
Two Distinct Genotypes of KPC-2-Producing Klebsiella pneumoniae Isolates from South Korea



Abstract
:1. Introduction
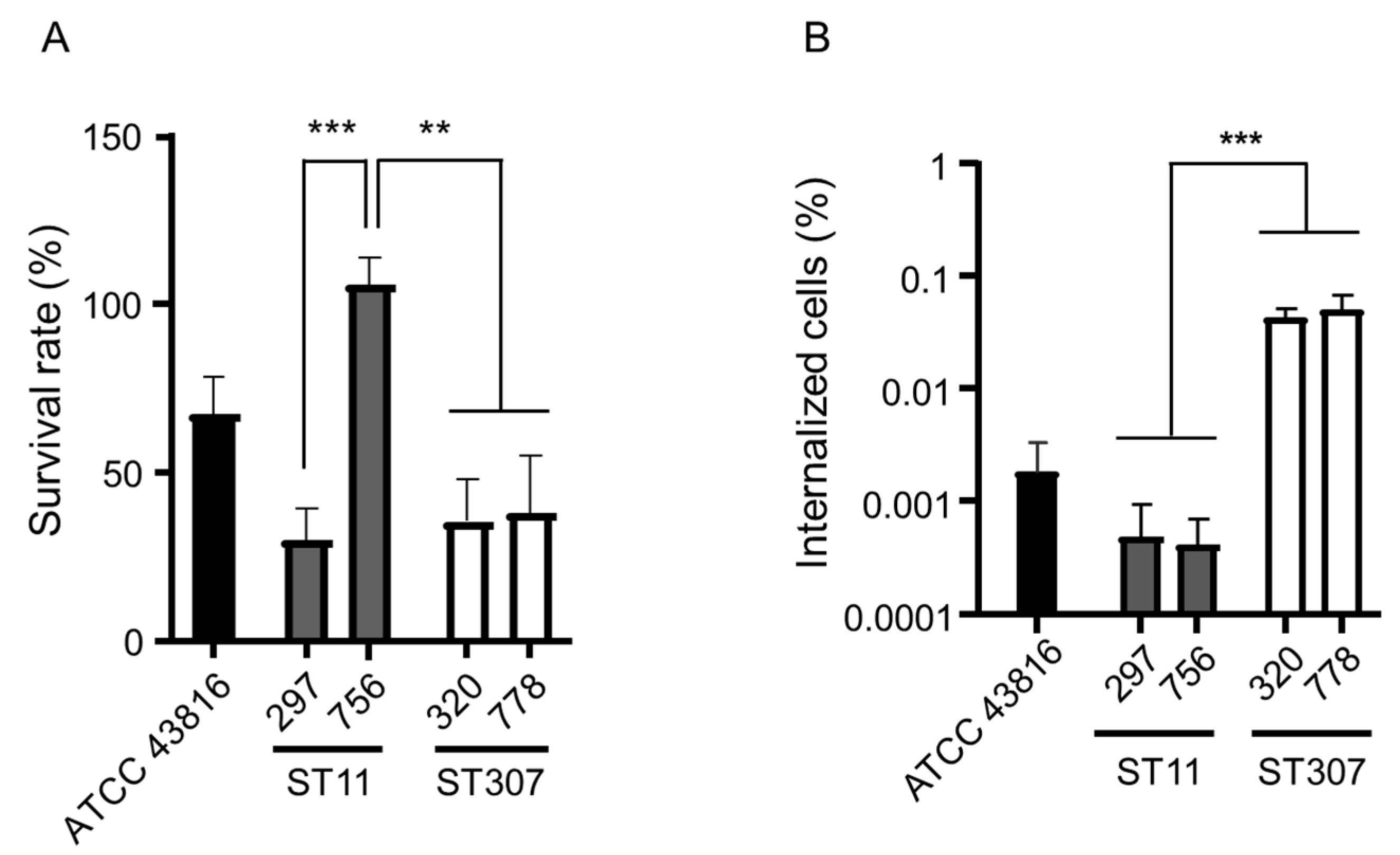
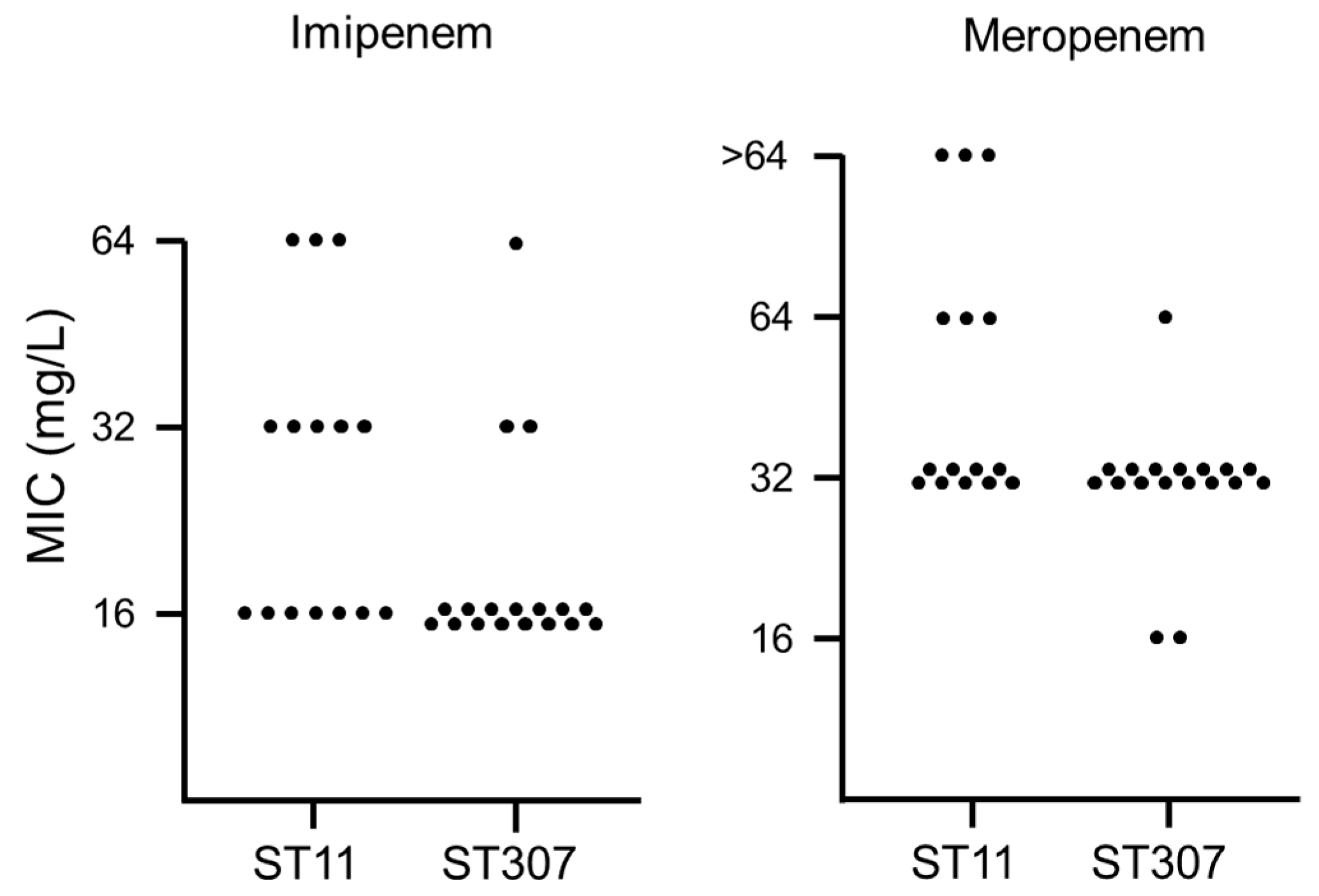
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antibiotic Susceptibility Testing
4.3. Genotyping and Plasmid Incompatibility Typing
4.4. String Test
4.5. Serum Resistance Assay
4.6. Macrophage Infection Assay
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rhee, J.Y.; Park, Y.K.; Shin, J.Y.; Choi, J.Y.; Lee, M.Y.; Peck, K.R.; Song, J.H.; Ko, K.S. KPC-producing extreme drug-resistant Klebsiella pneumoniae isolate from a patient with diabetes mellitus and chronic renal failure on hemodialysis in South Korea. Antimicrob. Agents Chemother. 2010, 54, 2278–2279. [Google Scholar] [CrossRef] [Green Version]
- Yoon, E.J.; Yang, J.W.; Kim, J.O.; Lee, H.; Lee, K.J.; Jeong, S.H. Carbapenemase-producing Enterobacteriaceae in South Korea: A report from the National Laboratory Surveillance System. Future Microbiol. 2018, 13, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Park, J.J.; Seo, Y.B.; Lee, J.; Eom, J.S.; Song, W.; Choi, Y.K.; Kim, S.R.; Son, H.J.; Cho, N.H. Positivity of Carbapenemase-producing Enterobacteriaceae in Patients Following Exposure within Long-term Care Facilities in Seoul, Korea. J. Korean Med. Sci. 2020, 35, e303. [Google Scholar] [CrossRef]
- Schwaber, M.J.; Lev, B.; Israeli, A.; Solter, E.; Smollan, G.; Rubinovitch, B.; Shalit, I.; Carmeli, Y. Containment of a Country-wide Outbreak of Carbapenem-Resistant Klebsiella pneumoniae in Israeli Hospitals via a Nationally Implemented Intervention. Clin. Infect. Dis. 2011, 52, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Mathema, B.; Chavda, K.D.; DeLeo, F.R.; Bonomo, R.A.; Kreiswirth, B.N. Carbapenemase-producing Klebsiella pneumoniae: Molecular and genetic decoding. Trends Microbiol. 2014, 22, 686–696. [Google Scholar] [CrossRef] [Green Version]
- Mathers, A.J.; Peirano, G.; Pitout, J.D.D. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef] [Green Version]
