
Seconeolitsine, the Novel Inhibitor of DNA Topoisomerase I, Protects against Invasive Pneumococcal Disease Caused by Fluoroquinolone-Resistant Strains

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates, Growth, Typing, and Susceptibility Tests
2.2. Animal Model Experiments
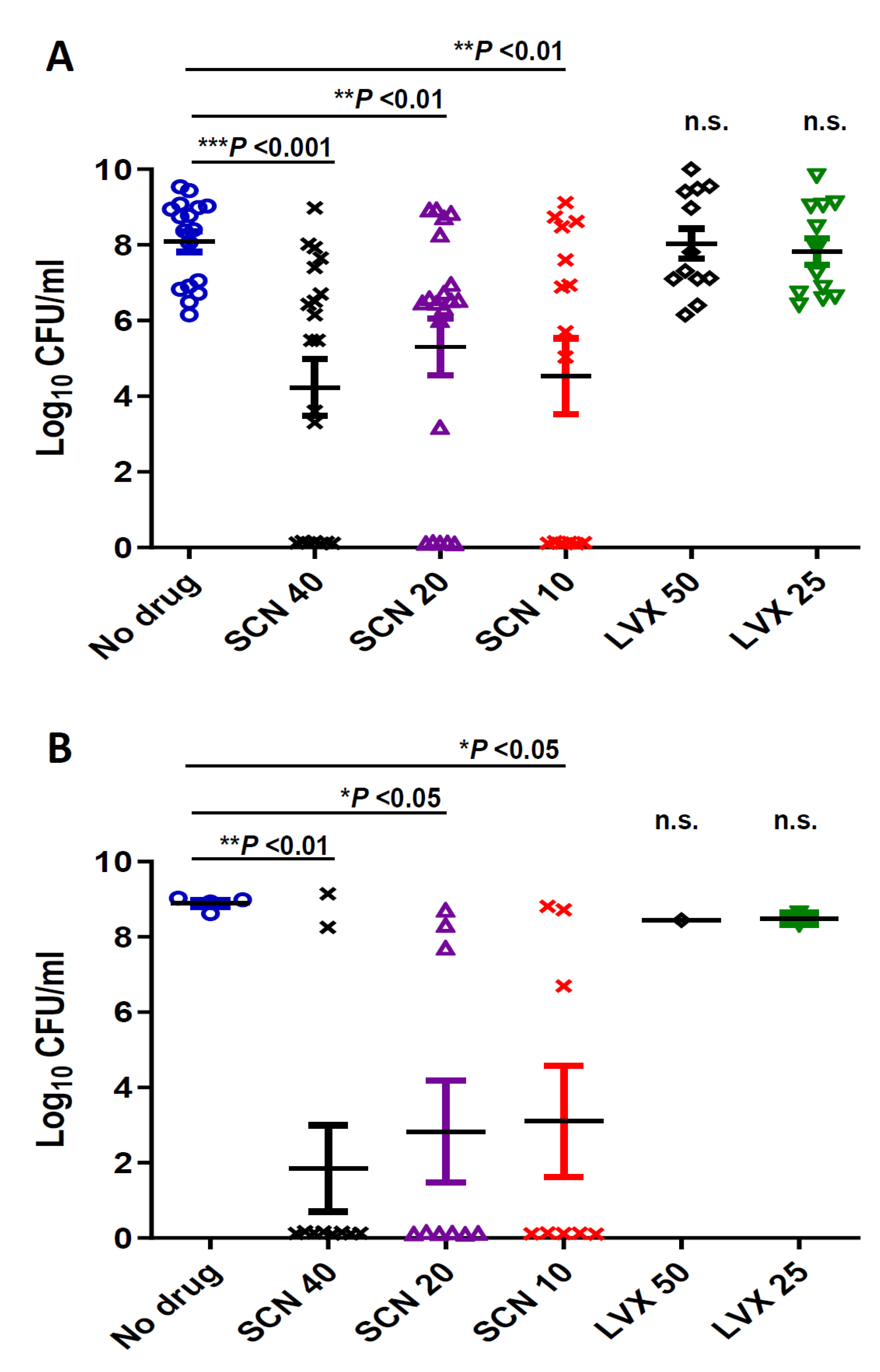
2.3. Determination of Bacteria in Blood
2.4. Determination Antibiotic Concentrations in Serum
2.5. Protein Binding
2.6. Drug Pharmacokinetics and Pharmacokinetic/Pharmacodynamics (PK/PD) Parameters
2.7. Statistical Analysis
2.8. Ethics Statement
3. Results
3.1. Virulence among FQ-Resistant Clinical Isolates Varies within Serotype
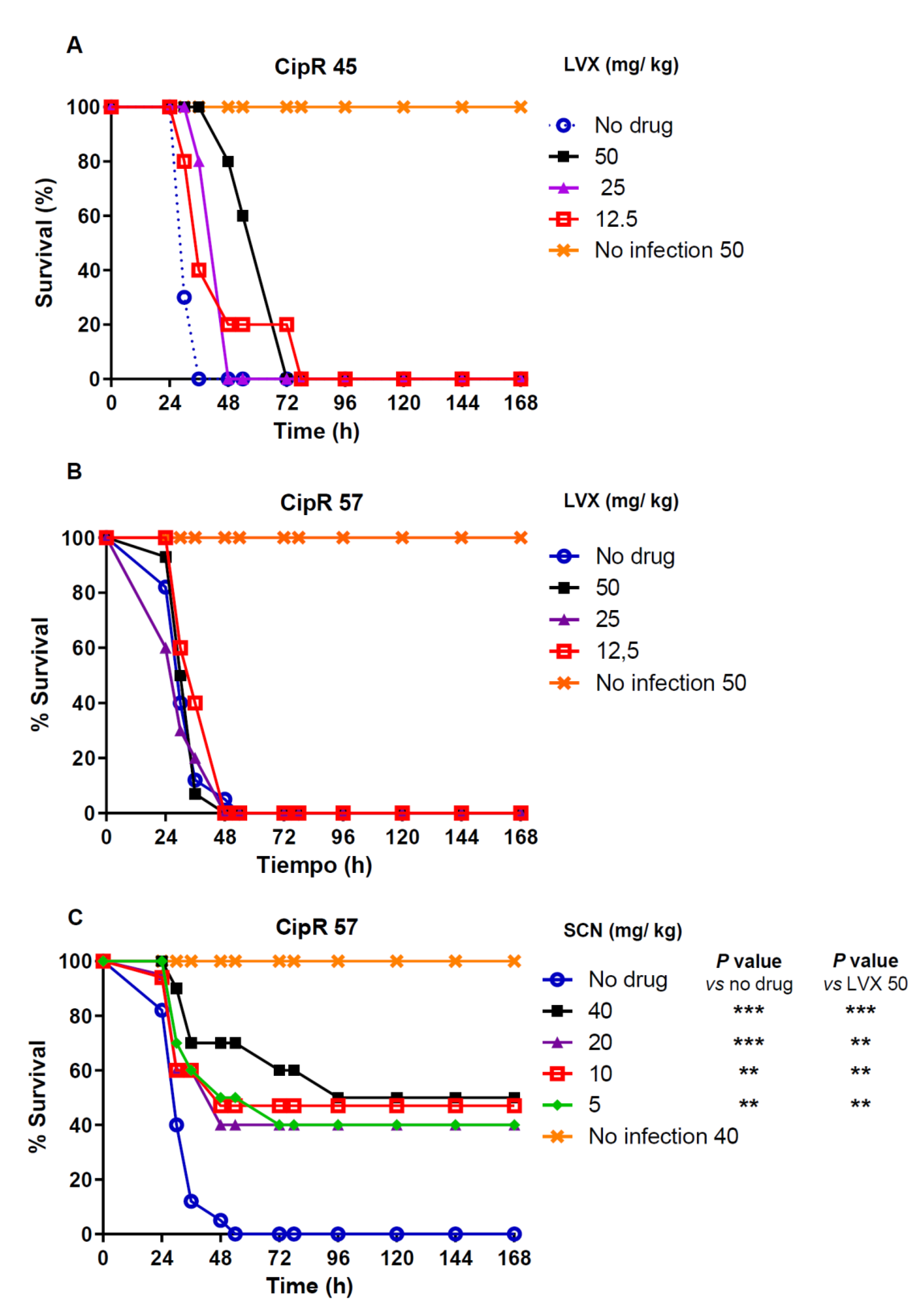
3.2. In Vivo Efficacy Studies
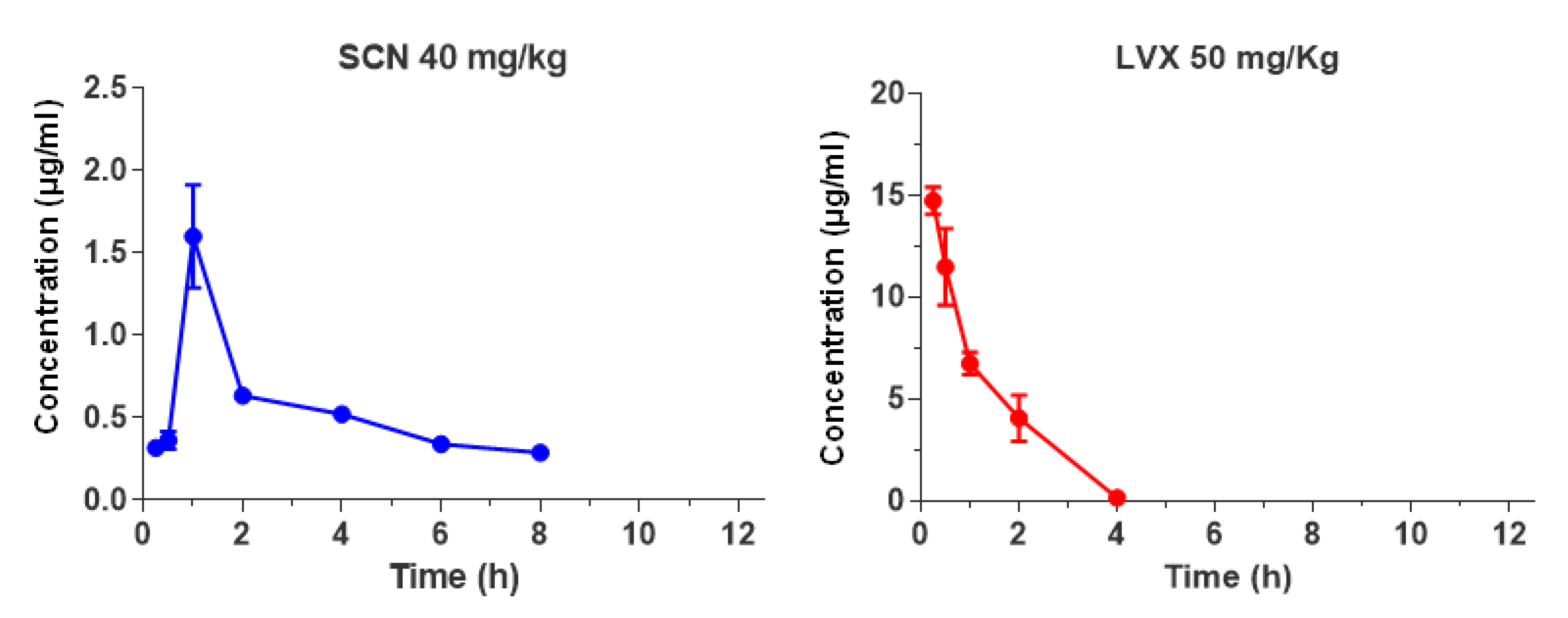
3.3. Pharmacokinetic and PK/PD Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2016 Lower Respiratory Infections Collaborators. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1191–1210. [Google Scholar] [CrossRef]
- Wahl, B.; O’Brien, K.L.; Greenbaum, A.; Majumder, A.; Liu, L.; Chu, Y.; Luksic, I.; Nair, H.; McAllister, D.A.; Campbell, H.; et al. Burden of Streptococcus pneumoniae and Haemophilus influenzae type b disease in children in the era of conjugate vaccines: Global, regional, and national estimates for 2000-15. Lancet Glob. Health 2018, 6, e744–e757. [Google Scholar] [CrossRef]
- O’Brien, K.L.; Wolfson, L.J.; Watt, J.P.; Henkle, E.; Deloria-Knoll, M.; McCall, N.; Lee, E.; Mulholland, K.; Levine, O.S.; Cherian, T. Burden of disease caused by Streptococcus pneumoniae in children younger than 5 years: Global estimates. Lancet 2009, 374, 893–902. [Google Scholar] [CrossRef]
- Pilishvili, T.; Lexau, C.; Farley, M.M.; Hadler, J.; Harrison, L.H.; Bennett, N.M.; Reingold, A.; Thomas, A.; Schaffner, W.; Craig, A.S.; et al. Sustained reductions in invasive pneumococcal disease in the era of conjugate vaccine. J. Infect. Dis. 2010, 201, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Fenoll, A.; Granizo, J.J.; Giménez, M.J.; Yuste, J.; Aguilar, L. Secular trends (1990–2013) in serotypes and associated non-susceptibility of S. pneumoniae isolates causing invasive disease in the pre-/post-era of pneumococcal conjugate vaccines in Spanish regions without universal paediatric pneumococcal vaccination. Vaccine 2015, 33, 5691–5699. