
Influence of Nisin-Biogel at Subinhibitory Concentrations on Virulence Expression in Staphylococcus aureus Isolates from Diabetic Foot Infections




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
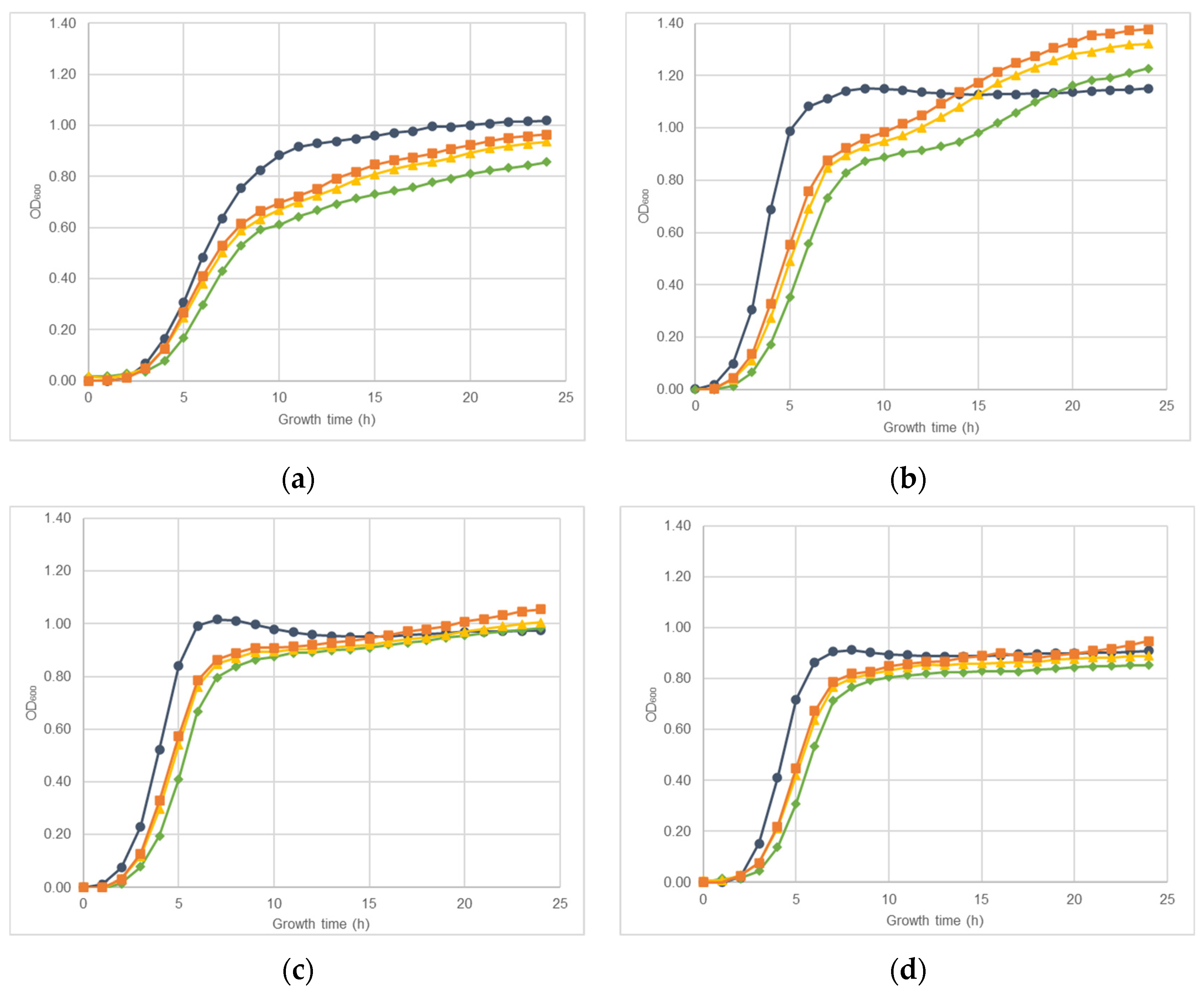
2.1. Effect of Nisin-Biogel Sub-MICs on S. aureus DFI Isolates Growth Rate
