
The Efficacy of Moina micrura Enriched with Probiotic Bacillus pocheonensis in Enhancing Survival and Disease Resistance of Red Hybrid Tilapia (Oreochromis spp.) Larvae


Abstract
1. Introduction
2. Results
2.1. Antagonistic Activity of Bacillus pocheonensis S2 against Freshwater Pathogens
2.2. Survival of Moina micrura after Challenged against Freshwater Pathogens
2.3. Relative Percentage Survival (RPS) of Moina micrura after Challenged against Freshwater Pathogens
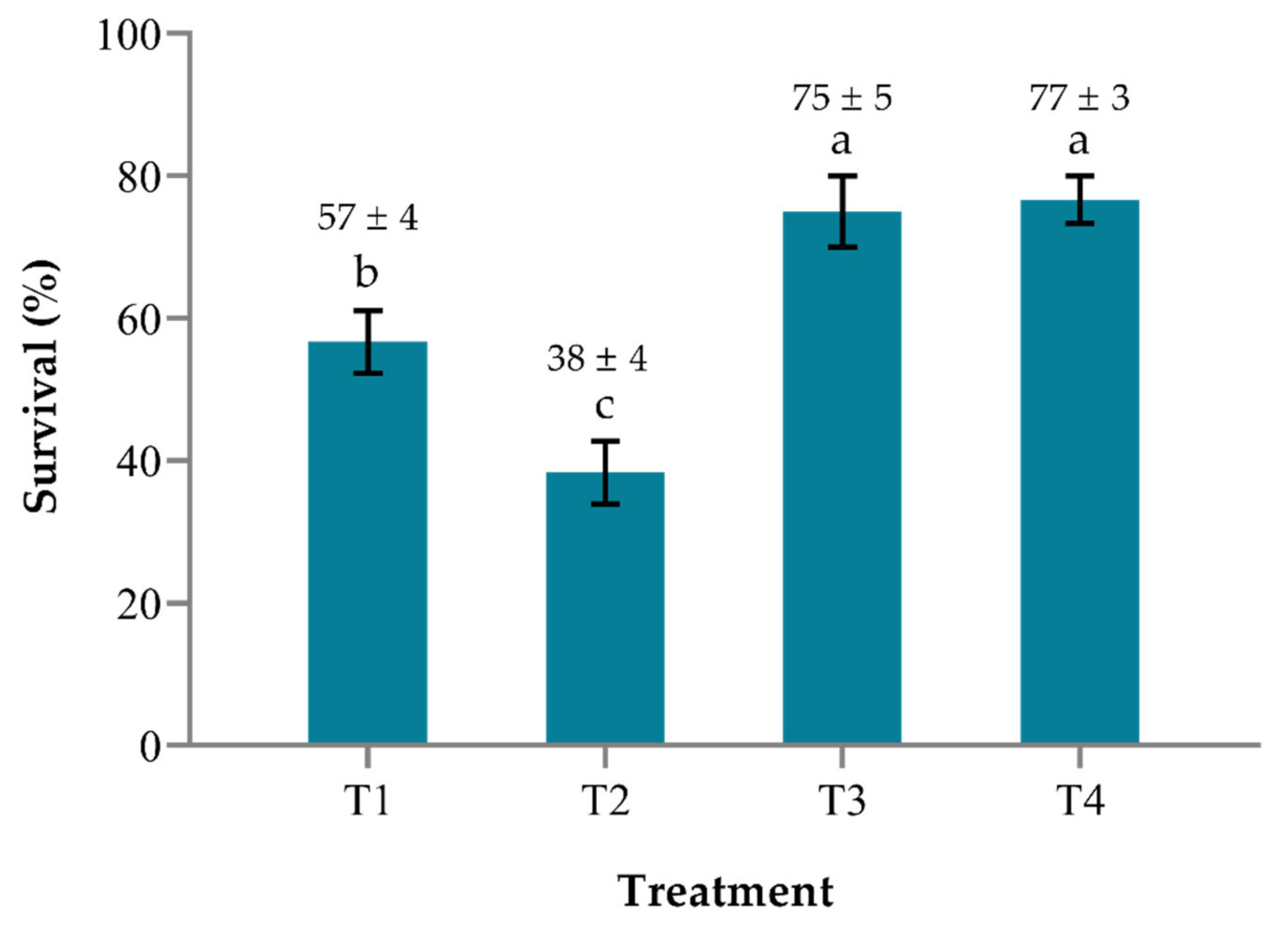
2.4. Survival and Relative Percentage Survival (RPS) of Red Hybrid Tilapia Larvae
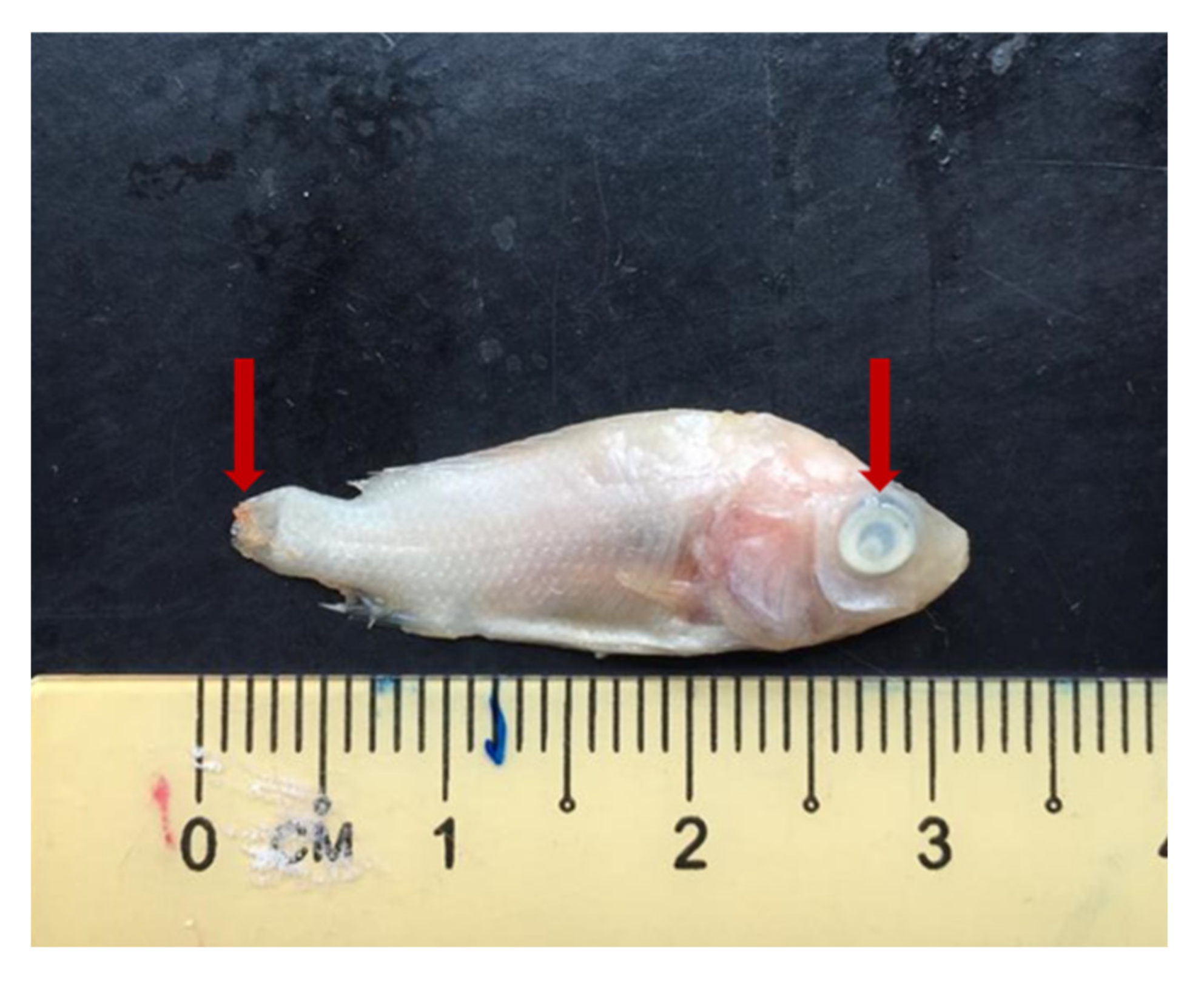
2.5. Streptococcus agalactiae Counts
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Cultures of Microalgae Chlorella vulgaris
4.3. Preliminary In Vitro Screening of Bacillus pocheonensis S2
4.4. Culture and Maintenance of Moina micrura
4.5. Preliminary In Vivo Bacterial Challenge Assay of Moina micrura
4.6. Enrichment of Moina micrura
4.7. Experimental Design
4.8. Streptococcus agalactiae Counts
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fletcher, R. 2020 Tilapia Production Figures Revealed. The Fish Site. 2020. Available online: https://thefishsite.com/articles/2020-tilapia-production-figures-revealed (accessed on 18 June 2021).