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.R.; Link-Gelles, R.; Schaffner, W.; Lynfield, R.; Lexau, C.; Bennett, N.M.; Petit, S.; Zansky, S.M.; Harrison, L.H.; Reingold, A.; et al. Effect of use of 13-valent pneumococcal conjugate vaccine in children on invasive pneumococcal disease in children and adults in the USA: Analysis of multisite, population-based surveillance. Lancet Infect. Dis. 2015, 15, 301–309. [Google Scholar] [CrossRef]
- Ladhani, S.N.; Collins, S.; Djennad, A.; Sheppard, C.L.; Borrow, R.; Fry, N.K.; Andrews, N.J.; Miller, E.; Ramsay, M.E. Rapid increase in non-vaccine serotypes causing invasive pneumococcal disease in England and Wales, 2000–2017: A prospective national observational cohort study. Lancet Infect. Dis. 2018, 18, 441–451. [Google Scholar] [CrossRef]
- de Miguel, S.; Domenech, M.; González-Camacho, F.; Sempere, J.; Vicioso, D.; Sanz, J.C.; García Comas, L.; Ardanuy, C.; Fenoll, A.; Yuste, J. Nationwide trends of invasive pneumococcal disease in Spain (2009-2019) in children and adults during the pneumococcal conjugate vaccine era. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Ouldali, N.; Varon, E.; Levy, C.; Angoulvant, F.; Georges, S.; Ploy, M.C.; Kempf, M.; Cremniter, J.; Cohen, R.; Bruhl, D.L.; et al. Invasive pneumococcal disease incidence in children and adults in France during the pneumococcal conjugate vaccine era: An interrupted time-series analysis of data from a 17-year national prospective surveillance study. Lancet Infect. Dis. 2020, 121, 137–147. [Google Scholar] [CrossRef]
- Jacobs, M.R.; Felmingham, D.; Appelbaum, P.C.; Gruneberg, R.N.; Alexander Project Group. The Alexander Project 1998–2000: Susceptibility of pathogens isolated from community-acquired respiratory tract infection to commonly used antimicrobial agents. J. Antimicrob. Chemother. 2003, 52, 229–246. [Google Scholar] [CrossRef]
- Champoux, J.J. DNA topoisomerases: Structure, function, and mechanism. Annu. Rev. Biochem. 2001, 70, 369–413. [Google Scholar] [CrossRef]
- Mandell, L.A.; Wunderink, R.G.; Anzueto, A.; Bartlett, J.G.; Campbell, G.D.; Dean, N.C.; Dowell, S.F.; File, T.M., Jr.; Musher, D.M.; Niederman, M.S.; et al. Infectious Diseases Society of America/American Thoracic Society consensus guidelines on the management of community-acquired pneumonia in adults. Clin. Infect. Dis. 2007, 44 (Suppl. 2), S27–S72. [Google Scholar] [CrossRef] [PubMed]