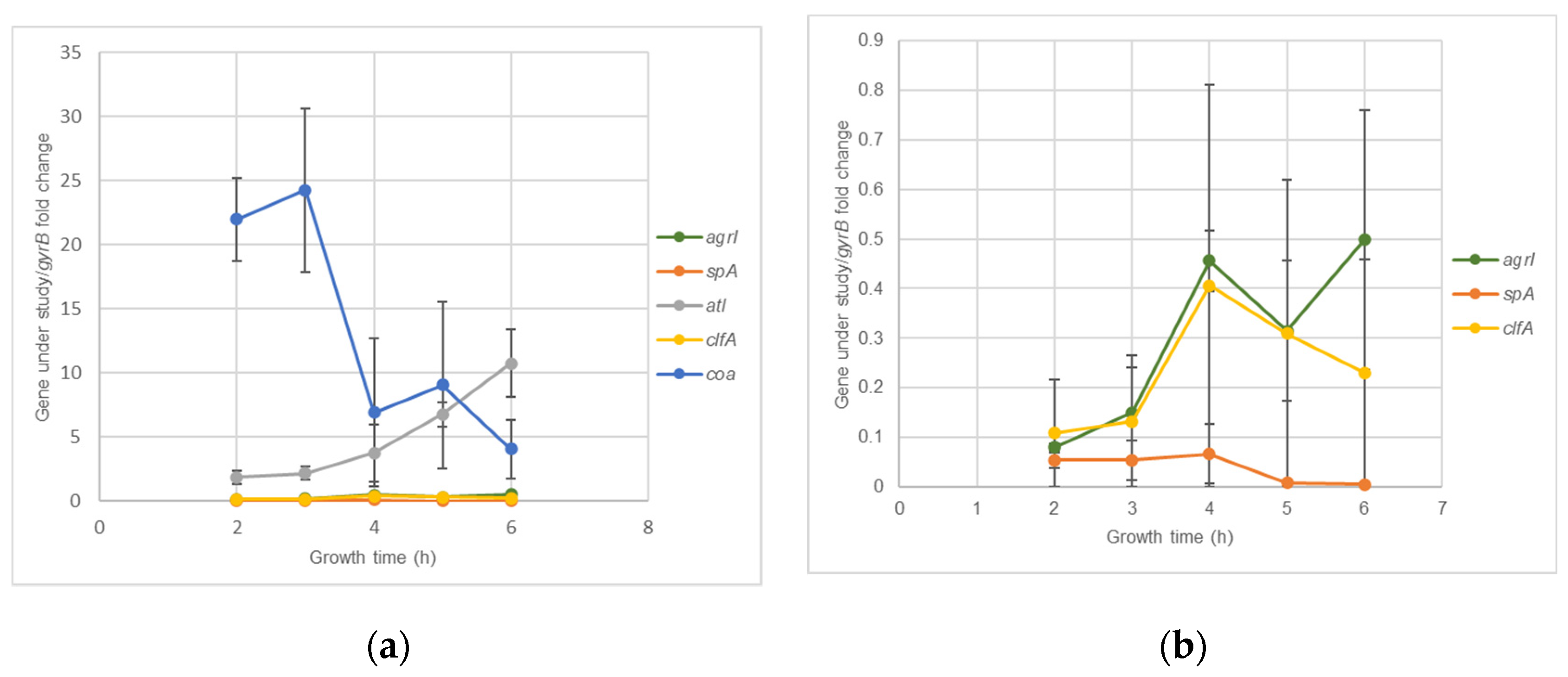
2.2. S. aureus DFI Isolates Gene Expression Kinetics
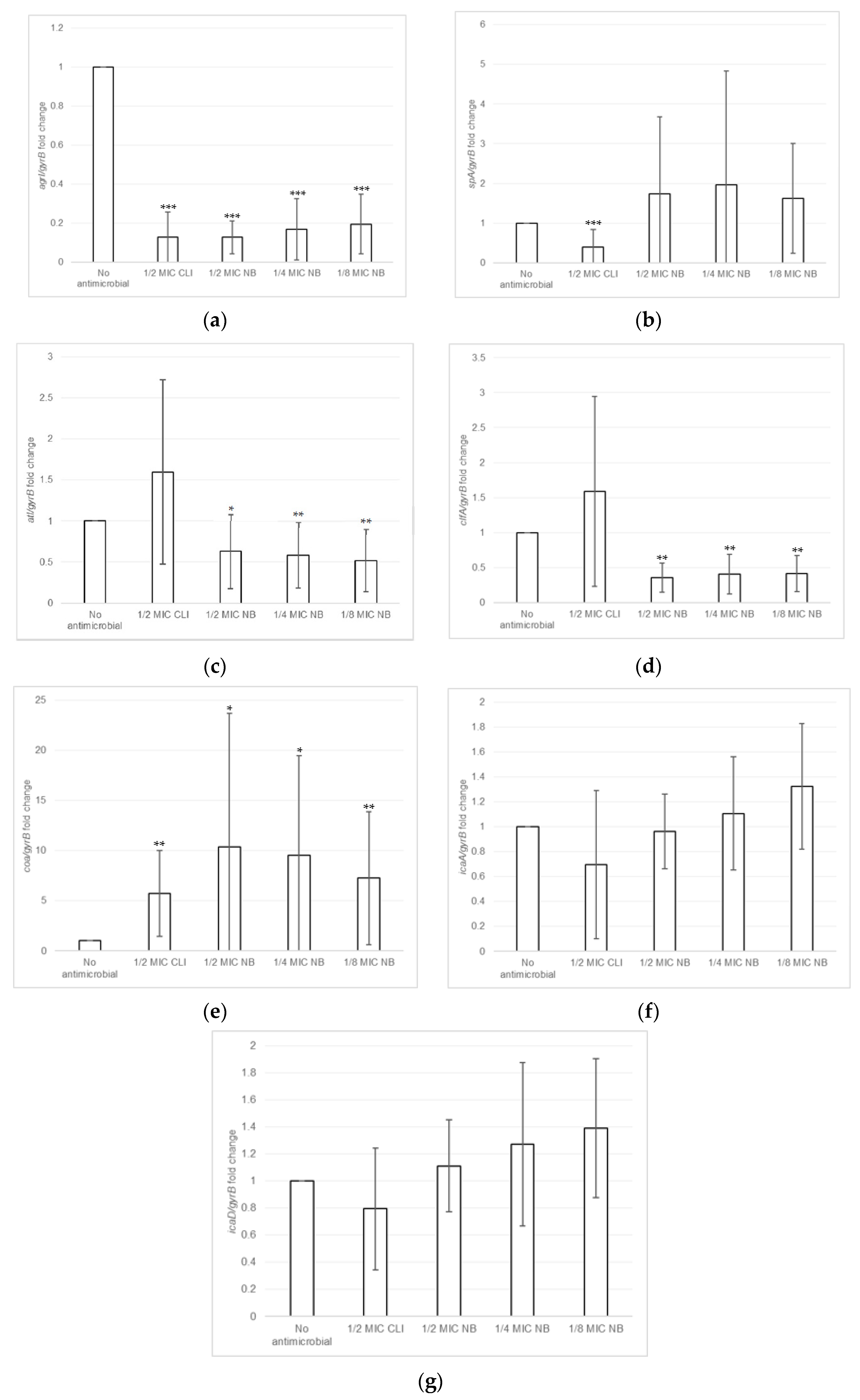
2.3. Effect of Nisin-Biogel at Sub-MICs on Gene Expression by S. aureus DFI Isolates
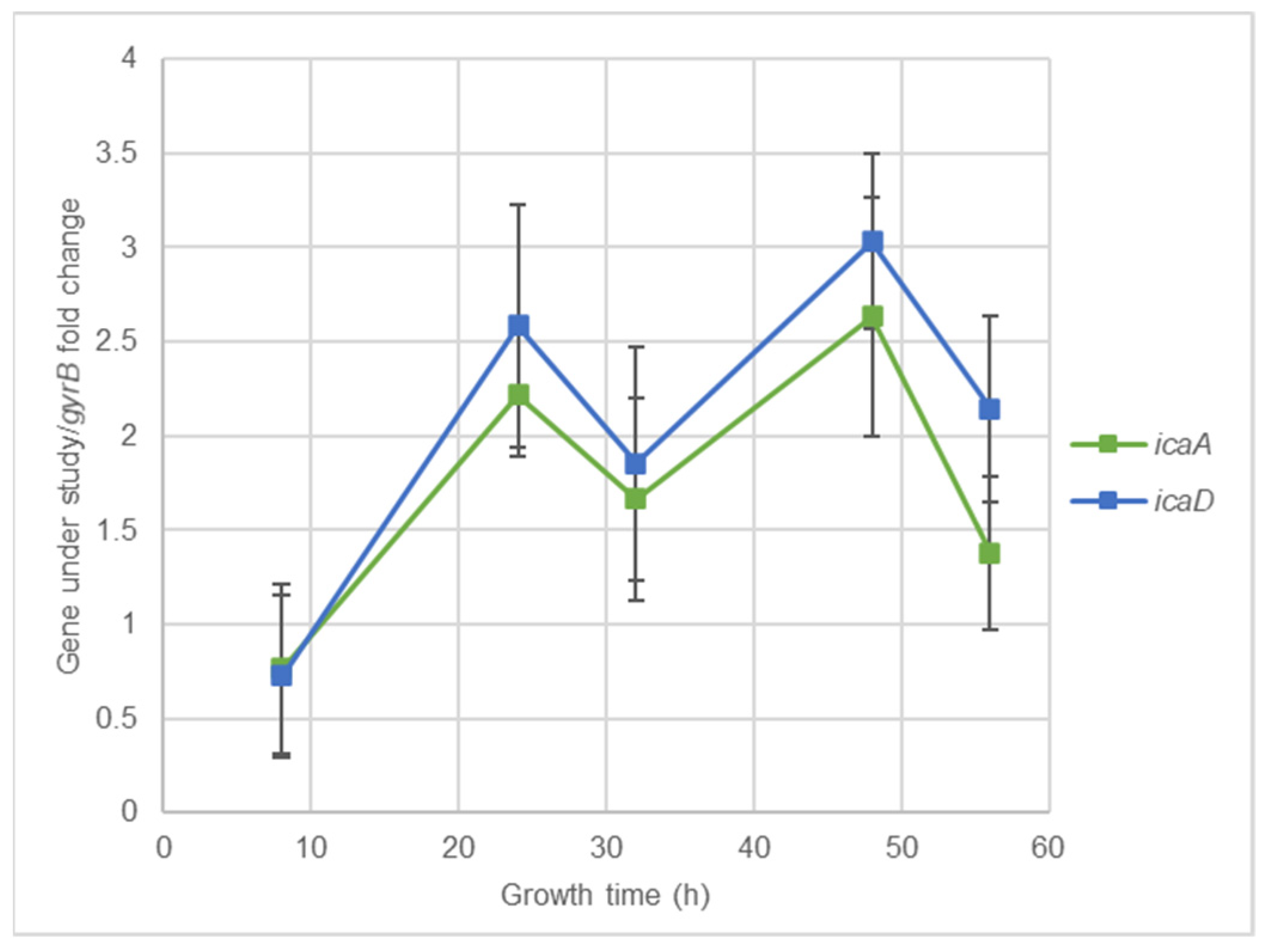
2.4. Effect of Nisin-Biogel Sub-MICs on the Ability of S. aureus DFI Isolates to Form Biofilm