- Miao, W.; Wang, W. Trends of aquaculture production and trade: Carp, tilapia, and shrimp. Asian Fish. Sci. 2020, 33, 1–10. [Google Scholar] [CrossRef]
- Fitzssimmons, K. The Global Status of Tilapia Production and Marketing under COVID-19 Conditions. The Fish Site. 2020. Available online: https://thefishsite.com/articles/the-global-status-of-tilapia-production-and-marketing-under-covid-19-conditions (accessed on 18 June 2021).
- Mohamad, S.N.; Noordin, W.N.M.; Ismail, N.F.; Hamzah, A. Red hybrid tilapia (Oreochromis spp.) broodstock development programme in Malaysia: Status, challenges and prospects for future development. Asian Fish. Sci. 2021, 34, 73–81. [Google Scholar] [CrossRef]
- DOF. Zon Industri Akuakultur. Available online: https://www.dof.gov.my/index.php/pages/view/78 (accessed on 20 January 2021).
- Dullah, H.; Malek, M.A.; Hanafiah, M.M. Life cycle assessment of Nile tilapia (Oreochromis niloticus) farming in Kenyir Lake, Terengganu. Sustainability 2020, 12, 2268. [Google Scholar] [CrossRef]
- Assefa, A.; Abunna, F. Maintenance of fish health in aquaculture: Review of epidemiological approaches for prevention and control of infectious disease of fish. Vet. Med. Int. 2018, 2018, 5432497. [Google Scholar] [CrossRef]
- Cruz, P.M.; Ibáñez, A.L.; Hermosillo, O.A.M.; Saad, H.C.R. Use of Probiotics in Aquaculture. ISRN Microbiol. 2012, 2012, 916845. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2018—Meeting THE Sustainable Development Goals; Food and Agriculture Organization: Rome, Italy, 2018. [Google Scholar]
- Najiah, M.; Aqilah, N.I.; Lee, K.L.; Khairulbariyyah, Z.; Mithun, S.; Jalal, K.C.A.; Shaharom-Harrison, F.; Nadirah, M. Massive mortality associated with Streptococcus agalactiae infection in cage-cultured red hybrid tilapia Oreochromis niloticus in Como river, Kenyir Lake, Malaysia. J. Biol. Sci. 2012, 12, 438–442. [Google Scholar] [CrossRef][Green Version]
- Siti-Zahrah, A.; Padilah, B.; Azila, A.; Rimatulhana, R.; Shahidan, H. Multiple streptococcal species infection in cage-cultured red tilapia but showing similar clinical signs. In Diseases in Asian Aquaculture VI; Bondad-Reantaso, M.G., Mohan, C.V., Crumlish, M., Subasinghe, R.P., Eds.; Asian Fisheries Society: Manila, Philippines, 2008; pp. 313–320. [Google Scholar]
- Basri, L.; Nor, R.M.; Salleh, A.; Yasin, I.S.M.; Saad, M.Z.; Rahaman, N.Y.A.; Barkham, T.; Amal, M.N.A. Co-infections of tilapia lake virus, Aeromonas hydrophila, and Streptococcus agalactiae in farmed red hybrid tilapia. Animals 2020, 10, 2141. [Google Scholar] [CrossRef] [PubMed]
- Musa, N.; Wei, L.S.; Musa, N.; Hamdan, R.H.; Leong, L.K.; Wee, W.; Amal, M.N.; Kutty, B.M.; Abdullah, S.Z. Streptococcosis in red hybrid tilapia (Oreochromis niloticus) commercial farms in Malaysia. Aquac. Res. 2009, 40, 630–632. [Google Scholar] [CrossRef]
- Amal, M.N.A.; Zamri-Saad, M.; Iftikhar, A.R.; Siti-Zahrah, A.; Aziel, S.; Fahmi, S. An outbreak of Streptococcus agalactiae infection in cage-cultured golden pompano, Trachinotus blochii (Lacépède), in Malaysia. J. Fish Dis. 2012, 35, 849–852. [Google Scholar] [CrossRef] [PubMed]
- Aisyhah, M.A.; Amal, M.N.A.; Zamri-Saad, M.; Siti-Zahrah, A.; Shaqinah, N.N. Streptococcus agalactiae isolates from cultured fishes in Malaysia manifesting low resistance pattern towards selected antibiotics. J. Fish Dis. 2015, 38, 1093–1098. [Google Scholar] [CrossRef]