- Riedel, S.; Beekmann, S.E.; Heilmann, K.P.; Richter, S.S.; García-de-Lomas, J.; Ferech, M.; Goosens, H.; Doern, G.V. Antimicrobial use in Europe and antimicrobial resistance in Streptococcus pneumoniae. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 485–490. [Google Scholar] [CrossRef]
- Domenech, A.; Tirado-Vélez, J.M.; Fenoll, A.; Ardanuy, C.; Yuste, J.; Linares, J.; de la Campa, A.G. Fluoroquinolone-resistant pneumococci: Dynamics of serotypes and clones in Spain in 2012 compared with those from 2002 and 2006. Antimicrob. Agents Chemother. 2014, 58, 2393–2399. [Google Scholar] [CrossRef] [PubMed]
- Fuller, J.D.; Low, D.E. A review of Streptococcus pneumoniae infection treatment failures associated with fluoroquinolone resistance. Clin. Infect. Dis. 2005, 41, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Adam, H.J.; Hoban, D.J.; Gin, A.S.; Zhanel, G.G. Association between fluoroquinolone usage and a dramatic rise in ciprofloxacin-resistant Streptococcus pneumoniae in Canada, 1997–2006. Int. J. Antimicrob. Agents 2009, 34, 82–85. [Google Scholar] [CrossRef]
- Chen, D.K.; McGeer, A.; de Azavedo, J.C.; Low, D.E. Decreased susceptibility of Streptococcus pneumoniae to fluoroquinolones in Canada. Canadian Bacterial Surveillance Network. N. Engl. J. Med. 1999, 341, 233–239. [Google Scholar] [CrossRef]
- Cheng, B.; Liu, I.F.; Tse-Dinh, Y.C. Compounds with antibacterial activity that enhance DNA cleavage by bacterial DNA topoisomerase I. J. Antimicrob. Chemother. 2007, 59, 640–645. [Google Scholar] [CrossRef]
- García, M.T.; Blázquez, M.A.; Ferrándiz, M.J.; Sanz, M.J.; Silva-Martín, N.; Hermoso, J.A.; de la Campa, A.G. New alkaloid antibiotics that target the DNA topoisomerase I of Streptococcus pneumoniae. J. Biol. Chem. 2011, 286, 6402–6413. [Google Scholar] [CrossRef]
- Ferrándiz, M.J.; Martín-Galiano, A.J.; Arnanz, C.; Camacho-Soguero, I.; Tirado-Vélez, J.M.; de la Campa, A.G. An increase in negative supercoiling in bacteria reveals topology-reacting gene clusters and a homeostatic response mediated by the DNA topoisomerase I gene. Nucleic Acids Res. 2016, 44, 7292–7303. [Google Scholar] [CrossRef]
- García, M.T.; Carreno, D.; Tirado-Vélez, J.M.; Ferrándiz, M.J.; Rodrigues, L.; Gracia, B.; Amblar, M.; Ainsa, J.A.; de la Campa, A.G. Boldine-derived alkaloids inhibit the activity of DNA topoisomerase I and growth of Mycobacterium tuberculosis. Front. Microbiol. 2018, 9, 1659. [Google Scholar] [CrossRef]