- Aslam, B.; Chaudhry, T.H.; Arshad, M.I.; Alvi, R.F.; Shahzad, N.; Yasmeen, N.; Idris, A.; Rasool, M.H.; Khurshid, M.; Ma, Z.; et al. The First blaKPC Harboring Klebsiella pneumoniae ST258 Strain Isolated in Pakistan. Microb. Drug Resist. 2020, 26, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Magi, G.; Tontarelli, F.; Caucci, S.; Sante, L.D.; Brenciani, A.; Morroni, G.; Giovanetti, E.; Menzo, S.; Mingoia, M. High prevalence of carbapenem-resistant Klebsiella pneumoniae ST307 recovered from fecal samples in an Italian hospital. Future Microbiol. 2021, 16, 703–711. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, Y.; Chen, G.; Lin, M.; Chen, Y.; He, R.; Galvão, K.N.; El-Gawad El-Sayed Ahmed, M.A.; Roberts, A.P.; Wu, Y.; et al. Molecular characterization of carbapenem-resistant and virulent plasmids in Klebsiella pneumoniae from patients with bloodstream infections in China. Emerg. Microbes. Infect. 2021, 10, 700–709. [Google Scholar] [CrossRef]
- Yoon, E.J.; Kim, J.O.; Kim, D.; Lee, H.; Yang, J.W.; Lee, K.J.; Jeong, S.H. Klebsiella pneumoniae Carbapenemase Producers in South Korea between 2013 and 2015. Front. Microbiol. 2018, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Park, B.Y.; Choi, M.H.; Yoon, E.J.; Lee, H.; Lee, K.J.; Park, Y.S.; Shin, J.H.; Un, Y.; Shin, K.S.; et al. Antimicrobial resistance and virulence factors of Klebsiella pneumoniae affecting 30 day mortality in patients with bloodstream infection. J. Antimicrob. Chemother. 2019, 74, 190–199. [Google Scholar] [CrossRef]
- García-Fernández, A.; Miriagou, V.; Papagiannitsis, C.C.; Giordano, A.; Venditti, M.; Mancini, C.; Carattoli, A. An ertapenem-resistant extended-spectrum-beta-lactamase-producing Klebsiella pneumoniae clone carries a novel OmpK36 porin variant. Antimicrob. Agents Chemother. 2010, 54, 4178–4184. [Google Scholar] [CrossRef] [Green Version]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the offense with a strong defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Y.; Wang, Y.; TWalsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Hunag, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Wi, Y.M.; Choi, J.Y.; Lee, J.Y.; Kang, C.I.; Chung, D.R.; Peck, K.R.; Song, J.H.; Ko, K.S. Emergence of colistin resistance in Pseudomonas aeruginosa ST235 clone in South Korea. Int. J. Antimicrob. Chemother. 2017, 49, 767–769. [Google Scholar] [CrossRef]
- Cortés, G.; Álvarez, D.; Saus, C.; Albertí, S. Role of lung epithelial cells in defense against Klebsiella pneumoniae pneumonia. Infect. Immun. 2002, 70, 1075–1080. [Google Scholar] [CrossRef] [Green Version]