- Aly, S.M.; Mohamed, M.F.; John, G. Effect of probiotics on the survival, growth and challenge infection in tilapia nilotica (Oreochromis niloticus). Aquac. Res. 2008, 39, 647–656. [Google Scholar] [CrossRef]
- Nimrat, S.; Suksawat, S.; Boonthai, T.; Vuthiphandchai, V. Potential Bacillus probiotics enhance bacterial numbers, water quality and growth during early development of white shrimp (Litopenaeus vannamei). Vet. Microbiol. 2012, 159, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Zokaeifar, H.; Babaei, N.; Saad, C.R.; Kamarudin, M.S.; Sijam, K.; Balcazar, J.L. Administration of Bacillus subtilis strains in the rearing water enhances the water quality, growth performance, immune response, and resistance against Vibrio harveyi infection in juvenile white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2014, 36, 68–74. [Google Scholar] [CrossRef] [PubMed]
- FAO. Probiotics in Animal Nutrition—Production, Impact and Regulation; Food and Agriculture Organization: Rome, Italy, 2016. [Google Scholar]
- Jahangiri, L.; Esteban, M.Á. Administration of probiotics in the water in finfish aquaculture systems: A review. Fishes 2018, 3, 33. [Google Scholar] [CrossRef]
- Sun, Y.Z.; Yang, H.L.; Huang, K.P.; Ye, J.D.; Zhang, C.X. Application of autochthonous Bacillus bioencapsulated in copepod to grouper Epinephelus coioides larvae. Aquaculture 2013, 392–395, 44–50. [Google Scholar] [CrossRef]
- Jamali, H.; Imani, A.; Abdollahi, D.; Roozbehfar, R.; Isari, A. Use of probiotic Bacillus spp. in rotifer (Brachionus plicatilis) and Artemia (Artemia urmiana) enrichment: Effects on growth and survival of Pacific white shrimp, Litopenaeus vannamei larvae. Probiotics Antimicrob. Proteins 2015, 7, 118–125. [Google Scholar] [CrossRef]
- Hai, N.V. The use of probiotics in aquaculture. J. Appl. Microbiol. 2015, 119, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Samat, N.A.; Yusoff, F.M.; Rasdi, N.W.; Karim, M. Enhancement of live food nutritional status with essential nutrients for improving aquatic animal health: A Review. Animals 2020, 10, 2457. [Google Scholar] [CrossRef]
- Gatesoupe, F.J. Updating the importance of lactic acid bacteria in fish farming: Natural occurrence and probiotic treatments. J. Mol. Microbiol. Biotechnol. 2008, 14, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Poynton, S.L.; Dashsel, P.; Lehmann, M.J.; Steinbergb, C.E. Culture of the cladoceran Moina macrocopa: Mortality associated with flagellate infection. Aquaculture 2013, 416–417, 374–379. [Google Scholar] [CrossRef]
- Rasdi, N.W.; Arshad, A.; Ikhwanuddin, M.; Hagiwara, A.; Yusoff, F.M.; Azani, N. A review on the improvement of cladocera (Moina) nutrition as live food for aquaculture: Using valuable plankton fisheries resources. J. Environ. Biol. 2020, 41, 1239–1248. [Google Scholar] [CrossRef]
- Gao, X.; Ke, C.; Zhang, M.; Li, X.; Wu, F.; Liu, Y. Effects of the probiotic Bacillus amyloliquefaciens on the growth, immunity, and disease resistance of Haliotis discus hannai. Fish Shellfish Immunol. 2019, 94, 617–627. [Google Scholar] [CrossRef]
- Adorian, T.J.; Jamali, H.; Farsani, H.G.; Darvishi, P.; Hasanpour, S.; Bagheri, T.; Roozbehfar, R. Effects of probiotic bacteria Bacillus on growth performance, digestive enzyme activity, and hematological parameters of Asian seabass, Lates calcarifer (Bloch). Probiotics Antimicrob. Proteins 2018, 11, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Tarnecki, A.M.; Wafapoor, M.; Phillips, R.N.; Rhody, N.R. Benefits of a Bacillus probiotic to larval fish survival and transport stress resistance. Sci. Rep. 2019, 9, 4892. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Ten, L.N.; Baek, S.-H.; Im, W.-T.; Larina, L.L.; Lee, J.-S.; Oh, H.-M.; Lee, S.-T. Bacillus pocheonensis sp. nov., a moderately halotolerant, aerobic bacterium isolated from soil of a ginseng field. Int. J. Syst. Evol. Microbiol. 2007, 57, 2532–2537. [Google Scholar] [CrossRef]
