
Comparison of Two Transmission Electron Microscopy Methods to Visualize Drug-Induced Alterations of Gram-Negative Bacterial Morphology

 , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Antibiotics and Chemicals
2.2. Bacterial Strains and Growth Conditions
2.3. Transmission Electron Microscopy
2.3.1. Xen14 Processing for TEM
2.3.2. Xen41 Processing for TEM
2.3.3. Cryo-Ultramicrotomy
2.4. Treated Samples Processing for TEM and Cryo-Ultramicrotomy
3. Results and Discussion
3.1. Bacterial Cell Morphology Is Affected by the Fixative Used, Buffer Conditions and the Embedding Method
- Fixative: Fixative containing 3.0% formaldehyde, 0.035% glutaraldehyde, 4% sucrose and 0.075% ruthenium red, 0.075% L-lysine acetate (Procedure 1) caused shrinkage of the bacterial cell as well as detachment and perturbation of the cell membrane (Figure 2A,B). Therefore, the low concentration of formaldehyde and glutaraldehyde used in this procedure was not high enough to preserve cell membrane structure and could have affected the cell size. To circumvent this, Li, et al. [28] described a 4.0% formaldehyde solution in fixative as optimal for preservation of bacterial cell size, and our result supports this observation.
- Buffer: We also found that the type of buffer used resulted in altered cell membrane morphology. PBS buffer caused detachment of the cell membrane (Figure 2C,D; Procedure 2), a similar observation as those described by others [14,29]. Furthermore, the addition of sucrose to the buffer improved preservation of cell morphology, as the cell membrane appeared brittle if sucrose was omitted from the fixative (Figure 2E,F; Procedure 3), in agreement with a previous study [30].
- Embedding method: Following from the optimized fixative and buffer conditions above (Procedure 2), we observed that a TEM protocol using cacodylate buffer with fixative containing 4.0% formaldehyde, 1.25% glutaraldehyde, 0.075% ruthenium red, 0.075% L-lysine acetate, and 4% sucrose followed by embedding in LR-White resin (Procedure 4) provided the best delineation of the outer membrane, cell wall and inner membrane, with no wavy, detached or shrunk membranes (Figure 3A). This protocol is similar in some respects to that described by Voget et al. [14], but differs in the buffer and fixative composition, treatment time and embedding method. The use of Epon-Araldite resin (Procedure 5) did not appear to make a difference to the overall TEM result (Figure 3A vs. Figure 3B). However, given our findings, we suggest using LR-White resin due to its ease of use during TEM processing.
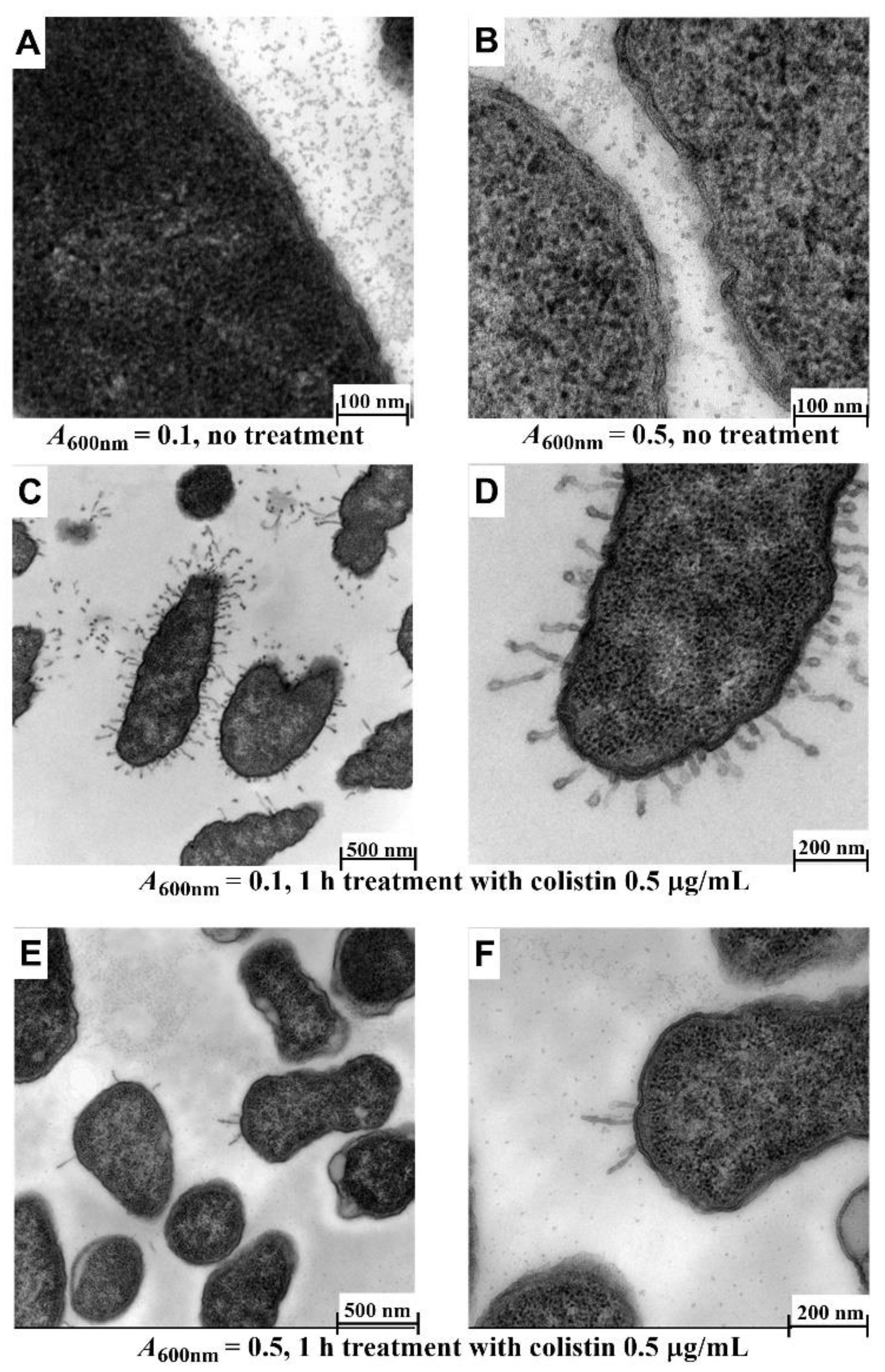
3.2. Bacterial Cell Morphology Is Affected by Cell Density and Exposure Time to Drugs
3.3. Comparison of TEM and Cryo-Ultramicrotomy for Visualizing NCL195-Colistin Interaction on Cell Membrane
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zabawa, T.P.; Pucci, M.J.; Parr, T.R., Jr.; Lister, T. Treatment of Gram-negative bacterial infections by potentiation of antibiotics. Curr. Opin. Microbiol. 2016, 33, 7–12. [Google Scholar] [CrossRef]