- Valenzuela, M.V.; Domenech, M.; Mateos-Martínez, P.; González-Camacho, F.; de la Campa, A.G.; García, M.T. Antibacterial activity of a DNA topoisomerase I inhibitor versus fluoroquinolones in Streptococcus pneumoniae. PLoS ONE 2020, 15, e0241780. [Google Scholar] [CrossRef] [PubMed]
- Fenoll, A.; Jado, I.; Vicioso, D.; Casal, J. Dot blot assay for the serotyping of pneumococci. J. Clin. Microbiol. 1997, 35, 764–766. [Google Scholar] [CrossRef] [PubMed]
- Elberse, K.E.; van de Pol, I.; Witteveen, S.; van der Heide, H.G.; Schot, C.S.; van Dijk, A.; van der Ende, A.; Schouls, L.M. Population structure of invasive Streptococcus pneumoniae in The Netherlands in the pre-vaccination era assessed by MLVA and capsular sequence typing. PLoS ONE 2011, 6, e20390. [Google Scholar] [CrossRef] [PubMed]
- Enright, M.C.; Spratt, B.G. A multilocus sequence typing scheme for Streptococcus pneumoniae: Identification of clones associated with serious invasive disease. Microbiology 1998, 144, 3049–3060. [Google Scholar] [CrossRef] [PubMed]
- Ardanuy, C.; de la Campa, A.G.; García, E.; Fenoll, A.; Calatayud, L.; Cercenado, E.; Pérez-Trallero, E.; Bouza, E.; Linares, J. Spread of Streptococcus pneumoniae serotype 8-ST63 multidrug-resistant recombinant Clone, Spain. Emerg. Infect. Dis. 2014, 20, 1848–1856. [Google Scholar] [CrossRef]
- de la Campa, A.G.; Ardanuy, C.; Balsalobre, L.; Pérez-Trallero, E.; Marimón, J.M.; Fenoll, A.; Linares, J. Changes in fluoroquinolone-resistant Streptococcus pneumoniae after 7-valent conjugate vaccination, Spain. Emerg. Infect. Dis. 2009, 15, 905–911. [Google Scholar] [CrossRef]
- Cafini, F.; Yuste, J.; Giménez, M.J.; Sevillano, D.; Aguilar, L.; Alou, L.; Ramos-Sevillano, E.; Torrico, M.; González, N.; García, E.; et al. Enhanced in vivo activity of cefditoren in pre-immunized mice against penicillin-resistant S. pneumoniae (serotypes 6B, 19F and 23F) in a sepsis model. PLoS ONE 2010, 5, e12041. [Google Scholar] [CrossRef]
- Ramos-Sevillano, E.; Urzainqui, A.; de Andrés, B.; González-Tajuelo, R.; Domenech, M.; González-Camacho, F.; Sánchez-Madrid, F.; Brown, J.S.; García, E.; Yuste, J. PSGL-1 on leukocytes is a critical component of the host immune response against invasive pneumococcal disease. PLoS Pathog. 2016, 12, e1005500. [Google Scholar] [CrossRef]