- Ahmad, V.; Iqbal, A.N.; Haseeb, M.; Khan, M.S. Antimicrobial potential of bacteriocin producing Lysinibacillus jx416856 against foodborne bacterial and fungal pathogens, isolated from fruits and vegetable waste. Anaerobe 2014, 27, 87–95. [Google Scholar] [CrossRef]
- Ahmad, V.; Ahmad, K.; Baig, M.H.; AL-Shwaiman, H.A.; Al Khulaifi, M.M.; Elgorban, A.M.; Khan, M.S. Efficacy of a novel bacteriocin isolated from Lysinibacillus sp. against Bacillus pumilus. LWT-Food Sci. Technol. 2019, 102, 260–267. [Google Scholar] [CrossRef]
- Patel, A.K.; Deshattiwar, M.K.; Chaudhari, B.L.; Chincholkar, S.B. Production, purification and chemical characterization of the catecholate siderophore from potent probiotic strains of Bacillus spp. Bioresour. Technol. 2009, 100, 368–373. [Google Scholar] [CrossRef]
- Liñan-Vidriales, M.A.; Peña-Rodríguez, A.; Tovar-Ramírez, D.; Elizondo-González, R.; Barajas-Sandoval, D.R.; Ponce-Gracía, E.I.; Rodríguez-Jaramillo, C.; Balcázar, J.L.; Quiroz-Guzmán, E. Effect of rice bran fermented with Bacillus and Lysinibacillus species on dynamic microbial activity of Pacific white shrimp (Penaeus vannamei). Aquaculture 2021, 531, 735958. [Google Scholar] [CrossRef]
- Yilmaz, M.; Soran, H.; Beyatli, Y. Antimicrobial activities of some Bacillus spp. strains isolated from the soil. Microbiol. Res. 2006, 161, 127–131. [Google Scholar] [CrossRef]
- Gullian, M.; Thompson, F.; Rodriguez, J. Selection of probiotic bacteria and study of their immunostimulatory effect in Penaeus vannamei. Aquaculture 2004, 233, 1–14. [Google Scholar] [CrossRef]
- Chaurasia, B.; Pandey, A.; Palni, L.M.; Trivedi, P.; Kumar, B.; Colvin, N. Diffusible and volatile compounds produced by an antagonistic Bacillus subtilis strain cause structural deformations in pathogenic fungi in vitro. Microbiol. Res. 2005, 160, 75–81. [Google Scholar] [CrossRef]
- Giarma, E.; Amanetidou, E.; Toufexi, A.; Touraki, M. Defense systems in developing Artemia franciscana nauplii and their modulation by probiotic bacteria offer protection against a Vibrio anguillarum challenge. Fish Shellfish Immunol. 2017, 66, 163–172. [Google Scholar] [CrossRef]
- Motlagh, H.R.; Farhangi, M.; Rafiee, G.; Noori, F. Modulating gut microbiota and digestive enzyme activities of Artemia urniana by administration of different levels of Bacillus subtilis and Bacillus licheniformis. Aquac. Int. 2012, 20, 693–705. [Google Scholar] [CrossRef]
- Ahmadifard, N.; Aminlooi, V.R.; Tukmechi, A.; Agh, N. Evaluation of the impacts of long-term enriched Artemia with Bacillus subtilis on growth performance, reproduction, intestinal microflora, and resistance to Aeromonas hydrophila of ornamental fish Poecilia latipinna. Probiotics Antimicrob. Proteins 2019, 11, 957–965. [Google Scholar] [CrossRef]
- Touraki, M.; Karamanlidou, G.; Karavida, P.; Chrysi, K. Evaluation of the probiotics Bacillus subtilis and Lactobacillus plantarum bioencapsulated in Artemia nauplii against vibriosis in European sea bass larvae (Dicentrarchus labrax, L.). World J. Microbiol Biotechnol. 2012, 28, 2425–2433. [Google Scholar] [CrossRef] [PubMed]
