
Comparison of Antivirulence Activities of Black Ginseng against Methicillin-Resistant Staphylococcus aureus According to the Number of Repeated Steaming and Drying Cycles

 , , ,
, , , 
Abstract
1. Introduction
2. Results
2.1. Metabolite Profiling of Black Ginseng Products by UPLC-QTOF/MS
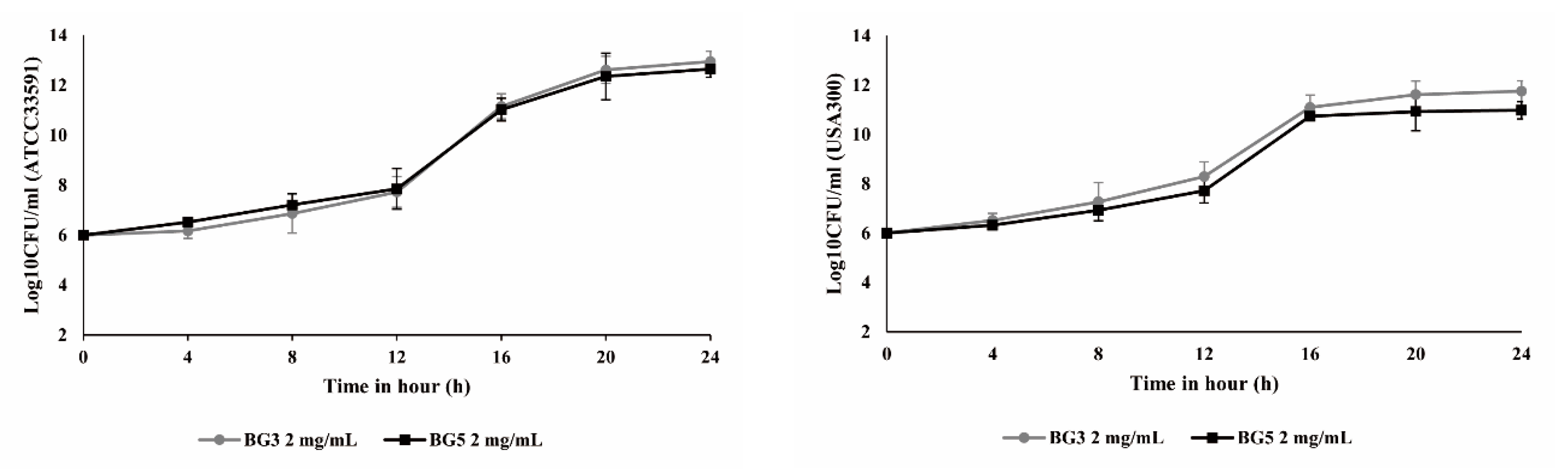
2.2. Growth Curve Analysis for Methicillin-Resistant Staphylococcus aureus
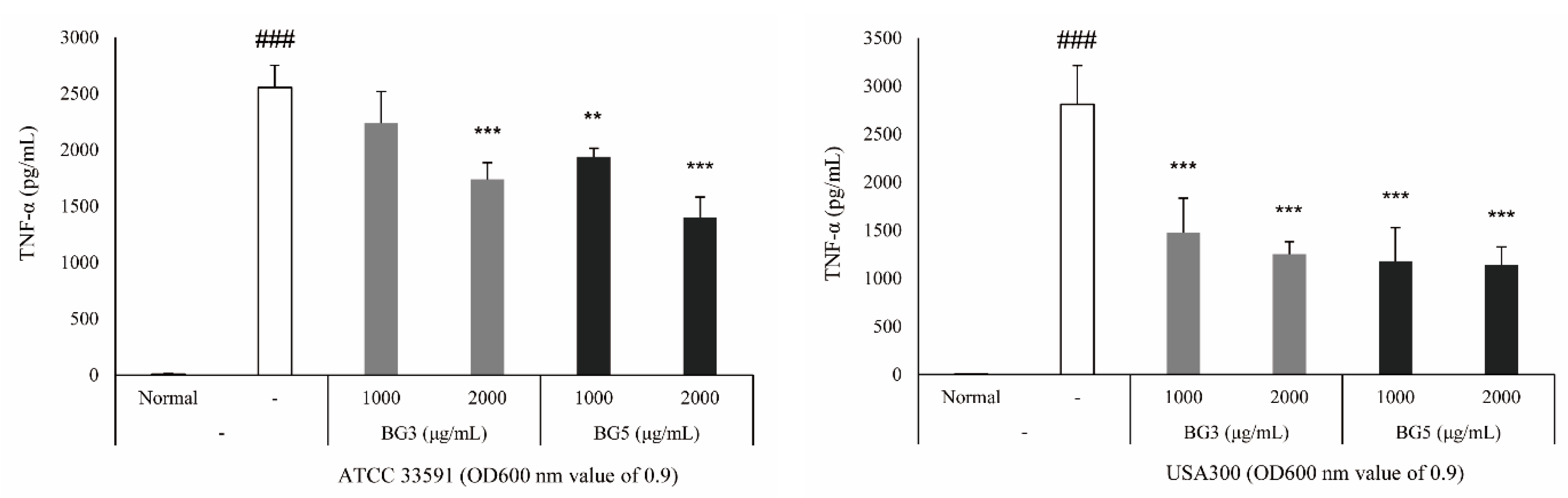
2.3. Inhibition of TNF-α Expression of BG Extracts in RAW 264.7 Cells
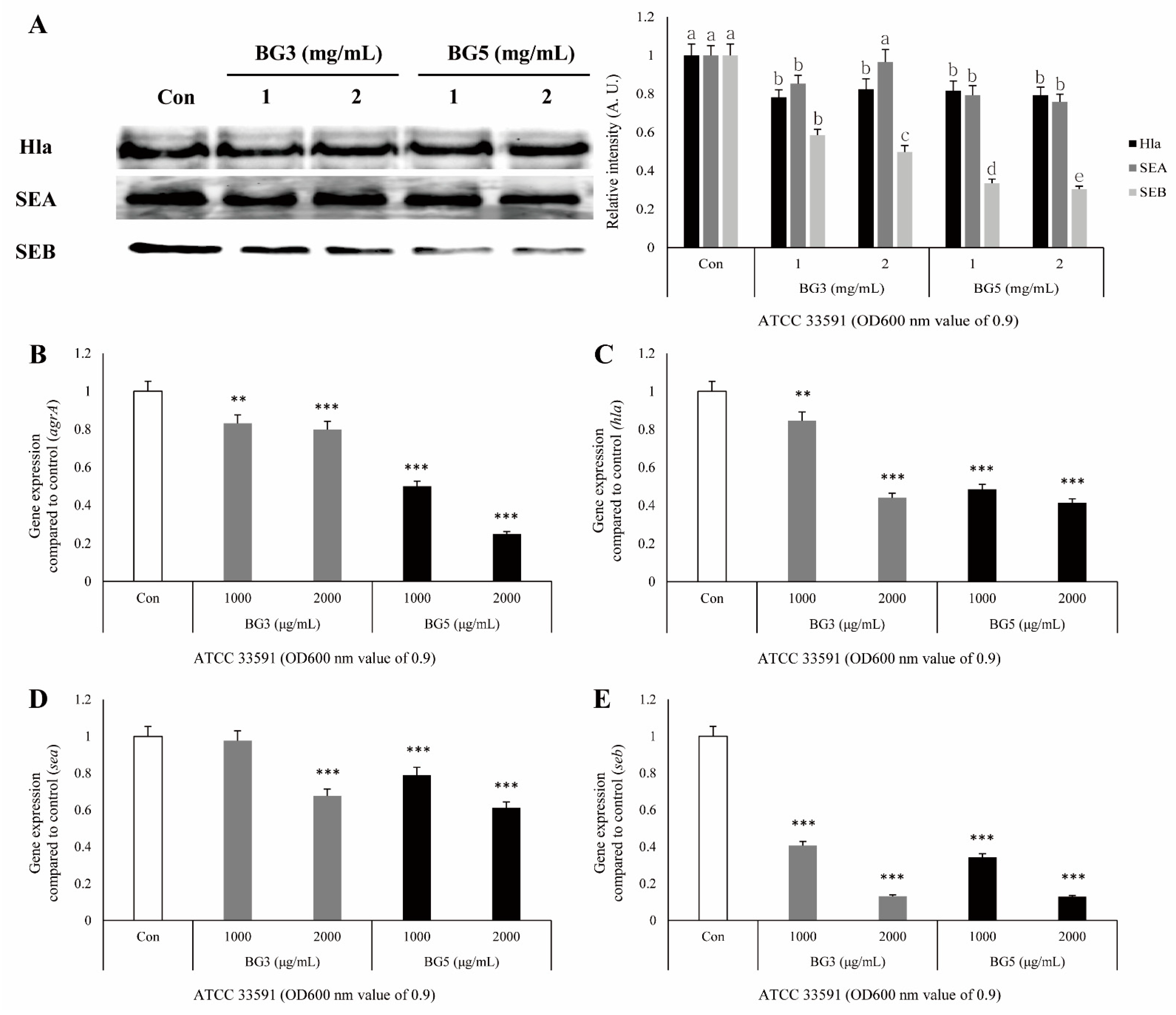
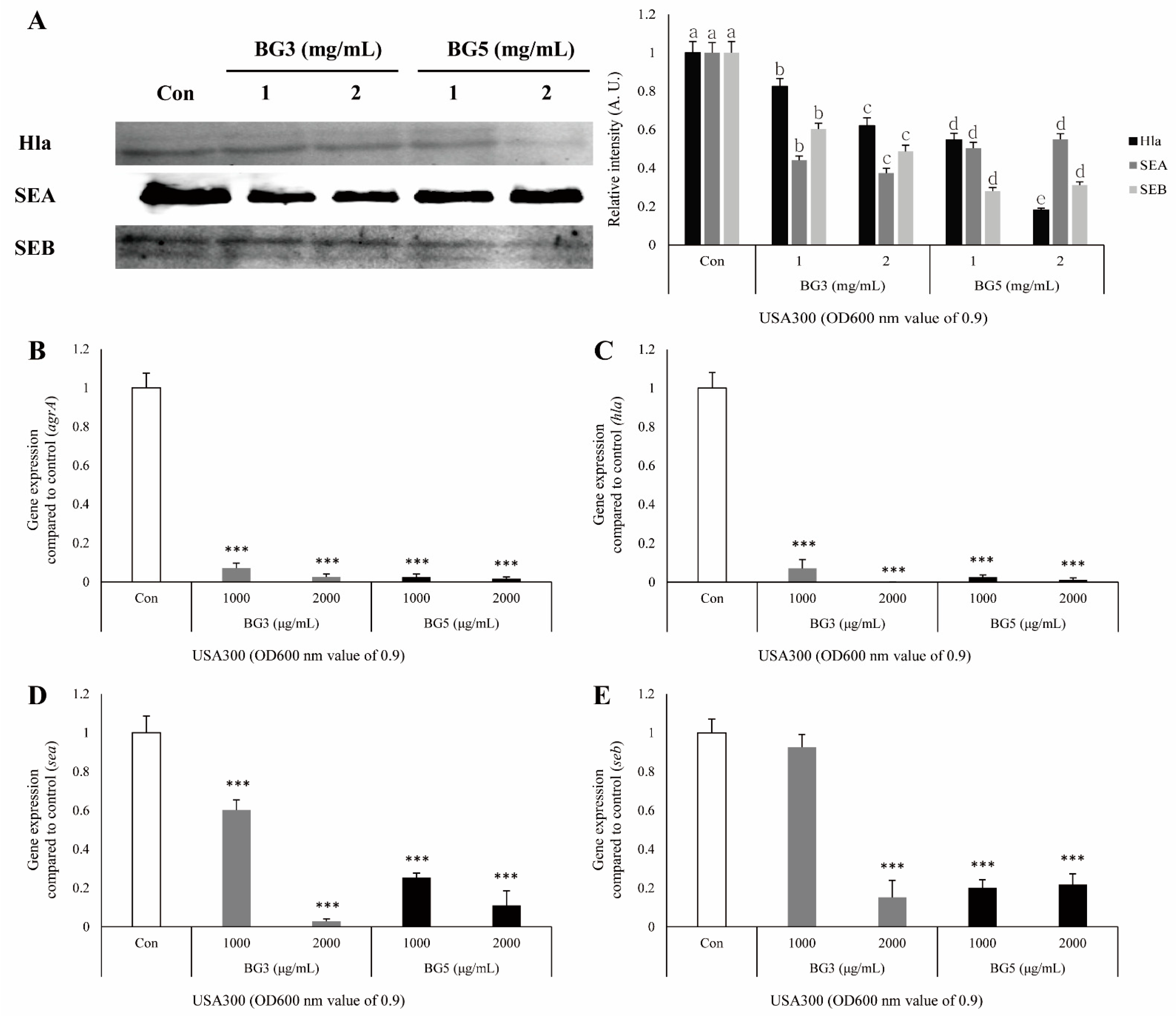
2.4. Inhibitory Effect of BG Extracts in Toxin Expression of S. aureus
2.5. Inhibitory Effect of BG Extracts on Expression of mRNA-Associated Virulence Factor of MRSA
3. Discussion
4. Materials and Methods
4.1. BG Extract Preparation
4.2. Bacterial Strains and Reagents
4.3. Measurement of Minimum Inhibitory Concentration, and Bacterial Growth Curve Assay
4.4. Western Blot Analysis
4.5. Tumor Necrosis Factor-α Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. RNA Extraction and Quantitative Reverse-Transcription–Polymerase Chain Reaction (qRT-PCR)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Uemura, E.; Kakinohana, S.; Higa, N.; Toma, C.; Nakasone, N. Comparative chracterization of Staphylococcus aureus isolates from throats and noses of healthy volunteers. Jpn. J. Infect. Dis. 2004, 57, 21–24. [Google Scholar]
- Sakr, A.; Brégeon, F.; Mège, J.-L.; Rolain, J.-M.; Blin, O. Staphylococcus aureus Nasal Colonization: An Update on Mechanisms, Epidemiology, Risk Factors, and Subsequent Infections. Front. Microbiol. 2018, 9, 2419. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Liu, Y.; Zhang, H.; Lei, L. The Pathogenicity and Transcriptome Analysis of Methicillin-Resistant Staphylococcus Aureus in Response to Water Extract of Galla Chinensis. Evid. Based Complement. Alternat. Med. 2019, 2019, 3276156. