
Activity of N-Chlorotaurine against Long-Term Biofilms of Bacteria and Yeasts




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial and Fungal Strains
2.2. N-Chlorotaurine
2.3. Devices for Growth of Biofilm
2.4. Growth of Biofilm of Bacteria in MBEC Inoculator and Incubation in NCT
2.5. Growth of Biofilm of Yeasts and Mixed Cultures of Yeast and Single Bacteria on Metal Discs and Incubation in NCT
2.6. Quantitative Cultures
2.7. Scanning Electron Microscopy
2.8. Statistics
3. Results
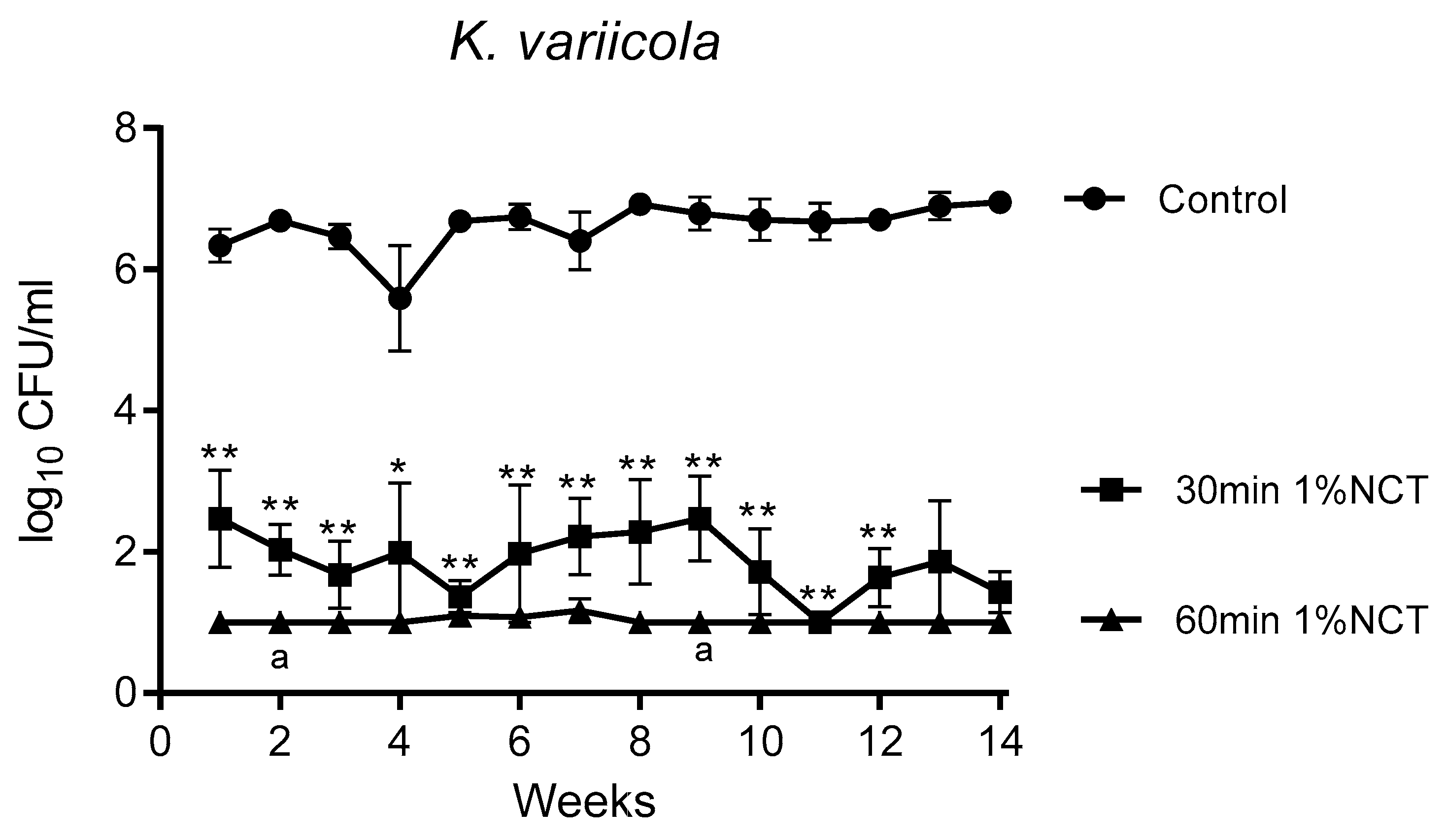
3.1. Bactericidal Effect of NCT in Long-Term Biofilms of Single Strains
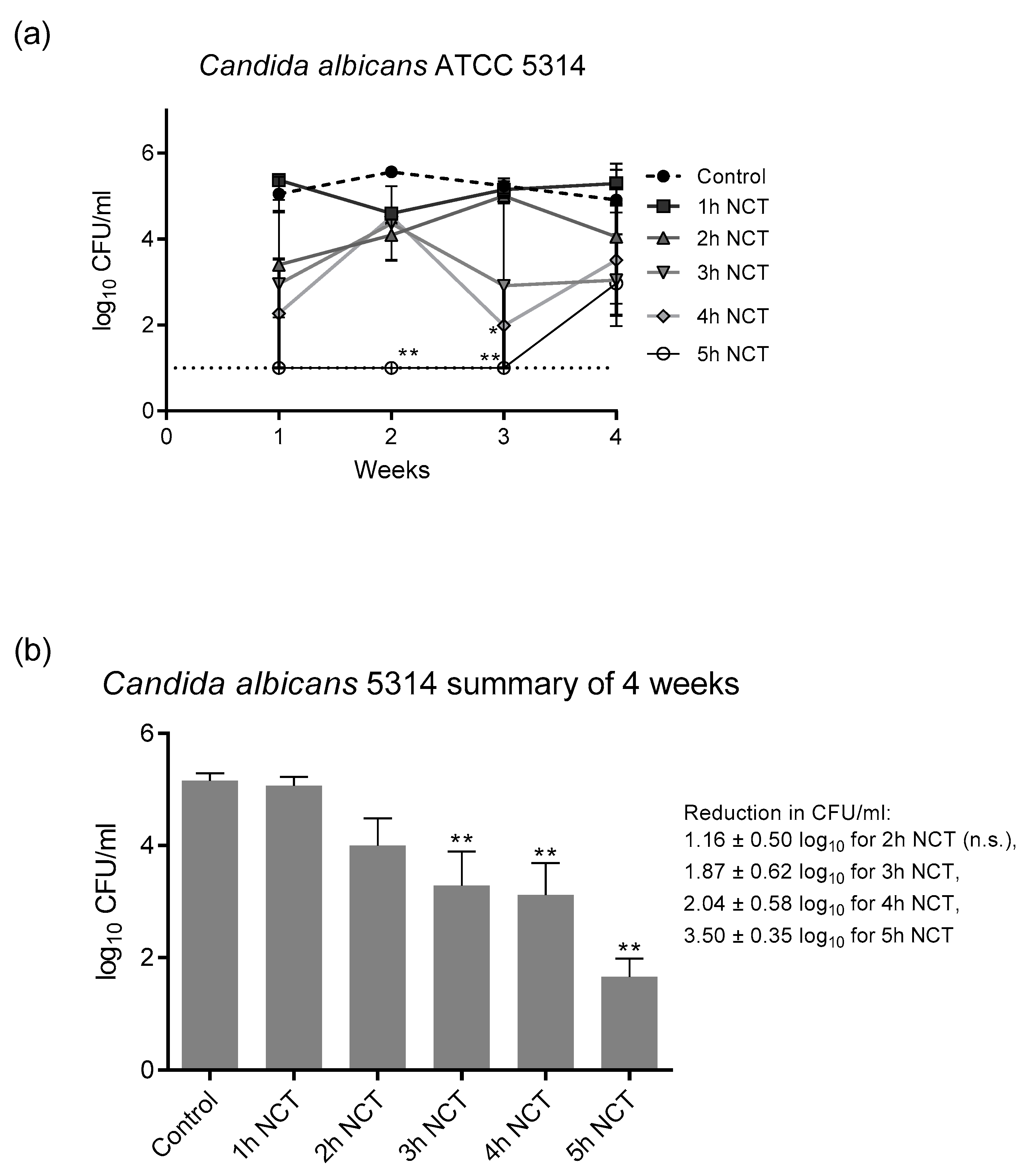
3.2. Fungicidal Effect of NCT in Long-Term Biofilms of Single C. albicans Strains
3.3. Bactericidal and Fungicidal Effect of NCT in Mixed Long-Term Biofilms
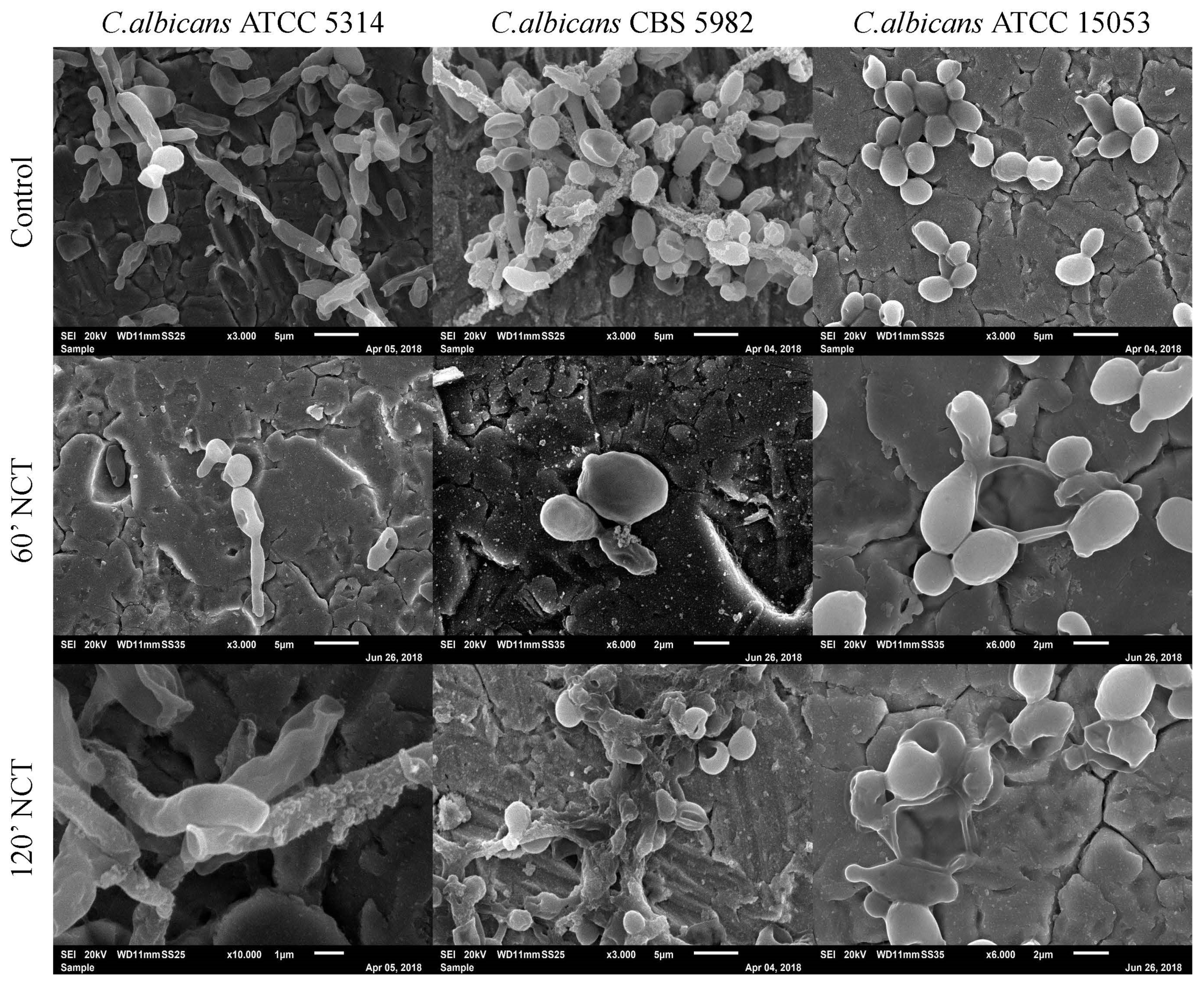
3.4. Scanning Electron Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schwarz, E.M.; McLaren, A.C.; Sculco, T.P.; Brause, B.; Bostrom, M.; Kates, S.L.; Parvizi, J.; Alt, V.; Arnold, W.V.; Carli, A.; et al. Adjuvant antibiotic-loaded bone cement: Concerns with current use and research to make it work. J. Orthop. Res. 2021, 39, 227–239. [Google Scholar] [CrossRef]
- Saeed, K.; McLaren, A.C.; Schwarz, E.M.; Antoci, V.; Arnold, W.V.; Chen, A.F.; Clauss, M.; Esteban, J.; Gant, V.; Hendershot, E.; et al. 2018 international consensus meeting on musculoskeletal infection: Summary from the biofilm workgroup and consensus on biofilm related musculoskeletal infections. J. Orthop. Res. 2019, 37, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.; Campoccia, D.; An, Y.; Baldassarri, L.; Pirini, V.; Donati, M.B.; Pegreffi, F.; Montanaro, L. Prevalence and Antibiotic Resistance of 15 Minor Staphylococcal Species Colonizing Orthopedic Implants. Int. J. Artif. Organs 2006, 29, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Campoccia, D.; Montanaro, L.; Von Eiff, C.; Pirini, V.; Ravaioli, S.; Becker, K.; Arciola, C.R. Cluster