2.5. Effect of Nisin-Biogel Sub-MICs on Coa Production by S. aureus DFI Isolates
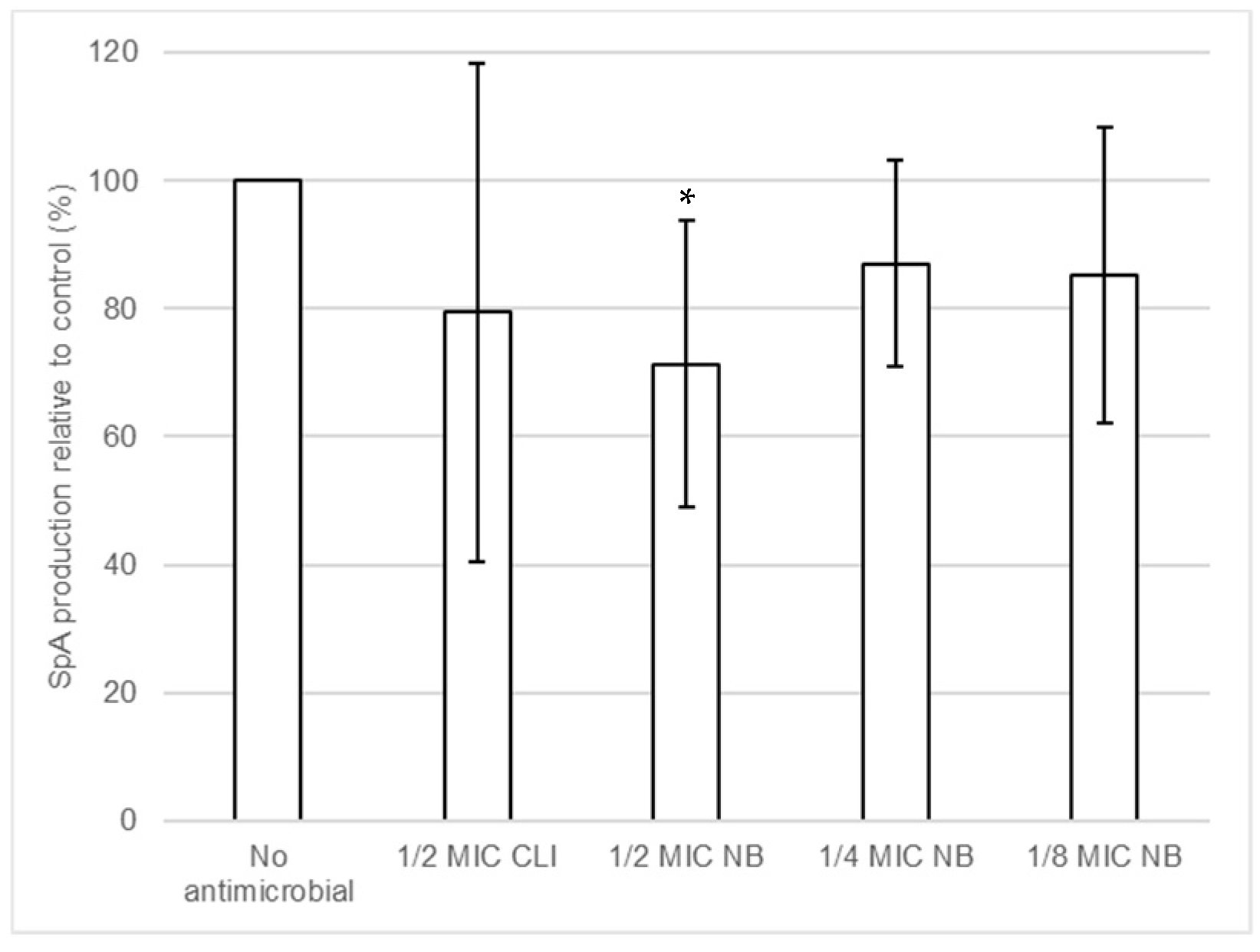
2.6. Effect of Nisin-Biogel Sub-MICs on SpA Release by S. aureus DFI Isolates
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Antimicrobial Solutions
4.3. Effects of Nisin-Biogel Sub-MICs on S. aureus DFI Isolates Growth Rate
4.4. S. aureus DFI Isolates Gene Expression Kinetics
4.5. Effects of Nisin-Biogel Sub-MICs on Gene Expression by S. aureus DFI Isolates
4.6. Effect of Nisin-Biogel Sub-MICs on the Ability of S. aureus DFI Isolates to Form Biofilm
4.7. Effect of Nisin-Biogel Sub-MICs on Coa Production by S. aureus DFI Isolates
4.8. Effect of Nisin-Biogel Sub-MICs on SpA Release by S. aureus DFI Isolates
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IDF Diabetes Atlas—9th Edition. Available online: https://diabetesatlas.org/ (accessed on 27 November 2021).