- Ko, K.S. The contribution of capsule polysaccharide genes to virulence of Klebsiella pneumoniae. Virulence 2017, 8, 485–486. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Shin, J.; Shin, S.Y.; Ko, K.S. Characteristics of carbapenem-resistant Enterobacteriaceae isolates from Korea. Diagn. Microbiol. Infect. Dis. 2013, 76, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Findlay, J.; Hamouda, A.; Dancer, S.J.; Amyes, S.G.B. Rapid acquisition of decreased carbapenem susceptibility in a strain of Klebsiella pneumoniae arising during meropenem therapy. Clin. Microbiol. Infect. 2012, 18, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Pérez, F.J.; Hanson, N.D. Detection of plasmid-mediated AmpC β-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.K.; Lee, J.Y.; Ko, K.S. Colistin resistance in Enterobacter spp. isolates in Korea. J. Microbiol. 2018, 56, 435–440. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; 31st Informational Supplement. Document M100-S31; CLSI: Wayne, PA, USA, 2021. [Google Scholar]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.A.; Brisse, S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J. Clin. Microbiol. 2005, 43, 4178–4182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisse, S.; Passet, V.; Haugaard, A.B.; Babosan, A.; Kassis-Chikhani, N.; Struve, C.; Decré, D. wzi gene sequencing, a rapid method for determination of capsular type for Klebsiella strains. J. Clin. Microbiol. 2013, 51, 4073–4078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Shon, A.S.; Bajwa, R.P.S.; Russo, T.A. Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: A new and dangerous breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Podschun, R.; Sievers, D.; Fischer, A.; Serotypes, U.U. hemagglutinins, siderophore synthesis, and serum resistance of Klebsiella isolates causing human urinary tract infections. J. Infect. Dis. 1993, 168, 1415–1421. [Google Scholar] [CrossRef]
- Lee, H.; Baek, J.Y.; Kim, S.Y.; Jo, H.; Kang, K.; Ko, J.H.; Cho, S.Y.; Chung, D.R.; Peck, K.R.; Song, J.H.; et al. Comparison of virulence between matt and mucoid colonies of Klebsiella pneumoniae coproducing NDM-1 and OXA-232 isolated from a single patient. J. Microbiol. 2018, 56, 665–672. [Google Scholar] [CrossRef]