analysis of ribotyping profiles of Staphylococcus epidermidis isolates recovered from foreign body-associated orthopedic infections. J. Biomed. Mater. Res. Part A 2008, 88, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Zimmerli, W.; Trampuz, A.; Ochsner, P.E. Prosthetic-Joint Infections. N. Engl. J. Med. 2004, 351, 1645–1654. [Google Scholar] [CrossRef]
- Kresken, M.; Hafner, D.; The Study Group Bacterial Resistance of the Paul-Ehrlich-Society for Chemotherapy. Drug resistance among clinical isolates of frequently encountered bacterial species in central Europe during 1975–1995. Infection 1999, 27, S2–S8. [Google Scholar] [CrossRef]
- Giormezis, N.; Kolonitsiou, F.; Foka, A.; Drougka, E.; Liakopoulos, A.; Makri, A.; Papanastasiou, A.D.; Vogiatzi, A.; Dimitriou, G.; Marangos, M.; et al. Coagulase-negative staphylococcal bloodstream and prosthetic-device-associated infections: The role of biofilm formation and distribution of adhesin and toxin genes. J. Med. Microbiol. 2014, 63, 1500–1508. [Google Scholar] [CrossRef]
- Tande, A.J.; Patel, R. Prosthetic Joint Infection. Clin. Microbiol. Rev. 2014, 27, 302–345. [Google Scholar] [CrossRef]
- Weiss, S.J.; Klein, R.; Slivka, A.; Wei, M. Chlorination of Taurine by Human Neutrophils. J. Clin. Investig. 1982, 70, 598–607. [Google Scholar] [CrossRef]
- Zgliczyński, J.; Stelmaszyńska, T.; Domański, J.; Ostrowski, W. Chloramines as intermediates of oxidation reaction of amino acids by myeloperoxidase. Biochim. Biophys. Acta (BBA) Enzym. 1971, 235, 419–424. [Google Scholar] [CrossRef]
- Gottardi, W.; Nagl, M. N-chlorotaurine, a natural antiseptic with outstanding tolerability. J. Antimicrob. Chemother. 2010, 65, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Nagl, M.; Arnitz, R.; Lackner, M. N-Chlorotaurine, a Promising Future Candidate for Topical Therapy of Fungal Infections. Mycopathologia 2018, 183, 161–170. [Google Scholar] [CrossRef]