- Armstrong, D.G.; Boulton, A.J.M.; Bus, S.A. Diabetic Foot Ulcers and Their Recurrence. N. Engl. J. Med. 2017, 376, 2367–2375. [Google Scholar] [CrossRef] [PubMed]
- Noor, S.; Khan, R.U.; Ahmad, J. Understanding Diabetic Foot Infection and Its Management. Diabetes Metab. Syndr. Clin. Res. Rev. 2017, 11, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Muir, T.W. Regulation of Virulence in Staphylococcus aureus: Molecular Mechanisms and Remaining Puzzles. Cell Chem. Biol. 2016, 23, 214–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Periasamy, S.; Joo, H.S.; Duong, A.C.; Bach, T.H.L.; Tan, V.Y.; Chatterjee, S.S.; Cheung, G.Y.C.; Otto, M. How Staphylococcus aureus Biofilms Develop Their Characteristic Structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1281–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, R.; Voggu, L.; Simon, U.K.; Hentschel, P.; Thumm, G.; Götz, F. Activity of the Major Staphylococcal Autolysin Atl. FEMS Microbiol. Lett. 2006, 259, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Porayath, C.; Suresh, M.K.; Biswas, R.; Nair, B.G.; Mishra, N.; Pal, S. Autolysin Mediated Adherence of Staphylococcus aureus with Fibronectin, Gelatin and Heparin. Int. J. Biol. Macromol. 2018, 110, 179–184. [Google Scholar] [CrossRef]
- Singh, V.K. High Level Expression and Purification of Atl, the Major Autolytic Protein of Staphylococcus aureus. Int. J. Microbiol. 2014, 2014, 1–7. [Google Scholar]
- Reffuveille, F.; Josse, J.; Vallé, Q.; Mongaret, C.; Gangloff, S.C. Staphylococcus aureus Biofilms and Their Impact on the Medical Field; InTech: London, UK, 2017; Volume 11, p. 187. [Google Scholar]
- Dickschat, J.S. Quorum Sensing and Bacterial Biofilms. Nat. Prod. Rep. 2010, 27, 343–369. [Google Scholar] [CrossRef] [PubMed]
- Maira-Litrán, T.; Kropec, A.; Abeygunawardana, C.; Joyce, J.; Mark, G.; Goldmann, D.A.; Pier, G.B. Immunochemical Properties of the Staphylococcal Poly-N-Acetylglucosamine Surface Polysaccharide. Infect. Immun. 2002, 70, 4433–4440. [Google Scholar] [CrossRef] [Green Version]
- Otto, M. Staphylococcal Biofilms. In Gram-Positive Pathogens; ASM Press: Washington, DC, USA, 2019; pp. 699–711. [Google Scholar]
- O’Gara, J.P. Ica and beyond: Biofilm Mechanisms and Regulation in Staphylococcus epidermidis and Staphylococcus aureus. FEMS Microbiol. Lett. 2007, 270, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Huntzinger, E.; Boisset, S.; Saveanu, C.; Benito, Y.; Geissmann, T.; Namane, A.; Lina, G.; Etienne, J.; Ehresmann, B.; Ehresmann, C.; et al. Staphylococcus aureus RNAIII and the Endoribonuclease III Coordinately Regulate Spa Gene Expression. EMBO J. 2005, 24, 824–835. [Google Scholar] [CrossRef] [Green Version]
- Kong, K.F.; Vuong, C.; Otto, M. Staphylococcus Quorum Sensing in Biofilm Formation and Infection. Int. J. Med. Microbiol. 2006, 133–139. [Google Scholar] [CrossRef]
- Chastain, C.A.; Klopfenstein, N.; Serezani, C.H.; Aronoff, D.M. A Clinical Review of Diabetic Foot Infections. Clin. Podiatr. Med. Surg. 2019, 36, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Field, D.; O’Connor, R.; Cotter, P.D.; Ross, R.P.; Hill, C. In Vitro Activities of Nisin and Nisin Derivatives Alone and In Combination with Antibiotics against Staphylococcus Biofilms. Front. Microbiol. 2016, 7, 508. [Google Scholar] [CrossRef] [Green Version]
- Grigoropoulou, P.; Eleftheriadou, I.; Jude, E.B.; Tentolouris, N. Diabetic Foot Infections: An Update in Diagnosis and Management. Curr. Diabetes Rep. 2017, 17, 3. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Sherman, S.; Filocamo, S.; Steckl, A.J. Long-Term Antimicrobial Effect of Nisin Released from Electrospun Triaxial Fiber Membranes. Acta Biomater. 2017, 53, 242–249. [Google Scholar] [CrossRef]
