ssp. sepedonicus, the Causal Agent of Bacterial Ring Rot of Potato. Microbiology 2017, 163, 523–530. [Google Scholar] [CrossRef]
- Mu, J.; Li, X.; Jiao, J.; Ji, G.; Wu, J.; Hu, F.; Li, H. Biocontrol Potential of Vermicompost through Antifungal Volatiles Produced by Indigenous Bacteria. Biol. Control 2017, 112, 49–54. [Google Scholar] [CrossRef]
- Lim, S.M.; Yoon, M.-Y.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Shin, T.S.; Park, H.W.; Yu, N.H.; Kim, Y.H.; Kim, J.-C. Diffusible and Volatile Antifungal Compounds Produced by an Antagonistic Bacillus velezensis G341 against Various Phytopathogenic Fungi. Plant Pathol. J. 2017, 33, 488–498. [Google Scholar] [CrossRef] [Green Version]
- Eida, A.A.; Bougouffa, S.; L’Haridon, F.; Alam, I.; Weisskopf, L.; Bajic, V.B.; Saad, M.M.; Hirt, H. Genome Insights of the Plant-Growth Promoting Bacterium Cronobacter muytjensii JZ38 with Volatile-Mediated Antagonistic Activity against Phytophthora infestans. Front. Microbiol. 2020, 11, 369. [Google Scholar] [CrossRef] [Green Version]
- Juarez, G.E.; Mateyca, C.; Galvan, E.M. Proteus mirabilis Outcompetes Klebsiella pneumoniae in Artificial Urine Medium through Secretion of Ammonia and Other Volatile Compounds. Heliyon 2020, 6, e03361. [Google Scholar] [CrossRef]
- Tagele, S.B.; Lee, H.G.; Kim, S.W.; Lee, Y.S. Phenazine and 1-Undecene Producing Pseudomonas Chlororaphis Subsp. Aurantiaca Strain KNU17Pc1 for Growth Promotion and Disease Suppression in Korean Maize Cultivars. Environ. Microbiol. Biotechnol. 2019, 29, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, T.; Liu, Y.; Li, X.; Zhang, C.; Feng, Z.; Peng, X.; Li, Z.; Qin, S.; Xing, K. Volatile Organic Compounds Produced by Pseudomonas chlororaphis subsp. aureofaciens SPS-41 as Biological Fumigants to Control Ceratocystis fimbriata in Postharvest Sweet Potatoes. J. Agric. Food Chem. 2019, 67, 3702–3710. [Google Scholar] [CrossRef] [PubMed]
- Prigigallo, M.I.; De Stradis, A.; Anand, A.; Mannerucci, F.; L’Haridon, F.; Weisskopf, L.; Bubici, G. Basidiomycetes Are Particularly Sensitive to Bacterial Volatile Compounds: Mechanistic Insight Into the Case Study of Pseudomonas protegens Volatilome against Heterobasidion abietinum. Front. Microbiol. 2021, 12, 684664. [Google Scholar] [CrossRef] [PubMed]
- Archana, T.J.; Gogoi, R.; Kaur, C.; Varghese, E.; Sharma, R.R.; Srivastav, M.; Tomar, M.; Kumar, M.; Kumar, A. Bacterial Volatile Mediated Suppression of Postharvest Anthracnose and Quality Enhancement in Mango. Postharvest Biol. Technol. 2021, 177, 111525. [Google Scholar] [CrossRef]
- Agisha, V.N.; Kumar, A.; Eapen, S.J.; Sheoran, N.; Suseelabhai, R. Broad-Spectrum Antimicrobial Activity of Volatile Organic Compounds from Endophytic Pseudomonas putida BP25 against Diverse Plant Pathogens. Biocontrol Sci. Technol. 2019, 29, 1069–1089. [Google Scholar] [CrossRef]
- Rojas-Solis, D.; Zetter-Salmon, E.; Contreras-Perez, M.; del Carmen Rocha-Granados, M.; Macias-Rodriguez, L.; Santoyo, G. Pseudomonas stutzeri E25 and Stenotrophomonas maltophilia CR71 Endophytes Produce Antifungal Volatile Organic Compounds and Exhibit Additive Plant Growth-Promoting Effects. Biocatal. Agric. Biotechnol. 2018, 13, 46–52. [Google Scholar] [CrossRef]