- Zgurskaya, H.I.; Lopez, C.A.; Gnanakaran, S. Permeability barrier of Gram-negative cell envelopes and approaches to bypass it. ACS Infect. Dis. 2015, 1, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Lenhard, J.R.; Nation, R.L.; Tsuji, B.T. Synergistic combinations of polymyxins. Int. J. Antimicrob. Agents 2016, 48, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Khazandi, M.; Pi, H.; Chan, W.Y.; Ogunniyi, A.D.; Sim, J.X.F.; Venter, H.; Garg, S.; Page, S.W.; Hill, P.B.; McCluskey, A. In vitro antimicrobial activity of robenidine, ethylenediaminetetraacetic acid and polymyxin B nonapeptide against important human and veterinary pathogens. Front. Microbiol. 2019, 10, 837. [Google Scholar] [CrossRef] [PubMed]
- Brennan-Krohn, T.; Pironti, A.; Kirby, J.E. Synergistic activity of colistin-containing combinations against colistin-resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, e00873-18. [Google Scholar] [CrossRef]
- Elemam, A.; Rahimian, J.; Doymaz, M. In vitro evaluation of antibiotic synergy for polymyxin B-resistant carbapenemase-producing Klebsiella pneumoniae. J. Clin. Microbiol. 2010, 48, 3558–3562. [Google Scholar] [CrossRef] [PubMed]
- Garonzik, S.; Li, J.; Thamlikitkul, V.; Paterson, D.; Shoham, S.; Jacob, J.; Silveira, F.; Forrest, A.; Nation, R.L. Population pharmacokinetics of colistin methanesulfonate and formed colistin in critically ill patients from a multicenter study provide dosing suggestions for various categories of patients. Antimicrob. Agents Chemother. 2011, 55, 3284–3294. [Google Scholar] [CrossRef] [PubMed]
- Nigam, A.; Kumari, A.; Jain, R.; Batra, S. Colistin neurotoxicity: Revisited. BMJ Case Rep. 2015, 2015, bcr2015210787. [Google Scholar] [CrossRef] [PubMed]
- Otto, R.G.; van Gorp, E.; Kloezen, W.; Meletiadis, J.; van den Berg, S.; Mouton, J.W. An alternative strategy for combination therapy: Interactions between polymyxin B and non-antibiotics. Int. J. Antimicrob. Agents 2019, 53, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Venter, H.; Veltman, T.; Williams, R.; O’Donovan, L.A.; Russell, C.C.; McCluskey, A.; Page, S.W.; Ogunniyi, A.D.; Trott, D.J. In vitro synergistic activity of NCL195 in combination with colistin against Gram-negative bacterial pathogens. Int. J. Antimicrob. Agents 2021. under review. [Google Scholar]
- Pi, H.; Nguyen, H.T.; Venter, H.; Boileau, A.R.; Woolford, L.; Garg, S.; Page, S.W.; Russell, C.C.; Baker, J.R.; McCluskey, A.; et al. In vitro activity of robenidine analog NCL195 in combination with outer membrane permeabilizers against Gram-negative bacterial pathogens and impact on systemic Gram-positive bacterial infection in mice. Front. Microbiol. 2020, 11, 1556. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Liu, Y.; Huang, X.; Ding, S.; Wang, Y.; Shen, J.; Zhu, K. A broad-spectrum antibiotic adjuvant reverses multidrug-resistant Gram-negative pathogens. Nat. Microbiol. 2020, 5, 1040–1050. [Google Scholar] [CrossRef]