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Saidi, N.; Owlia, P.; Marashi, S.M.A.; Saderi, H. Inhibitory effect of probiotic yeast Saccharomyces cerevisiae on biofilm formation and expression of α-hemolysin and enterotoxin A genes of Staphylococcus aureus. Iran. J. Microbiol. 2019, 11, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Diep, B.A.; Le, V.T.M.; Visram, Z.C.; Rouha, H.; Stulik, L.; Dip, E.C.; Nagy, G.; Nagy, E. Improved Protection in a Rabbit Model of Community-Associated Methicillin-Resistant Staphylococcus aureus Necrotizing Pneumonia upon Neutralization of Leukocidins in Addition to Alpha-Hemolysin. Antimicrob. Agents Chemother. 2016, 60, 6333–6340. [Google Scholar] [CrossRef]
- Seilie, E.S.; Bubeck Wardenburg, J. Staphylococcus aureus pore-forming toxins: The interface of pathogen and host complexity. Semin. Cell Dev. Biol. 2017, 72, 101–116. [Google Scholar] [CrossRef]
- Vandenesch, F.; Lina, G.; Henry, T. Staphylococcus aureus hemolysins, bi-component leukocidins, and cytolytic peptides: A redundant arsenal of membrane-damaging virulence factors? Front. Cell. Inf. Microbiol. 2012, 2, 12. [Google Scholar] [CrossRef]
- Qiu, J.; Wang, D.; Xiang, H.; Feng, H.; Jiang, Y.; Xia, L.; Dong, J.; Lu, J.; Yu, L.; Deng, X. Subinhibitory concentrations of thymol reduce enterotoxins A and B and alpha-hemolysin production in Staphylococcus aureus isolates. PLoS ONE 2010, 5, e9736. [Google Scholar] [CrossRef]
- Argudín, M.Á.; Mendoza, M.C.; Rodicio, M.R. Food poisoning and Staphylococcus aureus enterotoxins. Toxins 2010, 2, 1751–1773. [Google Scholar] [CrossRef]
- Rusnak, J.M.; Kortepeter, M.; Ulrich, R.; Poli, M.; Boudreau, E. Laboratory Exposures to Staphylococcal Enterotoxin B. Emerg. Infect. Dis. 2004, 10, 1544–1549. [Google Scholar] [CrossRef]
- Pinchuk, I.V.; Beswick, E.J.; Reyes, V.E. Staphylococcal Enterotoxins. Toxins 2010, 2, 2177–2197. [Google Scholar] [CrossRef] [PubMed]