- Yang, M.; Lu, L.; Pang, J.; Hu, Y.; Guo, Q.; Li, Z.; Wu, S.; Liu, H.; Wang, C. Biocontrol Activity of Volatile Organic Compounds from Streptomyces alboflavus TD-1 against Aspergillus flavus Growth and Aflatoxin Production. J. Microbiol. 2019, 57, 396–404. [Google Scholar] [CrossRef]
- Xing, M.; Zheng, L.; Deng, Y.; Xu, D.; Xi, P.; Li, M.; Kong, G.; Jiang, Z. Antifungal Activity of Natural Volatile Organic Compounds against Litchi Downy Blight Pathogen Peronophythora litchii. Molecules 2018, 23, 358. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, B.; Cai, S.; Zhang, Y.; Xu, M.; Zhang, C.; Yuan, B.; Xing, K.; Qin, S. Identification of Rhizospheric Actinomycete Streptomyces lavendulae SPS-33 and the Inhibitory Effect of Its Volatile Organic Compounds against Ceratocystis fimbriata in Postharvest Sweet Potato (Ipomoea batatas (L.) Lam.). Microorganisms 2020, 8, 319. [Google Scholar] [CrossRef] [Green Version]
- Lyu, A.; Yang, L.; Wu, M.; Zhang, J.; Li, G. High Efficacy of the Volatile Organic Compounds of Streptomyces yanglinensis 3-10 in Suppression of Aspergillus Contamination on Peanut Kernels. Front. Microbiol. 2020, 11, 142. [Google Scholar] [CrossRef]
- Chacon-Orozco, J.G.; Bueno, C.J.; Shapiro-Ilan, D.I.; Hazir, S.; Leite, L.G.; Harakava, R. Antifungal Activity of Xenorhabdus spp. and Photorhabdus spp. against the Soybean Pathogenic Sclerotinia sclerotiorum. Sci. Rep. 2020, 10, 20649. [Google Scholar] [CrossRef]
- Dandurishvili, N.; Toklikishvili, N.; Ovadis, M.; Eliashvili, P.; Giorgobiani, N.; Keshelava, R.; Tediashvili, M.; Vainstein, A.; Khmel, I.; Szegedi, E.; et al. Broad-Range Antagonistic Rhizobacteria Pseudomonas fluorescens and Serratia plymuthica Suppress Agrobacterium Crown Gall Tumours on Tomato Plants. J. Appl. Microbiol. 2010, 110, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Tyc, O.; Zweers, H.; de Boer, W.; Garbeva, P. Volatiles in Inter-Specific Bacterial Interactions. Front. Microbiol. 2015, 6, 1412. [Google Scholar] [CrossRef] [Green Version]
- Watve, M.G.; Tickoo, R.; Jog, M.M.; Bhole, B.D. How Many Antibiotics Are Produced by the Genus Streptomyces? Arch. Microbiol. 2001, 176, 386–390. [Google Scholar] [CrossRef]
- Gürtler, H.; Pedersen, R.; Anthoni, U.; Christophersen, C.; Nielsen, P.H.; Wellington, E.M.; Pedersen, C.; Bock, K. Albaflavenone, a Sesquiterpene Ketone with a Zizaene Skeleton Produced by a Streptomycete with a New Rope Morphology. J. Antibiot. 1994, 47, 434–439. [Google Scholar] [CrossRef] [Green Version]
- Moody, S.C.; Zhao, B.; Lei, L.; Nelson, D.R.; Mullins, J.G.L.; Waterman, M.R.; Kelly, S.L.; Lamb, D.C. Investigating Conservation of the Albaflavenone Biosynthetic Pathway and CYP170 Bifunctionality in Streptomycetes. FEBS J. 2012, 279, 1640–1649. [Google Scholar] [CrossRef]
- Takamatsu, S.; Lin, X.; Nara, A.; Komatsu, M.; Cane, D.E.; Ikeda, H. Characterization of a Silent Sesquiterpenoid Biosynthetic Pathway in Streptomyces avermitilis Controlling Epi-Isozizaene Albaflavenone Biosynthesis and Isolation of a New Oxidized Epi-Isozizaene Metabolite. Microb. Biotechnol. 2011, 4, 184–191. [Google Scholar] [CrossRef]