- Arnitz, R.; Stein, M.; Bauer, P.; Lanthaler, B.; Jamnig, H.; Scholl-Bürgi, S.; Stempfl-Al-Jazrawi, K.; Ulmer, H.; Baumgartner, B.; Embacher, S.; et al. Tolerability of inhaled N-chlorotaurine in humans: A double-blind randomized phase I clinical study. Ther. Adv. Respir. Dis. 2018, 12, 1–14. [Google Scholar] [CrossRef]
- Lackner, M.; Rössler, A.; Volland, A.; Stadtmüller, M.; Müllauer, B.; Banki, Z.; Ströhle, J.; Luttick, A.; Fenner, J.; Stoiber, H.; et al. N-chlorotaurine, a novel inhaled virucidal antiseptic is highly active against respiratory viruses including SARS-CoV-2 (COVID-19). Res. Sq. 2020. [Google Scholar] [CrossRef]
- Leiter, H.; Toepfer, S.; Messner, P.; Rabensteiner, M.; Gostner, J.M.; Lackner, M.; Hermann, M.; Nagl, M. Microbicidal activity of N-chlorotaurine can be enhanced in the presence of lung epithelial cells. J. Cyst. Fibros. 2020, 19, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Ammann, C.G.; Fille, M.; Hausdorfer, J.; Nogler, M.; Nagl, M.; Coraça-Huber, D.C. Influence of poly-N-acetylglucosamine in the extracellular matrix on N-chlorotaurine mediated killing of Staphylococcus epidermidis. New Microbiol. 2014, 37, 383–386. [Google Scholar]
- Coraca-Huber, D.C.; Ammann, C.; Fille, M.; Hausdorfer, J.; Nogler, M.; Nagl, M. Bactericidal activity of N-chlorotaurine against biofilm forming bacteria grown on metal discs. Antimicrob. Agents Chemother. 2014, 58, 2235–2239. [Google Scholar] [CrossRef][Green Version]
- Marcinkiewicz, J.; Strus, M.; Walczewska, M.; Machul, A.; Mikołajczyk, D. Influence of Taurine Haloamines (TauCl and TauBr) on the Development of Pseudomonas aeruginosa Biofilm: A Preliminary Study. Chem. Biol. Pteridines Folates 2013, 775, 269–283. [Google Scholar] [CrossRef]
- Nagl, M.; Nguyen, V.A.; Gottardi, W.; Ulmer, H.; Höpfl, R. Tolerability and efficacy of N-chlorotaurine compared to chloramine T for treatment of chronic leg ulcers with purulent coating. Br. J. Dermatol. 2003, 149, 590–597. [Google Scholar] [CrossRef]