- Park, S.; Lee, H.; Shin, D.; Ko, K.S. Change of hypermucoviscosity in the development of tigecycline resistance in hypervirulent Klebsiella pneumoniae Sequence Type 23 strains. Microorganisms 2020, 8, 1562. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Genotype | Number of Resistant Isolates (%) a | CTX-M-15 | Plasmid | wzi Type | K-Serotype | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IMI | MER | COL | AMP | CPM | CIP | GEN | AZT | SXT | P/T | TIG | |||||
| ST307 (18) | 18 (100) | 18 (100) | 0 | 18 (100) | 18 (100) | 18 (100) | 18(100) | 18 (100) | 18(100) | 18 (100) | 6 (33.3) | 18 (100) | FIIK7 (13) FIIK21 (5) IncN6 (1) | 173 (18) | KL102 (18) |
| ST11 (15) | 15 (100) | 15 (100) | 3 (20.0) | 15 (100) | 15 (100) | 15 (100) | 2 (13.3) | 15 (100) | 6 (40.0) | 15 (100) | 0 | 3 (20.0) | FIIK1 (1) FIIK2 (1) FIIK5 (2) FIIK6 (1) FIIK7 (3) FIIK15 (2) FIIK21 (3) FIIK23 (1) FIIY6 (1) | 14 (8) 50 (7) | K14 (8) K51 (7) |
| ST789 (2) | 2 (100) | 2 (100) | 0 | 2 (100) | 2 (100) | 2 (100) | 0 | 2 (100) | 2 (100) | 2 (100) | 0 | 2 (100) | FIIK2 (2) | 18 (2) | K18 (2) |
| ST4681 (1) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | 1 (100) | FIIK7 (1) | 173 (1) | KL102 (1) |
| ST11slv (1) | 1 (100) | 1 (100) | 0 | 1 (100) | 1 (100) | 1 (100) | 0 | 1 (100) | 0 | 1 (100) | 0 | 0 | ND b | 2 (1) | K2 (1) |
| Total (37) | 37 (100) | 37 (100) | 4 (10.8) | 37 (100) | 37 (100) | 37 (100) | 21 (56.9) | 37 (100) | 27 (73.0) | 37 (100) | 8 (21.6) | 24 (64.9) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Cho, Y.Y.; Choi, J.Y.; Wi, Y.M.; Ko, K.S. Two Distinct Genotypes of KPC-2-Producing Klebsiella pneumoniae Isolates from South Korea. Antibiotics 2021, 10, 911. https://doi.org/10.3390/antibiotics10080911
Kim JH, Cho YY, Choi JY, Wi YM, Ko KS. Two Distinct Genotypes of KPC-2-Producing Klebsiella pneumoniae Isolates from South Korea. Antibiotics. 2021; 10(8):911. https://doi.org/10.3390/antibiotics10080911
Chicago/Turabian StyleKim, Jee Hong, Yun Young Cho, Ji Young Choi, Yu Mi Wi, and Kwan Soo Ko. 2021. "Two Distinct Genotypes of KPC-2-Producing Klebsiella pneumoniae Isolates from South Korea" Antibiotics 10, no. 8: 911. https://doi.org/10.3390/antibiotics10080911
APA StyleKim, J. H., Cho, Y. Y., Choi, J. Y., Wi, Y. M., & Ko, K. S. (2021). Two Distinct Genotypes of KPC-2-Producing Klebsiella pneumoniae Isolates from South Korea. Antibiotics, 10(8), 911. https://doi.org/10.3390/antibiotics10080911