- Ontong, J.C.; Ozioma, N.F.; Voravuthikunchai, S.P.; Chusri, S. Synergistic antibacterial effects of colistin in combination with aminoglycoside, carbapenems, cephalosporins, fluoroquinolones, tetracyclines, fosfomycin, and piperacillin on multidrug resistant Klebsiella pneumoniae isolates. PLoS ONE 2021, 16, e0244673. [Google Scholar] [CrossRef] [PubMed]
- Voget, M.; Lorenz, D.; Lieber-Tenorio, E.; Hauck, R.; Meyer, M.; Cieslicki, M. Is transmission electron microscopy (TEM) a promising approach for qualitative and quantitative investigations of polymyxin B and miconazole interactions with cellular and subcellular structures of Staphylococcus pseudintermedius, Escherichia coli, Pseudomonas aeruginosa and Malassezia pachydermatis? Vet. Microbiol. 2015, 181, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Stukalov, O.; Korenevsky, A.; Beveridge, T.J.; Dutcher, J.R. Use of atomic force microscopy and transmission electron microscopy for correlative studies of bacterial capsules. Appl. Environ. Microbiol. 2008, 74, 5457–5465. [Google Scholar] [CrossRef]
- Liao, S.; Zhang, Y.; Pan, X.; Zhu, F.; Jiang, C.; Liu, Q.; Cheng, Z.; Dai, G.; Wu, G.; Wang, L.; et al. Antibacterial activity and mechanism of silver nanoparticles against multidrug-resistant Pseudomonas aeruginosa. Int. J. Nanomed. 2019, 14, 1469–1487. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Shafi, S.; Al-Mohammadi, A.R.; Sitohy, M.; Mosa, B.; Ismaiel, A.; Enan, G.; Osman, A. Antimicrobial activity and chemical constitution of the crude, phenolic-rich extracts of Hibiscus sabdariffa, Brassica oleracea and Beta vulgaris. Molecules 2019, 24, 4280. [Google Scholar] [CrossRef] [PubMed]
- Rapisarda, C.; Tassinari, M.; Gubellini, F.; Fronzes, R. Using Cryo-EM to investigate bacterial secretion systems. Annu. Rev. Microbiol. 2018, 72, 231–254. [Google Scholar] [CrossRef] [PubMed]
- Egelman, E.H. The Current Revolution in Cryo-EM. Biophys. J. 2016, 110, 1008–1012. [Google Scholar] [CrossRef] [PubMed]
- Pilhofer, M.; Ladinsky, M.S.; McDowall, A.W.; Jensen, G.J. Bacterial TEM: New insights from cryo-microscopy. Methods Cell Biol. 2010, 96, 21–45. [Google Scholar] [CrossRef]
- Yan, R.; Venkatakrishnan, S.V.; Liu, J.; Bouman, C.A.; Jiang, W. MBIR: A cryo-ET 3D reconstruction method that effectively minimizes missing wedge artifacts and restores missing information. J. Struct. Biol. 2019, 206, 183–192. [Google Scholar] [CrossRef]
- Griffiths, G.; Slot, J.W.; Webster, P. Kiyoteru Tokuyasu: A pioneer of cryo-ultramicrotomy. Microscopy 2015, 64, 377–379. [Google Scholar] [CrossRef]
- Thompson, R.F.; Walker, M.; Siebert, C.A.; Muench, S.P.; Ranson, N.A. An introduction to sample preparation and imaging by cryo-electron microscopy for structural biology. Methods 2016, 100, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Abraham, R.J.; Stevens, A.J.; Young, K.A.; Russell, C.; Qvist, A.; Khazandi, M.; Wong, H.S.; Abraham, S.; Ogunniyi, A.D.; Page, S.W.; et al. Robenidine analogues as Gram-positive antibacterial agents. J. Med. Chem. 2016, 59, 2126–2138. [Google Scholar] [CrossRef]