- Neher, A.; Nagl, M.; Appenroth, E.; Gstöttner, M.; Wischatta, M.; Reisigl, F.; Schindler, M.; Ulmer, H.; Stephan, K. Acute Otitis Externa: Efficacy and Tolerability of N-Chlorotaurine, a Novel Endogenous Antiseptic Agent. Laryngoscope 2004, 114, 850–854. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, K.; Mayer, D.; Bruhn, G.; Noack, B.; Brecx, M.; Heumann, C.; Toutenburg, H.; Netuschil, L.; Nagl, M.; Gottardi, W.; et al. Effect of N-chlorotaurine mouth rinses on plaque regrowth and plaque vitality. Clin. Oral Investig. 2008, 13, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Stojicic, S.; Shen, Y.; Haapasalo, M. Effect of the Source of Biofilm Bacteria, Level of Biofilm Maturation, and Type of Disinfecting Agent on the Susceptibility of Biofilm Bacteria to Antibacterial Agents. J. Endod. 2013, 39, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Devivilla, S.; Lekshmi, M.; Kumar, S.H.; Valappil, R.K.; Roy, S.D.; Nayak, B.B. Effect of Sodium Hypochlorite on Biofilm-Forming Ability of Histamine-Producing Bacteria Isolated from Fish. J. Food Prot. 2019, 82, 1417–1422. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Medina, N.; Barrios-Camacho, H.; Duran-Bedolla, J.; Garza-Ramos, U. Klebsiella variicola: An emerging pathogen in humans. Emerg. Microbes Infect. 2019, 8, 973–988. [Google Scholar] [CrossRef]
- Gottardi, W.; Nagl, M. Chemical properties of N-chlorotaurine sodium, a key compound in the human defence system. Arch. Pharm. Pharm. Med. Chem. 2002, 335, 411–421. [Google Scholar] [CrossRef]
- Gottardi, W.; Nagl, M. Less is more, transferring a principle from art to science. Curr. Trends Microbiol. 2019, 13, 47–54. [Google Scholar]
- Kyriakopoulos, A.M.; Grapsa, E.; Marcinkiewicz, J.; Nagl, M. Swift Cure of a Chronic Wound Infected with Multiresistant Staphylococcus aureus in an Elderly Patient with Stage 5 Renal Disease. Int. J. Low. Extrem. Wounds 2019, 18, 192–196. [Google Scholar] [CrossRef]