- Ramos-Sevillano, E.; Urzainqui, A.; Campuzano, S.; Moscoso, M.; González-Camacho, F.; Domenech, M.; Rodríguez de Córdoba, S.; Sánchez-Madrid, F.; Brown, J.S.; García, E.; et al. Pleiotropic effects of cell wall amidase LytA on Streptococcus pneumoniae sensitivity to the host immune response. Infect. Immun. 2015, 83, 591–603. [Google Scholar] [CrossRef]
- Yuste, J.; Fenoll, A.; Casal, J.; Giménez, M.J.; Aguilar, L. Combined effect of specific antibodies (as serotherapy or preimmunization) and amoxicillin doses in treatment of Streptococcus pneumoniae sepsis in a mouse model. Antimicrob. Agents Chemother. 2002, 46, 4043–4044. [Google Scholar] [CrossRef]
- Tarragó, D.; Lara, N.; Fenoll, A.; Casal, J.; Giménez, M.J.; Aguilar, L.; Sevillano, D. Specific antibodies, levofloxacin, and modulation of capsule-associated virulence in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2005, 49, 3095–3096. [Google Scholar] [CrossRef]
- Yuste, J.; Jado, I.; Fenoll, A.; Aguilar, L.; Giménez, M.J.; Casal, J. Beta-lactam modification of the bacteraemic profile and its relationship with mortality in a pneumococcal mouse sepsis model. J. Antimicrob. Chemother. 2002, 49, 331–335. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Alkorta, M.; Giménez, M.J.; Vicente, D.; Aguilar, L.; Pérez-Trallero, E. In vivo activity of gemifloxacin, moxifloxacin and levofloxacin against pneumococci with gyrA and parC point mutations in a sepsis mouse model measured with the all or nothing mortality end-point. Int. J. Antimicrob. Agents 2005, 25, 163–167. [Google Scholar] [CrossRef]
- Casal, J.; Aguilar, L.; Jado, I.; Yuste, J.; Giménez, M.J.; Prieto, J.; Fenoll, A. Effects of specific antibodies against Streptococcus pneumoniae on pharmacodynamic parameters of beta-lactams in a mouse sepsis model. Antimicrob. Agents Chemother. 2002, 46, 1340–1344. [Google Scholar] [CrossRef]
- Huelves, L.; Sevillano, D.; Martínez-Marín, C.; López-Casla, M.T.; Gracia, M.; Alou, L.; del Carmen Ponte, M.; Prieto, J.; Soriano, F.; Spanish Pneumococcal Infection Study, N. Correlation between in vitro and in vivo activity of levofloxacin and moxifloxacin against pneumococcal strains with different susceptibilities to fluoroquinolones. Int. J. Antimicrob. Agents 2006, 27, 294–299. [Google Scholar] [CrossRef]