- Cosgriff, C.J.; White, C.R.; Teoh, W.P.; Grayczyk, J.P.; Alonzo, F. Control of Staphylococcus aureus Quorum Sensing by a Membrane-Embedded Peptidase. Infect. Immun. 2019, 87, e00019-19. [Google Scholar] [CrossRef]
- Kong, R.; Lee, Y.-S.; Kang, D.-H.; Wang, S.; Li, Q.; Kwon, D.-Y.; Kang, O.-H. The antibacterial activity and toxin production control of bee venom in mouse MRSA pneumonia model. BMC Complement. Med. Ther. 2020, 20, 238. [Google Scholar] [CrossRef]
- Zuo, G.-Y.; Zhang, X.-J.; Yang, C.-X.; Han, J.; Wang, G.-C.; Bian, Z.-Q. Evaluation of traditional Chinese medicinal plants for anti-MRSA activity with reference to the treatment record of infectious diseases. Molecules 2012, 17, 2955–2967. [Google Scholar] [CrossRef]
- Hemaiswarya, S.; Kruthiventi, A.K.; Doble, M. Synergism between Natural Products and Antibiotics against Infectious Diseases. Phytomedicine 2008, 15, 639–652. [Google Scholar] [CrossRef]
- Ha, Y.W.; Lim, S.S.; Ha, I.J.; Na, Y.-C.; Seo, J.-J.; Shin, H.; Son, S.H.; Kim, Y.S. Preparative isolation of four ginsenosides from Korean red ginseng (steam-treated Panax ginseng C. A. Meyer), by high-speed counter-current chromatography coupled with evaporative light scattering detection. J. Chromatogr. A 2007, 1151, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Kachur, K.; Suntres, Z.E. The antimicrobial properties of ginseng and ginseng extracts. Expert Rev. Anti Infect. Ther. 2016, 14, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Luan, X.; Dou, D.; Huang, L. Comparative Analysis of Ginsenosides and Oligosaccharides in White Ginseng (WG), Red Ginseng (RG) and Black Ginseng (BG). J. Chromatogr. Sci. 2019, 57, 403–410. [Google Scholar] [CrossRef]