- Kyriakopoulos, A.M.; Nagl, M.; Orth-Höller, D.; Marcinkiewicz, J.; Baliou, S.; Zoumbourlis, V. Successful treatment of a unique chronic multi-bacterial scalp infection with N-chlorotaurine, N-bromotaurine and bromamine T. Access Microbiol. 2020, 2, e000126. [Google Scholar] [CrossRef] [PubMed]
- Tré-Hardy, M.; Traore, H.; El Manssouri, N.; Vanderbist, F.; Vaneechoutte, M.; Devleeschouwer, M.J. Evaluation of long-term co-administration of tobramycin and clarithromycin in a mature biofilm model of cystic fibrosis clinical isolates of Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2009, 34, 370–374. [Google Scholar] [CrossRef][Green Version]
- Martini, C.; Hammerer-Lercher, A.; Zuck, M.; Jekle, A.; Debabov, D.; Anderson, M.; Nagl, M. Antimicrobial and anticoagulant activity of N-chlorotaurine (NCT), N,N-dichloro-2,2-dimethyltaurine (NVC-422) and N-monochloro-2,2-dimethyltaurine (NVC-612) in human blood. Antimicrob. Agents Chemother. 2012, 56, 1979–1984. [Google Scholar] [CrossRef][Green Version]
- Nagl, M.; Gruber, A.; Fuchs, A.; Lell, C.P.; Lemberger, E.-M.; Zepelin, M.B.-V.; Würzner, R. Impact of N-Chlorotaurine on Viability and Production of Secreted Aspartyl Proteinases of Candida spp. Antimicrob. Agents Chemother. 2002, 46, 1996–1999. [Google Scholar] [CrossRef] [PubMed]
- Gottardi, W.; Nagl, M. Chlorine covers on living bacteria: The initial step in antimicrobial action of active chlorine compounds. J. Antimicrob. Chemother. 2005, 55, 475–482. [Google Scholar] [CrossRef]
- Gruber, M.; Moser, I.; Nagl, M.; Lackner, M. Bactericidal and Fungicidal Activity of N-Chlorotaurine Is Enhanced in Cystic Fibrosis Sputum Medium. Antimicrob. Agents Chemother. 2017, 61, e02527-16. [Google Scholar] [CrossRef] [PubMed]
- Lackner, M.; Binder, U.; Reindl, M.; Gönül, B.; Fankhauser, H.; Mair, C.; Nagl, M. N-Chlorotaurine Exhibits Fungicidal Activity against Therapy-Refractory Scedosporium Species and Lomentospora prolificans. Antimicrob. Agents Chemother. 2015, 59, 6454–6462. [Google Scholar] [CrossRef]