- Braña, A.F.; Rodríguez, M.; Pahari, P.; Rohr, J.; García, L.A.; Blanco, G. Activation and Silencing of Secondary Metabolites in Streptomyces albus and Streptomyces lividans after Transformation with Cosmids Containing the Thienamycin Gene Cluster from Streptomyces cattleya. Arch. Microbiol. 2014, 196, 345–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Yuan, J.E.Y.; Raza, W.; Shen, Q.; Huang, Q. Effects of Volatile Organic Compounds from Streptomyces albulus NJZJSA2 on Growth of Two Fungal Pathogens. J. Basic Microbiol. 2015, 55, 1104–1117. [Google Scholar] [CrossRef] [PubMed]
- Schulz-Bohm, K.; Zweers, H.; de Boer, W.; Garbeva, P. A Fragrant Neighborhood: Volatile Mediated Bacterial Interactions in Soil. Front. Microbiol. 2015, 6, 1212. [Google Scholar] [CrossRef] [Green Version]
- Kai, M.; Effmert, U.; Lemfack, M.C.; Piechulla, B. Interspecific Formation of the Antimicrobial Volatile Schleiferon. Sci. Rep. 2018, 8, 16852. [Google Scholar] [CrossRef]
- Abis, L.; Loubet, B.; Ciuraru, R.; Lafouge, F.; Houot, S.; Nowak, V.; Tripied, J.; Dequiedt, S.; Maron, P.A.; Sadet-Bourgeteau, S. Reduced Microbial Diversity Induces Larger Volatile Organic Compound Emissions from Soils. Sci. Rep. 2020, 10, 6104. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Schmidt, R.; de Jager, V.; Krzyzanowska, D.; Jongedijk, E.; Cankar, K.; Beekwilder, J.; van Veen, A.; de Boer, W.; van Veen, J.A.; et al. Exploring the Genomic Traits of Fungus-Feeding Bacterial Genus Collimonas. BMC Genom. 2015, 16, 1103. [Google Scholar] [CrossRef]
- Chernin, L.; Toklikishvili, N.; Ovadis, M.; Kim, S.; Ben-Ari, J.; Khmel, I.; Vainstein, A. Quorum-Sensing Quenching by Rhizobacterial Volatiles. Environ. Microbiol. Rep. 2011, 3, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, D.; Castelli, F.; Sarpietro, M.G.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of Antibacterial Action of Three Monoterpenes. Antimicrob. Agents Chemother. 2005, 49, 2474–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbeva, P.; Weisskopf, L. Airborne Medicine: Bacterial Volatiles and Their Influence on Plant Health. New Phytol. 2019, 226, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Garbeva, P.; Hordijk, C.; Gerards, S.; De Boer, W. Volatiles Produced by the Mycophagous Soil Bacterium Collimonas. FEMS Microbiol. Ecol. 2014, 87, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P. A Method for Screening of Volatile Antimicrobial Compounds. Bull. Environ. Contam. Toxicol. 2011, 86, 145–148. [Google Scholar] [CrossRef]
- Brożyna, M.; Żywicka, A.; Fijałkowski, K.; Gorczyca, D.; Oleksy-Wawrzyniak, M.; Dydak, K.; Migdał, P.; Dudek, B.; Bartoszewicz, M.; Junka, A. The Novel Quantitative Assay for Measuring the Antibiofilm Activity of Volatile Compounds (AntiBioVol). Appl. Sci. 2020, 10, 7343. [Google Scholar] [CrossRef]
- Bonev, B.; Hooper, J.; Parisot, J. Principles of Assessing Bacterial Susceptibility to Antibiotics Using the Agar Diffusion Method. J. Antimicrob. Chemother. 2008, 61, 1295–1301. [Google Scholar] [CrossRef] [Green Version]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in Vitro Evaluating Antimicrobial Activity: A Review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Létoffé, S.; Audrain, B.; Bernier, S.P.; Delepierre, M.; Ghigo, J.-M. Aerial Exposure to the Bacterial Volatile Compound Trimethylamine Modifies Antibiotic Resistance of Physically Separated Bacteria by Raising Culture Medium PH. mBio 2014, 5, e00944-13. [Google Scholar] [CrossRef] [Green Version]