- Kim, Y.-R.; Yang, C.-S. Protective roles of ginseng against bacterial infection. Microb. Cell 2018, 5, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Park, I.H.; Han, S.B.; Kim, J.M.; Piao, L.; Kwon, S.W.; Kim, N.Y.; Kang, T.L.; Park, M.K.; Park, J.H. Four new acetylated ginsenosides from processed ginseng (sun ginseng). Arch. Pharm. Res. 2002, 25, 837–841. [Google Scholar] [CrossRef]
- Nam, K.-Y.; Lee, N.-R.; Moon, B.-D.; Song, G.-Y.; Shin, H.-S.; Choi, J.-E. Changes of Ginsenosides and Color from Black Ginsengs Prepared by Steaming-Drying Cycles. Korean J. Med. Crop Sci. 2012, 20, 27–35. [Google Scholar] [CrossRef]
- Jin, Y.; Kim, Y.-J.; Jeon, J.-N.; Wang, C.; Min, J.-W.; Noh, H.-Y.; Yang, D.-C. Effect of white, red and black ginseng on physicochemical properties and ginsenosides. Plant Foods Hum. Nutr. 2015, 70, 141–145. [Google Scholar] [CrossRef]
- Ban, Y.-J. Optimization of the Manufacturing Process for Black Ginseng. J. Korean Soc. Appl. Biol. Chem. 2009, 53, 71–77. [Google Scholar] [CrossRef]
- Park, S.-J.; Park, M.; Sharma, A.; Kim, K.; Lee, H.-J. Black Ginseng and Ginsenoside Rb1 Promote Browning by Inducing UCP1 Expression in 3T3-L1 and Primary White Adipocytes. Nutrients 2019, 11, 2747. [Google Scholar] [CrossRef] [PubMed]
- Metwaly, A.M.; Lianlian, Z.; Luqi, H.; Deqiang, D. Black Ginseng and Its Saponins: Preparation, Phytochemistry and Pharmacological Effects. Molecules 2019, 24, 1856. [Google Scholar] [CrossRef] [PubMed]
- Youn, U.J.; Gu, B.-S.; Kim, K.H.; Ha, C.; Jung, I.C. Variation of main components according to the number of steaming and drying of Rehmanniae radix preparata. J. Pharmacopunct. 2018, 21, 112–119. [Google Scholar]
- Lee, D.Y.; Kim, M.-J.; Yoon, D.; Lee, Y.-S.; Kim, G.-S.; Yoo, Y.C. Ginseng Berry Prevents Alcohol-Induced Liver Damage by Improving the Anti-Inflammatory System Damage in Mice and Quality Control of Active Compounds. Int. J. Mol. Sci. 2019, 20, 3522. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Wang, R.; Khan, B.A.; Sturdevant, D.E.; Otto, M. Role of the accessory gene regulator agr in community-associated methicillin-resistant Staphylococcus aureus pathogenesis. Infect. Immun. 2011, 79, 1927–1935. [Google Scholar] [CrossRef]