- Nagl, M.; Gottardi, W. Enhancement of the bactericidal efficacy of N-chlorotaurine by inflammation samples and selected N-H compounds. Hyg. Med. 1996, 21, 597–605. [Google Scholar]
- Grisham, M.B.; Jefferson, M.M.; Melton, D.F.; Thomas, E.L. Chlorination of endogenous amines by isolated neutrophils. Ammonia-dependent bactericidal, cytotoxic, and cytolytic activities of the chloramines. J. Biol. Chem. 1984, 259, 10404–10413. [Google Scholar] [CrossRef]
- Orazi, G.; O’Toole, G.A. “It Takes a Village”: Mechanisms Underlying Antimicrobial Recalcitrance of Polymicrobial Biofilms. J. Bacteriol. 2019, 202, e00530-19. [Google Scholar] [CrossRef]
- Fourie, R.; Ells, R.; Swart, C.W.; Sebolai, O.M.; Albertyn, J.; Pohl, C.H. Candida albicans and Pseudomonas aeruginosa Interaction, with Focus on the Role of Eicosanoids. Front. Physiol. 2016, 7, 64. [Google Scholar] [CrossRef]
- Koo, H.; Andes, D.; Krysan, D.J. Candida–streptococcal interactions in biofilm-associated oral diseases. PLoS Pathog. 2018, 14, e1007342. [Google Scholar] [CrossRef]
- Xu, H.; Jenkinson, H.F.; Dongaribagtzoglou, A. Innocent until proven guilty: Mechanisms and roles of Streptococcus–Candida interactions in oral health and disease. Mol. Oral Microbiol. 2014, 29, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Bamunuarachchi, N.I.; Pham, D.T.N.; Tabassum, N.; Khan, M.S.A.; Kim, Y.-M. Mixed biofilms of pathogenic Candida-bacteria: Regulation mechanisms and treatment strategies. Crit. Rev. Microbiol. 2021, 1–29. [Google Scholar] [CrossRef]
- Eitzinger, C.; Ehrlenbach, S.; Lindner, H.; Kremser, L.; Gottardi, W.; Debabov, D.; Anderson, M.; Nagl, M.; Orth-Höller, D. N-Chlorotaurine, a Long-Lived Oxidant Produced by Human Leukocytes, Inactivates Shiga Toxin of Enterohemorrhagic Escherichia coli. PLoS ONE 2012, 7, e47105. [Google Scholar] [CrossRef]
- Jekle, A.; Yoon, J.; Zuck, M.; Najafi, R.; Wang, L.; Shiau, T.; Francavilla, C.; Rani, S.A.; Eitzinger, C.; Nagl, M.; et al. NVC-422 Inactivates Staphylococcus aureus Toxins. Antimicrob. Agents Chemother. 2012, 57, 924–929. [Google Scholar] [CrossRef]