- Traxler, S.; Bischoff, A.-C.; Saß, R.; Trefz, P.; Gierschner, P.; Brock, B.; Schwaiger, T.; Karte, C.; Blohm, U.; Schröder, C.; et al. VOC Breath Profile in Spontaneously Breathing Awake Swine during Influenza A Infection. Sci. Rep. 2018, 8, 14857. [Google Scholar] [CrossRef]
- Lawal, O.; Knobel, H.; Weda, H.; Nijsen, T.M.E.; Goodacre, R.; Fowler, S.J. TD/GC-MS Analysis of Volatile Markers Emitted from Mono- and Co-Cultures of Enterobacter Cloacae and Pseudomonas Aeruginosa in Artificial Sputum. Metabolomics 2018, 14, 66. [Google Scholar] [CrossRef] [Green Version]
- Lawal, O.; Knobel, H.; Weda, H.; Bos, L.D.; Nijsen, T.M.E.; Goodacre, R.; Fowler, S.J. Volatile Organic Compound Signature from Co-Culture of Lung Epithelial Cell Line with Pseudomonas aeruginosa. Analyst 2018, 143, 3148–3155. [Google Scholar] [CrossRef] [PubMed]
- Neerincx, A.H.; Geurts, B.P.; van Loon, J.; Tiemes, V.; Jansen, J.J.; Harren, F.J.M.; Kluijtmans, L.A.J.; Merkus, P.J.F.M.; Cristescu, S.M.; Buydens, L.M.C.; et al. Detection of Staphylococcus Aureus in Cystic Fibrosis Patients Using Breath VOC Profiles. J. Breath Res. 2016, 10, 046014. [Google Scholar] [CrossRef] [PubMed]
- Cellini, A.; Buriani, G.; Rocchi, L.; Rondelli, E.; Savioli, S.; Rodriguez Estrada, M.T.; Cristescu, S.M.; Costa, G.; Spinelli, F. Biological Relevance of Volatile Organic Compounds Emitted during the Pathogenic Interactions between Apple Plants and Erwinia amylovora. Mol. Plant Pathol. 2018, 19, 158–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Volatile Producer | Volatile(s) | Bioactivity | Reference |
|---|---|---|---|
| Bacillus amyloliquefaciens CPA-8 | blend 1,3-pentadiene thiophene acetoine | f | [47] |
| Bacillus amyloliquefaciens DA12 | blend | f | [48] |
| Bacillus amyloliquefaciens FZB42 | blend 1,2-benzisothiazol-3(2H)-one benzaldehyde other | b | [49] |
| Bacillus amyloliquefaciens L3 other | blend 2-heptanone 2-ethyl-1-hexanol 2-nonanone other | f | [50] |
| Bacillus artrophaeus LSSC22 | blend 1,2-benzisothiazol-3(2H)-one other | b | [49] |
| Bacillus cereus CHP20 | blend | o | [44] |
| Bacillus megaterium KU143 | blend | f | [51] |
| Bacillus pumilus TM-R | blend | f | [52] |
| Bacillus siamensis LZ88 | blend | f | [53] |
| Bacillus (diverse spp.) | blend | f | [54] |
| Bacillus (diverse spp.) | blend 2-undecanone benzothiazole 1,3-butadiene N,N-dimethyldodecylamine other | f | [55] |
| Bacillus sp. BO53 | blend | b | [43] |
| Bacillus sp. D13 | blend | b | [56] |
| Bacillus sp. TM-I-3 | blend | f | [57] |
| Bacillus subtilis CHP14 | blend | o | [44] |
| Bacillus subtilis FA26 | blend benzaldehyde nonanal benzothiazole acetophenone | b | [58] |
| Bacillus subtilis M29 | blend 1-butanol acetic acid butyl ester 1-heptylene-4-alcohol 3-methyl-3-hexanol other | f | [59] |
| Bacillus velezensis BUZ-14 | blend diacetyl benzaldehyde isoamyl alcohol other | f | [41] |
| Bacillus velezensis G341 | blend | f | [60] |
| Bacillus velezensis I3 | blend | f | [41] |
| bacterial community | blend | f | [31] |
| Cronobacter muytjensii JZ38 | blend | o | [61] |
| Frigoribacterium endophyticum CHP33 | blend | o | [44] |
| Microbacterium testaceum KU313 | blend | f | [51] |
| Paenibacillus sp. AD87 | 2,5-bis(1-methylethyl)-pyrazine | b, f | [21] |
| Paenibacillus polymyxa Sb3-1 | blend | f | [42] |