- Ziaei-Nejad, S.; Rezaei, M.H.; Takami, G.A.; Lovett, D.L.; Mirvaghefi, A.R.; Shakouri, M. The effect of Bacillus spp. bacteria used as probiotics on digestive enzyme activity, survival and growth in the Indian white shrimp Fenneropenaeus indicus. Aquaculture 2006, 252, 516–524. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Khanongnuch, C.; Kanpiengjai, A.; Unban, K.; Van Kim, V.; Srichaiyo, S. Host-associated probiotics boosted mucosal and serum immunity, disease resistance and growth performance of Nile tilapia (Oreochromis niloticus). Aquaculture 2018, 491, 94–100. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Bradley, G.; Baker, R.T.; Davies, S.J. Probiotic applications for rainbow trout (Oncorhynchus mykiss Walbaum) II. Effects on growth performance, feed utilization, intestinal microbiota and related health criteria post antibiotic treatment. Aquac. Nutr. 2010, 16, 496–503. [Google Scholar] [CrossRef]
- Addo, S.; Carrias, A.A.; Williams, M.A.; Liles, M.; Terhune, J.S.; Davis, D.A. Effects of Bacillus subtilis strains on growth, immune parameters, and Streptococcus iniae susceptibility in Nile tilapia, Oreochromis niloticus. J. World Aquac. Soc. 2016, 48, 257–267. [Google Scholar] [CrossRef]
- Irianto, A.; Austin, B. Probiotics in aquaculture. J. Fish Dis. 2002, 25, 633–642. [Google Scholar] [CrossRef]
- Azrin, N.A.; Yuzine, E.; Ina-Salwany, M.Y.; Karim, M. The efficacy of potential probiont Bacillus amyloliquefaciens strain L11 in protecting Artemia nauplii and blue crab juveniles against Vibrio harveyi infection. J. Pure Appl. Microbiol. 2019, 13, 923–931. [Google Scholar] [CrossRef]
- Liu, K.F.; Chiu, C.H.; Shiu, Y.L.; Cheng, W.; Liu, C.H. Effects of the probiotic, Bacillus subtilis E20, on the survival, development, stress tolerance, and immune status of white shrimp, Litopenaeus vannamei larvae. Fish Shellfish Immunol. 2010, 28, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Singh, R. Probiotics in aquaculture: A promising emerging alternative approach. Symbiosis 2019, 77, 99–113. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, M.; Gao, F.; Lu, M.; Chen, G. Effects of dietary probiotic supplementation on the growth, gut health and disease resistance of juvenile Nile tilapia (Oreochromis niloticus). Anim. Nutr. 2020, 6, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Standen, B.T.; Rodiles, A.; Peggs, D.L.; Davies, S.J.; Santos, G.A.; Merrifield, D.L. 2015. Modulation of the intestinal microbiota and morphology of tilapia, Oreochromis niloticus, following the application of a multi-species probiotic. Appl. Microbiol. Biotechnol. 2015, 99, 8403–8417. [Google Scholar] [CrossRef]
- Pérez, T.; Balcázar, J.L.; Ruiz-Zarzuela, I.; Halaihel, N.; Vendrell, D.; de Blas, I.; Múzquiz, J.L. Host–microbiota interactions within the fish intestinal ecosystem. Mucosal Immunol. 2010, 3, 355–360. [Google Scholar] [CrossRef]
- Natasya-Ain, R. Evaluation of Potential Probiotic Bacteria for Microalgae Propagation and Artemia fransiscana (Kellog, 1906) Bioencapsulation. Master’s Thesis, Universiti Putra Malaysia, Serdang, Malaysia, November 2018. [Google Scholar]
- Samat, N.A.; Yusoff, F.M.; Chong, C.M.; Karim, M. Enrichment of freshwater zooplankton Moina micrura with probiotics isolated from microalgae. J. Environ. Biol. 2020, 41, 1215–1223. [Google Scholar] [CrossRef]
- Habib, M.A.B.; Yusoff, F.M.; Phang, S.M.; Mohamed, S. Growth and nutritional values of Moina micrura fed on Chlorella vulgaris grown in digested palm oil mill effluent. Asian Fish. Sci. 2003, 16, 107–120. [Google Scholar]