- Onyeji, C.O.; Bui, K.Q.; Owens, R.C., Jr.; Nicolau, D.P.; Quintiliani, R.; Nightingale, C.H. Comparative efficacies of levofloxacin and ciprofloxacin against Streptococcus pneumoniae in a mouse model of experimental septicaemia. Int. J. Antimicrob. Agents 1999, 12, 107–114. [Google Scholar] [CrossRef]
- Yuste, J.; Botto, M.; Bottoms, S.E.; Brown, J.S. Serum amyloid P aids complement-mediated immunity to Streptococcus pneumoniae. PLoS Pathog 2007, 3, 1208–1219. [Google Scholar] [CrossRef] [PubMed]
- Alou, L.; Giménez, M.J.; Sevillano, D.; Aguilar, L.; Cafini, F.; Echeverría, O.; Pérez-Trallero, E.; Prieto, J. A pharmacodynamic approach to antimicrobial activity in serum and epithelial lining fluid against in vivo-selected Streptococcus pneumoniae mutants and association with clinical failure in pneumonia. J. Antimicrob. Chemother. 2006, 58, 349–358. [Google Scholar] [CrossRef]
- Sevillano, D.; Aguilar, L.; Alou, L.; Giménez, M.J.; González, N.; Torrico, M.; Cafini, F.; Fenoll, A.; Coronel, P.; Prieto, J. High protein binding and cidal activity against penicillin-resistant S. pneumoniae: A cefditoren in vitro pharmacodynamic simulation. PLoS ONE 2008, 3, e2717. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Avial, I.; Ramos, B.; Ríos, E.; Cercenado, E.; Ordobás, M.; Sanz, J.C.; Madrid Streptococcus pneumoniae Microbiological, Group. Clonal spread of levofloxacin-resistant Streptococcus pneumoniae invasive isolates in Madrid, Spain, 2007 to 2009. Antimicrob. Agents Chemother. 2011, 55, 2469–2471. [Google Scholar] [CrossRef]
- Sanz, J.C.; Cercenado, E.; Marín, M.; Ramos, B.; Ardanuy, C.; Rodríguez-Avial, I.; Bouza, E. Multidrug-resistant pneumococci (serotype 8) causing invasive disease in HIV+ patients. Clin. Microbiol. Infect. 2011, 17, 1094–1098. [Google Scholar] [CrossRef] [PubMed]
- Amin-Chowdhury, Z.; Collins, S.; Sheppard, C.; Litt, D.; Fry, N.K.; Andrews, N.; Ladhani, S.N. Characteristics of invasive pneumococcal disease caused by emerging serotypes after the introduction of the 13-valent pneumococcal conjugate vaccine in England: A prospective observational cohort study, 2014–2018. Clin. Infect. Dis. 2020, 71, e235–e243. [Google Scholar] [CrossRef]