| Proteus mirabilis 04 | blend | b | [62] |
| Pseudoalteromonas sp. GA327 | blend | b | [43] |
| Pseudomonas chlororaphis subsp. aurantiaca KNU17Pc1 | blend | f | [63] |
| Pseudomonas chlororaphis subsp. aureofaciens SPS-41 | blend 3-methyl-1-butanol phenylethyl alcohol 2-methyl-1-butanol other | f | [64] |
| Pseudomonas protegens AS15 | blend | f | [51] |
| Pseudomonas protegens CHA0 | dimethyl trisulfide 2-ethylhexanol ammonium hydroxide phenol acetophenone 1,3-diphenylpropane 3-phenylpropiophenone | f | [65] |
| Pseudomonas putida BP25 | blend 2-ethyl-5-methylpyrazine | f | [66] |
| Pseudomonas putida BP25 | 2,5-dimethyl pyrazine 2-methyl pyrazine 2-ethyl-5-methyl pyrazine 2-ethyl-3,6-dimethyl pyrazine dimethyl trisulfide | b, f, o | [67] |
| Pseudomonas stutzeri E25 | blend dimethyl disulfide | f | [68] |
| Sphingobacterium multivorum Bel3-4 | blend | f | [54] |
| Stenotrophomonas maltophilia CR71 | blend dimethyl disulfide | f | [68] |
| Stenotrophomonas maltophilia (TD1 and GH1-5) | blend | f | [54] |
| Streptomyces alboflavus TD-1 | blend anisole dimethyl trisulfide β-pinene benzenamine 1,5-cyclooctadiene | f | [69] |
| Streptomyces fimicarius BWL-H1 | phenylethyl alcohol ethyl phenylacetate methyl anthranilate α-copaene caryophyllene methyl salicylate 4-ethylphenol | o | [70] |
| Streptomyces lavendulae SPS-33 | blend 2-methyl-1-butanol 3-methyl-1-butanol pyridine phenylethyl alcohol other | f | [71] |
| Streptomyces sp. MBT11 | blend | b | [39] |
| Streptomyces venezuelae (ATCC 15439) | blend | b | [39] |
| Streptomyces yanglinensis 3–10 | blend methyl 2-methylbutyrate 2-phenylethanol β-caryophyllene | f, o f f f | [72] |
| Xenorhabdus szentirmaii PAM 25 | blend | f | [73] |
| Co-Culture | Volatile(s) | Bioactivity | Reference |
|---|---|---|---|
| Burkholderia sp. AD24 Paenibacillus sp. AD87 | 2,5-bis(1-methylethyl)-pyrazine | b, f | [21] |
| Burkholderia sp. AD024 Paenibacillus sp. AD087 Dyella sp. AD056 Janthinobacterium sp. AD080 Pseudomonas sp. AD021 | 2,5-bis(1-methylethyl)-pyrazine other | na | [82] |
| Chryseobacterium sp. AD48 Tsukamurella sp. AD106 | blend dimethyl trisulfide other | b, f, o b | [75] |
| Janthinobacterium sp. AD80 Dyella sp. AD56 | blend dimethyl trisulfide other | f, o b | [75] |
| Paenibacillus polymyxa Sb3-1 Verticillium longisporum EVL43 | blend trans-2,2,4,5-tetramethyl-1,3-dioxolane 1-butanol other | f na na | [42] |
| Serratia plymuthica 4Rx13 Staphylococcus delphini | schleiferon A and B | na | [83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lammers, A.; Lalk, M.; Garbeva, P. Air Ambulance: Antimicrobial Power of Bacterial Volatiles. Antibiotics 2022, 11, 109. https://doi.org/10.3390/antibiotics11010109
Lammers A, Lalk M, Garbeva P. Air Ambulance: Antimicrobial Power of Bacterial Volatiles. Antibiotics. 2022; 11(1):109. https://doi.org/10.3390/antibiotics11010109
Chicago/Turabian StyleLammers, Alexander, Michael Lalk, and Paolina Garbeva. 2022. "Air Ambulance: Antimicrobial Power of Bacterial Volatiles" Antibiotics 11, no. 1: 109. https://doi.org/10.3390/antibiotics11010109
APA StyleLammers, A., Lalk, M., & Garbeva, P. (2022). Air Ambulance: Antimicrobial Power of Bacterial Volatiles. Antibiotics, 11(1), 109. https://doi.org/10.3390/antibiotics11010109