- Choi, J.H.; Jang, A.Y.; Lin, S.; Lim, S.; Kim, D.; Park, K.; Han, S.-M.; Yeo, J.-H.; Seo, H.S. Melittin, a Honeybee venom-derived antimicrobial peptide, may target methicillin-resistant Staphylococcus aureus. Mol. Med. Rep. 2015, 12, 6483–6490. [Google Scholar] [CrossRef]
- Mun, S.-H.; Kong, R.; Seo, Y.-S.; Zhou, T.; Kang, O.-H.; Shin, D.-W.; Kwon, D.-Y. Subinhibitory concentrations of punicalagin reduces expression of virulence-related exoproteins by Staphylococcus aureus. FEMS Microbiol. Lett. 2016, 363, fnw253. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Wang, J.; Luo, H.; Du, X.; Li, H.; Luo, M.; Dong, J.; Chen, Z.; Deng, X. The effects of subinhibitory concentrations of costus oil on virulence factor production in Staphylococcus aureus. J. Appl. Microbiol. 2011, 110, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Pontzer, C.H.; Russell, J.K.; Johnson, H.M. Localization of an Immune Functional Site on Staphylococcal Enterotoxin A Using the Synthetic Peptide Approach. J. Immunol. 1989, 143, 280–284. [Google Scholar] [PubMed]
- Lindsay, J.A. Hospital-associated MRSA and antibiotic resistance-what have we learned from genomics? Int. J. Med. Microbiol. 2013, 303, 318–323. [Google Scholar] [CrossRef]
- Bukharie, H. A review of community-acquired methicillin-resistant Staphylococcus aureus for primary care physicians. J. Fam. Community Med. 2010, 17, 117–120. [Google Scholar] [CrossRef]
- Bhakdi, S.; Tranum-Jensen, J. Alpha-toxin of Staphylococcus aureus. Microbiol. Rev. 1991, 55, 733–751. [Google Scholar] [CrossRef]
- Bantel, H.; Sinha, B.; Domschke, W.; Peters, G.; Schulze-Osthoff, K.; Jänicke, R.U. alpha-Toxin is a mediator of Staphylococcus aureus-induced cell death and activates caspases via the intrinsic death pathway independently of death receptor signaling. J. Cell Biol. 2001, 155, 637–648. [Google Scholar] [CrossRef]