- Reeves, E.P.; Nagl, M.; O’Keeffe, J.; Kelly, J.; Kavanagh, K. Effect of N-chlorotaurine on Aspergillus, with particular reference to destruction of secreted gliotoxin. J. Med Microbiol. 2006, 55, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, G.; Nagl, M.; Kavanagh, K. Exposure to N-chlorotaurine induces oxidative stress responses in Aspergillus fumigatus. J. Med. Microbiol. 2019, 68, 279–288. [Google Scholar] [CrossRef]
- Arnitz, R.; Sarg, B.; Ott, H.W.; Neher, A.; Lindner, H.; Nagl, M. Protein sites of attack of N-chlorotaurine in Escherichia coli. Proteomics 2006, 6, 865–869. [Google Scholar] [CrossRef] [PubMed]
- D’Lima, L.; Friedman, L.; Wang, L.; Xu, P.; Anderson, M.; Debabov, D. No Decrease in Susceptibility to NVC-422 in Multiple-Passage Studies with Methicillin-Resistant Staphylococcus aureus, S. aureus, Pseudomonas aeruginosa, and Escherichia coli. Antimicrob. Agents Chemother. 2012, 56, 2753–2755. [Google Scholar] [CrossRef] [PubMed][Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grimus, V.; Coraça-Huber, D.C.; Steixner, S.J.M.; Nagl, M. Activity of N-Chlorotaurine against Long-Term Biofilms of Bacteria and Yeasts. Antibiotics 2021, 10, 891. https://doi.org/10.3390/antibiotics10080891
Grimus V, Coraça-Huber DC, Steixner SJM, Nagl M. Activity of N-Chlorotaurine against Long-Term Biofilms of Bacteria and Yeasts. Antibiotics. 2021; 10(8):891. https://doi.org/10.3390/antibiotics10080891
Chicago/Turabian StyleGrimus, Victoria, Débora C. Coraça-Huber, Stephan J. M. Steixner, and Markus Nagl. 2021. "Activity of N-Chlorotaurine against Long-Term Biofilms of Bacteria and Yeasts" Antibiotics 10, no. 8: 891. https://doi.org/10.3390/antibiotics10080891
APA StyleGrimus, V., Coraça-Huber, D. C., Steixner, S. J. M., & Nagl, M. (2021). Activity of N-Chlorotaurine against Long-Term Biofilms of Bacteria and Yeasts. Antibiotics, 10(8), 891. https://doi.org/10.3390/antibiotics10080891