- Rasdi, N.W.; Qin, J.G. Impact of food type on growth, survival and reproduction of the cyclopoid copepod Cyclopina kasignete as a potential live food in aquaculture. Aquac. Int. 2018, 26, 1281–1295. [Google Scholar] [CrossRef]
- Jusoh, M.; Kasan, N.A.; Hashim, F.S.; Haris, N.; Zakaria, M.F.; Mohamed, N.N.; Rasdi, N.W.; Wahid, M.E.A.; Katayama, T.; Takahashi, K. Isolation of freshwater and marine indigenous microalgae species from Terengganu water bodies for potential uses as live feeds in aquaculture industry. Int. Aquat. Res. 2020, 12, 74–83. [Google Scholar] [CrossRef]
- Masduki, F.; Zakaria, T.; Min, C.C.; Karim, M. Evaluation of Enterococcus hirae LAB3 as potential probiont against Vibrio harveyi in Artemia nauplii and Asian seabass larvae (Lates calcarifer) cultures. J. Environ. Biol. 2020, 41, 1153–1159. [Google Scholar] [CrossRef]
- Azuraidi, O.M.; Yusoff, F.M.; Shamsudin, M.N.; Raha, R.A.; Alekseev, V.R.; Matias-Peralta, H.M. Effect of food density on male appearance and ephippia production in a tropical cladoceran, Moina micrura Kurz, 1874. Aquaculture 2013, 412–413, 131–135. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Pathogens | Concentration | Diameter of Inhibition Zone (mm) |
|---|---|---|
| SA | 18.2 ± 0.4 | |
| - | ||
| - | ||
| AH | 10.7 ± 0.3 | |
| 11.0 ± 0.0 | ||
| 8.8 ± 0.2 |
| Treatment (T) | Descriptions | Average Mortality | RPS |
|---|---|---|---|
| 2 | M. micrura + C. vulgaris + SA | a | - |
| 5 | M. micrura + C. vulgaris + B. pocheonensis at + SA | b | 38.33 |
| 6 | M. micrura + C. vulgaris + B. pocheonensis at + SA | b | 38.33 |
| 2 | M. micrura + C. vulgaris + AH | a | - |
| 5 | M. micrura + C. vulgaris + B. pocheonensis at + AH | b | 30.19 |
| 6 | M. micrura + C. vulgaris + B. pocheonensis at + AH | ab | 18.87 |
| Treatment (T) | Descriptions | Average Mortality | RPS |
|---|---|---|---|
| 2 | Tilapia + M. micrura + S. agalactiae | a | - |
| 4 | Tilapia + M. micrura + B. pocheonensis + S. agalactiae | b | 62.90 |
| Treatment (T) | Descriptions | |
|---|---|---|
| 2 | Tilapia + M. micrura + S. agalactiae | a |
| 4 | Tilapia + M. micrura + B. pocheonensis + S. agalactiae | a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samat, N.A.; Yusoff, F.M.; Rasdi, N.W.; Karim, M. The Efficacy of Moina micrura Enriched with Probiotic Bacillus pocheonensis in Enhancing Survival and Disease Resistance of Red Hybrid Tilapia (Oreochromis spp.) Larvae. Antibiotics 2021, 10, 989. https://doi.org/10.3390/antibiotics10080989
Samat NA, Yusoff FM, Rasdi NW, Karim M. The Efficacy of Moina micrura Enriched with Probiotic Bacillus pocheonensis in Enhancing Survival and Disease Resistance of Red Hybrid Tilapia (Oreochromis spp.) Larvae. Antibiotics. 2021; 10(8):989. https://doi.org/10.3390/antibiotics10080989
Chicago/Turabian StyleSamat, Nur Amalina, Fatimah Md Yusoff, Nadiah Wan Rasdi, and Murni Karim. 2021. "The Efficacy of Moina micrura Enriched with Probiotic Bacillus pocheonensis in Enhancing Survival and Disease Resistance of Red Hybrid Tilapia (Oreochromis spp.) Larvae" Antibiotics 10, no. 8: 989. https://doi.org/10.3390/antibiotics10080989
APA StyleSamat, N. A., Yusoff, F. M., Rasdi, N. W., & Karim, M. (2021). The Efficacy of Moina micrura Enriched with Probiotic Bacillus pocheonensis in Enhancing Survival and Disease Resistance of Red Hybrid Tilapia (Oreochromis spp.) Larvae. Antibiotics, 10(8), 989. https://doi.org/10.3390/antibiotics10080989