- Leng, B.-F.; Qiu, J.-Z.; Dai, X.-H.; Dong, J.; Wang, J.-F.; Luo, M.-J.; Li, H.-E.; Niu, X.-D.; Zhang, Y.; Ai, Y.-X.; et al. Allicin reduces the production of α-toxin by Staphylococcus aureus. Molecules 2011, 16, 7958–7968. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Zhao, X.; Li, W.; Meng, R.; Liu, Z.; Liu, M.; Guo, N.; Yu, L. Inhibitory effect of totarol on exotoxin proteins hemolysin and enterotoxins secreted by Staphylococcus aureus. World J. Microbiol. Biotechnol. 2015, 31, 1565–1573. [Google Scholar] [CrossRef]
- Qiu, J.; Zhang, X.; Luo, M.; Li, H.; Dong, J.; Wang, J.; Leng, B.; Wang, X.; Feng, H.; Ren, W.; et al. Subinhibitory concentrations of perilla oil affect the expression of secreted virulence factor genes in Staphylococcus aureus. PLoS ONE 2011, 6, e16160. [Google Scholar] [CrossRef]
- Kim, E.-S.; Kang, S.-Y.; Kim, Y.-H.; Lee, Y.-E.; Choi, N.-Y.; You, Y.-O.; Kim, K.-J. Chamaecyparis obtusa Essential Oil Inhibits Methicillin-Resistant Staphylococcus aureus Biofilm Formation and Expression of Virulence Factors. J. Med. Food 2015, 18, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zheng, Y.; Gao, H.; Xu, P.; Wang, M.; Li, A.; Miao, M.; Xie, X.; Deng, Y.; Zhou, H.; et al. Identification and Characterization of Staphylococcus aureus Strains with an Incomplete Hemolytic Phenotype. Front. Cell Infect. Microbiol. 2016, 6, 146. [Google Scholar] [CrossRef] [PubMed]
- Da, F.; Yao, L.; Su, Z.; Hou, Z.; Li, Z.; Xue, X.; Meng, J.; Luo, X. Antisense locked nucleic acids targeting agrA inhibit quorum sensing and pathogenesis of community-associated methicillin-resistant Staphylococcus aureus. J. Appl. Microbiol. 2017, 122, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Nimmo, G.R. USA300 abroad: Global spread of a virulent strain of community-associated methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2012, 18, 725–734. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Primer Sequences (5′–3′) | |