- Shoji, H.; Vázquez-Sánchez, D.A.; González-Díaz, A.; Cubero, M.; Tubau, F.; Santos, S.; García-Somoza, D.; Linares, J.; Yuste, J.; Martí, S.; et al. Overview of pneumococcal serotypes and genotypes causing diseases in patients with chronic obstructive pulmonary disease in a Spanish hospital between 2013 and 2016. Infect. Drug Resist. 2018, 11, 1387–1400. [Google Scholar] [CrossRef] [PubMed]
- Craig, W.; Andes, D.A. Correlation of the magnitude of the AUC24/MIC for 6 fluoroquinolones against Streptococcus pneumoniae with survival and bactericidal activity in an animal model [abstract 289]. In Proceedings of the Program and abstracts of the 40th Interscience Conference of Antimicrobial Chemotherapy (Toronto), Toronto, ON, Canada, 17–20 September 2000; American Society for Microbiology: Washington, DC, USA, 2000; Volume 7. [Google Scholar]
- Ambrose, P.G.; Bhavnani, S.M.; Rubino, C.M.; Louie, A.; Gumbo, T.; Forrest, A.; Drusano, G.L. Pharmacokinetics-pharmacodynamics of antimicrobial therapy: It’s not just for mice anymore. Clin. Infect. Dis. 2007, 44, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Bowker, K.E.; Garvey, M.I.; Noel, A.R.; Tomaselli, S.G.; Macgowan, A.P. Comparative antibacterial effects of moxifloxacin and levofloxacin on Streptococcus pneumoniae strains with defined mechanisms of resistance: Impact of bacterial inoculum. J. Antimicrob. Chemother. 2013, 68, 1130–1138. [Google Scholar] [CrossRef][Green Version]
- Hoet, S.; Stevigny, C.; Block, S.; Opperdoes, F.; Colson, P.; Baldeyrou, B.; Lansiaux, A.; Bailly, C.; Quetin-Leclercq, J. Alkaloids from Cassytha filiformis and related aporphines: Antitrypanosomal activity, cytotoxicity, and interaction with DNA and topoisomerases. Planta Med. 2004, 70, 407–413. [Google Scholar] [CrossRef]
- Sobarzo-Sánchez, E.; Soto, P.G.; Valdés Rivera, C.; Sánchez, G.; Hidalgo, M.E. Applied biological and physicochemical activity of isoquinoline alkaloids: Oxoisoaporphine and boldine. Molecules 2012, 17, 10958–10970. [Google Scholar] [CrossRef]
- Ferrándiz, M.J.; Hernández, P.; de la Campa, A.G. Genome-wide proximity between RNA polymerase and DNA topoisomerase I supports transcription in Streptococcus pneumoniae. PLoS Genet. 2021, 17, e1009542. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Isolate | Type a | MIC (µg/ mL) b: | Pattern c | QRDR Statuses d: | MLD e | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| SCN | CIP | LVX | MXF | ParC | ParE | GyrA | ||||
| CipR5 | 33F | 5.2 | 16 | 8 | 2.00 | EClCip | D83N | None | E85K | Avirulent |