- Ogunniyi, A.D.; Khazandi, M.; Stevens, A.J.; Sims, S.K.; Page, S.W.; Garg, S.; Venter, H.; Powell, A.; White, K.; Petrovski, K.R. Evaluation of robenidine analog NCL195 as a novel broad-spectrum antibacterial agent. PLoS ONE 2017, 12, e0183457. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, S.; Wolff, S.; Hocke, A.; Rosseau, S.; Müller, E.; Rohde, M. Illustration of pneumococcal polysaccharide capsule during adherence and invasion of epithelial cells. Infect. Immun. 2005, 73, 4653–4667. [Google Scholar] [CrossRef] [PubMed]
- Birkhead, M.; Ganesh, K.; Ndlangisa, K.M.; Koornhof, H.J. Transmission electron microscopy protocols for capsule visualisation in pathogenic respiratory and meningeal bacteria. In Microscopy and Imaging Science: Practical Approaches to Applied Research and Education; Researchgate: Berlin, Germany, 2017; pp. 628–639. [Google Scholar]
- Li, Y.; Almassalha, L.M.; Chandler, J.E.; Zhou, X.; Stypula-Cyrus, Y.E.; Hujsak, K.A.; Roth, E.W.; Bleher, R.; Subramanian, H.; Szleifer, I. The effects of chemical fixation on the cellular nanostructure. Exp. Cell Res. 2017, 358, 253–259. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Wu, T.; Pan, S.; Xu, X. Antimicrobial mechanism of flavonoids against Escherichia coli ATCC 25922 by model membrane study. Appl. Surf. Sci. 2014, 305, 515–521. [Google Scholar] [CrossRef]
- Leslie, S.B.; Israeli, E.; Lighthart, B.; Crowe, J.H.; Crowe, L.M. Trehalose and sucrose protect both membranes and proteins in intact bacteria during drying. Appl. Environ. Microbiol. 1995, 61, 3592–3597. [Google Scholar] [CrossRef] [PubMed]
- Bergen, P.J.; Smith, N.M.; Bedard, T.B.; Bulman, Z.P.; Cha, R.; Tsuji, B.T. Rational Combinations of Polymyxins with Other Antibiotics. In Polymyxin Antibiotics: From Laboratory Bench to Bedside; Springer: Berlin, Germany, 2019; pp. 251–288. [Google Scholar]
- Kwa, A.; Kasiakou, S.K.; Tam, V.H.; Falagas, M.E. Polymyxin B: Similarities to and differences from colistin (polymyxin E). Expert Rev. Anti-Infect. Ther. 2007, 5, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Möbius, W.; Posthuma, G. Sugar and ice: Immunoelectron microscopy using cryosections according to the Tokuyasu method. Tissue Cell 2019, 57, 90–102. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Buffer | Fixation | Post-fixation | Resin |
|---|---|---|---|---|
| Procedure 1 (TEM) | Cacodylate + 4% sucrose | 3.0% formaldehyde, 0.035% glutaraldehyde, 4% sucrose and 0.075% ruthenium red, 0.075% L-lysine acetate | 1% osmium tetroxide; 0.075% ruthenium red | LR-White |
| Procedure 2 (TEM) | PBS + 4% sucrose | 4.0% formaldehyde, 1.25% glutaraldehyde, 0.01 M CaCl2, 4% sucrose and 0.075% ruthenium red | 1% osmium tetroxide; 0.075% ruthenium red; | Epon-Araldite |
| Procedure 3 (TEM) | Cacodylate | 4.0% formaldehyde, 1.25% glutaraldehyde, 0.01 M CaCl2 and 0.075% ruthenium red, 0.075% L-lysine acetate | 1% osmium tetroxide; 0.075% ruthenium red | Epon-Araldite |
| Procedure 4 (TEM) | Cacodylate + 4% sucrose | 4.0% formaldehyde, 1.25% glutaraldehyde, 0.01 M CaCl2, 4% sucrose and 0.075% ruthenium red, 0.075% L-lysine acetate | 1% osmium tetroxide; 0.075% ruthenium red | LR-White |