|---|---|---|
| Forward Primer | Reverse Primer | |
| hla | TTGGTGCAAATGTTTC | TCACTTTCCAGCCTACT |
| sea | ATGGTGCTTATTATGGTTATC | ATGGTGCTTATTATGGTTATC |
| seb | TGTTCGGGTATTTGAAGATGG | CGTTTCATAAGGCGAGTTGTT |
| agrA | TGATAATCCTTATGAGGTGC | CACTGTGACTCGTAACGAAA |
| 16s rRNA | CGTGCTACAATGGACAATAC | ATCTACGATTACTAGCGATT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-S.; Kim, K.-W.; Yoon, D.; Kim, G.-S.; Kwon, D.-Y.; Kang, O.-H.; Lee, D.Y. Comparison of Antivirulence Activities of Black Ginseng against Methicillin-Resistant Staphylococcus aureus According to the Number of Repeated Steaming and Drying Cycles. Antibiotics 2021, 10, 617. https://doi.org/10.3390/antibiotics10060617
Lee Y-S, Kim K-W, Yoon D, Kim G-S, Kwon D-Y, Kang O-H, Lee DY. Comparison of Antivirulence Activities of Black Ginseng against Methicillin-Resistant Staphylococcus aureus According to the Number of Repeated Steaming and Drying Cycles. Antibiotics. 2021; 10(6):617. https://doi.org/10.3390/antibiotics10060617
Chicago/Turabian StyleLee, Young-Seob, Kwan-Woo Kim, Dahye Yoon, Geum-Soog Kim, Dong-Yeul Kwon, Ok-Hwa Kang, and Dae Young Lee. 2021. "Comparison of Antivirulence Activities of Black Ginseng against Methicillin-Resistant Staphylococcus aureus According to the Number of Repeated Steaming and Drying Cycles" Antibiotics 10, no. 6: 617. https://doi.org/10.3390/antibiotics10060617
APA StyleLee, Y.-S., Kim, K.-W., Yoon, D., Kim, G.-S., Kwon, D.-Y., Kang, O.-H., & Lee, D. Y. (2021). Comparison of Antivirulence Activities of Black Ginseng against Methicillin-Resistant Staphylococcus aureus According to the Number of Repeated Steaming and Drying Cycles. Antibiotics, 10(6), 617. https://doi.org/10.3390/antibiotics10060617