| CipR15 | 16F | 2.6 | 32 | 16 | 4.00 | SxTCip | S79F | None | S81F | Avirulent |
| CipR45 | 8 | 5.2 | 4 | 2 | 0.25 | TEClCip | S79F | None | None | ≥5 × 101 |
| CipR57 | 8 | 2.6 | 64 | 16 | 4.00 | TECip | S79F | None | S81F | ≥5 × 101 |
| CipR72 | 15A | 5.2 | 64 | 32 | 4.00 | PTEClCip | S79F | E474K | S81F | Avirulent |
| PK Parameters | Seconeolitsine | Levofloxacin | p Values |
|---|---|---|---|
| Cmax (µg/ mL) | 1.6 ± 0.3 | 14.7 ± 0.7 | <0.01 |
| Cmin(µg/ mL) | 0.3 ± 0.04 | 0.1 ± 0.0 | <0.05 |
| Tmax (h) | 1.0 ± 0.0 | 0.25 ± 0.0 | <0.001 |
| t1/2 (h) | 7.8 ± 2.4 | 0.7 ± 0.0 | 0.054 |
| Tlast (h) | 9.0 ± 1.4 | 4.0 ± 0.0 | <0.5 |
| AUC0-12h (µg/mL×h) | 5.0 ± 0.0 | 17.3 ± 1.5 | <0.01 |
| AUC0-24h (µg/mL×h) | 10.0 ± 0.1 | 34.6 ± 2.9 | <0.01 |
| PK/PD parameters | |||
| Strain CipR45; MIC (µg/ mL) | 5.2 | 2 | |
| fAUC/MIC (h) | 1.61 ± 0.0 | 15.2 ± 1.3 | <0.01 |
| fT>MIC (%) | 0.0 | 24.2 ± 2.8 | - |
| Strain CipR57; MIC (µg/ mL) | 2.6 | 16 | |
| fAUC/MIC (h) | 2.3 ± 0.0 | 1.9 ± 0.16 | 0.074 |
| fT>MIC (%) | 0.0 | 0.0 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tirado-Vélez, J.M.; Carreño, D.; Sevillano, D.; Alou, L.; Yuste, J.; de la Campa, A.G. Seconeolitsine, the Novel Inhibitor of DNA Topoisomerase I, Protects against Invasive Pneumococcal Disease Caused by Fluoroquinolone-Resistant Strains. Antibiotics 2021, 10, 573. https://doi.org/10.3390/antibiotics10050573
Tirado-Vélez JM, Carreño D, Sevillano D, Alou L, Yuste J, de la Campa AG. Seconeolitsine, the Novel Inhibitor of DNA Topoisomerase I, Protects against Invasive Pneumococcal Disease Caused by Fluoroquinolone-Resistant Strains. Antibiotics. 2021; 10(5):573. https://doi.org/10.3390/antibiotics10050573
Chicago/Turabian StyleTirado-Vélez, Jose Manuel, David Carreño, David Sevillano, Luis Alou, José Yuste, and Adela G. de la Campa. 2021. "Seconeolitsine, the Novel Inhibitor of DNA Topoisomerase I, Protects against Invasive Pneumococcal Disease Caused by Fluoroquinolone-Resistant Strains" Antibiotics 10, no. 5: 573. https://doi.org/10.3390/antibiotics10050573
APA StyleTirado-Vélez, J. M., Carreño, D., Sevillano, D., Alou, L., Yuste, J., & de la Campa, A. G. (2021). Seconeolitsine, the Novel Inhibitor of DNA Topoisomerase I, Protects against Invasive Pneumococcal Disease Caused by Fluoroquinolone-Resistant Strains. Antibiotics, 10(5), 573. https://doi.org/10.3390/antibiotics10050573