| Procedure 5 (TEM) | Cacodylate + 4% sucrose | 4.0% formaldehyde, 1.25% glutaraldehyde, 0.01 M CaCl2, 4% sucrose and 0.075% ruthenium red, 0.075% L-lysine acetate | 1% osmium tetroxide; 0.075% ruthenium red | Epon-Araldite |
| Procedure 6 (Cryo-ultramicrotomy) | Cacodylate + 4% sucrose | 4.0% formaldehyde, 1.25% glutaraldehyde, 0.01 M CaCl2, 4% sucrose and 0.075% ruthenium red, 0.075% L-lysine acetate | N/A | N/A |
| Treatment | Cryo-Ultramicrotomy | Traditional TEM |
|---|---|---|
| No treatment | clear delineation of outer and inner membrane, cell wall and peptidoglycan layer | poor delineation of membrane structure |
| NCL195 (2 µg/mL) | same as a control | same as a control |
| Colistin (0.125 µg/mL) | mesosome-like structures; swollen membranes | same as a control |
| Colistin (0.25 µg/mL) | mesosome-like structures; more swollen membranes | presence of tubular appendages |
| NCL195/colistin (2/0.25 µg/mL) | increased morphological damage; coronate tubular appendages; mesosome-like structures | increased morphological damage; coronate tubular appendages; swollen and detached membrane |
| Treatment | Cryo-Ultramicrotomy | Traditional TEM |
|---|---|---|
| No treatment | clear delineation of outer and inner membrane, cell wall and peptidoglycan layer | poor delineation of membrane structure |
| NCL195 (2 µg/mL) | same as control | slightly wavy membrane morphology |
| Colistin (1 µg/mL) | clearly visualized membrane damage; presence of tubular appendages | ruffling of the cells; wavy cell membrane structure; presence of tubular appendages |
| NCL195/colistin (2/1 µg/mL) | clearer increased morphological changes; broken outer and inner membrane; mesosome-like structures | increased ultrastructural damage; damaged outer and inner membrane |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, H.T.; O’Donovan, L.A.; Venter, H.; Russell, C.C.; McCluskey, A.; Page, S.W.; Trott, D.J.; Ogunniyi, A.D. Comparison of Two Transmission Electron Microscopy Methods to Visualize Drug-Induced Alterations of Gram-Negative Bacterial Morphology. Antibiotics 2021, 10, 307. https://doi.org/10.3390/antibiotics10030307
Nguyen HT, O’Donovan LA, Venter H, Russell CC, McCluskey A, Page SW, Trott DJ, Ogunniyi AD. Comparison of Two Transmission Electron Microscopy Methods to Visualize Drug-Induced Alterations of Gram-Negative Bacterial Morphology. Antibiotics. 2021; 10(3):307. https://doi.org/10.3390/antibiotics10030307
Chicago/Turabian StyleNguyen, Hang Thi, Lisa A. O’Donovan, Henrietta Venter, Cecilia C. Russell, Adam McCluskey, Stephen W. Page, Darren J. Trott, and Abiodun D. Ogunniyi. 2021. "Comparison of Two Transmission Electron Microscopy Methods to Visualize Drug-Induced Alterations of Gram-Negative Bacterial Morphology" Antibiotics 10, no. 3: 307. https://doi.org/10.3390/antibiotics10030307
APA StyleNguyen, H. T., O’Donovan, L. A., Venter, H., Russell, C. C., McCluskey, A., Page, S. W., Trott, D. J., & Ogunniyi, A. D. (2021). Comparison of Two Transmission Electron Microscopy Methods to Visualize Drug-Induced Alterations of Gram-Negative Bacterial Morphology. Antibiotics, 10(3), 307. https://doi.org/10.3390/antibiotics10030307