- Peterson, E.; Kaur, P. Antibiotic Resistance Mechanisms in Bacteria: Relationships between Resistance Determinants of Antibiotic Producers, Environmental Bacteria, and Clinical Pathogens. Front. Microbiol. 2018, 9, 2928. [Google Scholar] [CrossRef] [PubMed]
- Perez, R.; Perez, M.T.; Elegado, F. Bacteriocins from Lactic Acid Bacteria: A Review of Biosynthesis, Mode of Action, Fermentative Production, Uses, and Prospects. Int. J. Philipp. Sci. Technol. 2015, 8, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Delves-Broughton, J.; Blackburn, P.; Evans, R.J.; Hugenholtz, J. Applications of the Bacteriocin, Nisin. Antonie Van Leeuwenhoek 1996, 69, 193–202. [Google Scholar] [CrossRef]
- Punyauppa-Path, S.; Phumkhachorn, P. Nisin: Production and Mechanism of Antimicrobial Action. Int. J. Curr. Res. Rev. 2015, 7, 47–53. [Google Scholar]
- Mirhosseini, H.; Amid, B.T. A Review Study on Chemical Composition and Molecular Structure of Newly Plant Gum Exudates and Seed Gums. Food Res. Int. 2012, 46, 387–398. [Google Scholar] [CrossRef]
- Narsaiah, K.; Jha, S.N.; Wilson, R.A.; Mandge, H.M.; Manikantan, M.R. Optimizing Microencapsulation of Nisin with Sodium Alginate and Guar Gum. J. Food Sci. Technol. 2014, 51, 4054–4059. [Google Scholar] [CrossRef] [PubMed]
- Prabaharan, M. Prospective of Guar Gum and Its Derivatives as Controlled Drug Delivery Systems. Int. J. Biol. Macromol. 2011, 49, 117–124. [Google Scholar] [CrossRef]
- Santos, R.; Gomes, D.; Macedo, H.; Barros, D.; Tibério, C.; Veiga, A.S.; Tavares, L.; Castanho, M.; Oliveira, M. Guar Gum as a New Antimicrobial Peptide Delivery System against Diabetic Foot Ulcers Staphylococcus aureus Isolates. J. Med. Microbiol. 2016, 65, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Ruza, D.; Cunha, E.; Tavares, L.; Oliveira, M. Diabetic Foot Infections: Application of a Nisin-Biogel to Complement the Activity of Conventional Antibiotics and Antiseptics against Staphylococcus aureus Biofilms. PLoS ONE 2019, 14, e0220000. [Google Scholar] [CrossRef] [Green Version]
- Gomes, D.; Santos, R.; Soares, R.S.; Reis, S.; Carvalho, S.; Rego, P.; Peleteiro, M.C.; Tavares, L.; Oliveira, M. Pexiganan in Combination with Nisin to Control Polymicrobial Diabetic Foot Infections. Antibiotics 2020, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Soares, R.S.; Santos, R.; Cunha, E.; Tavares, L.; Trindade, A.; Oliveira, M. Influence of Storage on the Antimicrobial and Cytotoxic Activities of a Nisin-biogel with Potential to be Applied to Diabetic Foot Infections Treatment. Antibiotics 2020, 9, 781. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.I.; Hughes, D. Microbiological Effects of Sublethal Levels of Antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef]
- Li, J.; Xie, S.; Ahmed, S.; Wang, F.; Gu, Y.; Zhang, C.; Chai, X.; Wu, Y.; Cai, J.; Cheng, G. Antimicrobial Activity and Resistance: Influencing Factors. Front. Pharmacol. 2017, 8, 364. [Google Scholar] [CrossRef] [Green Version]
- Otto, M.P.; Martin, E.; Badiou, C.; Lebrun, S.; Bes, M.; Vandenesch, F.; Etienne, J.; Lina, G.; Dumitrescu, O. Effects of Subinhibitory Concentrations of Antibiotics Factor Expression by Community-Acquired Methicillin-Staphylococcus aureus. J. Antimicrob. Chemother. 2013, 68, 1524–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, J.; Spiegelman, G.B.; Yim, G. The World of Subinhibitory Antibiotic Concentrations. Curr. Opin. Microbiol. 2006, 9, 445–453. [Google Scholar] [CrossRef]
- Herbert, S.; Barry, P.; Novick, R.P. Subinhibitory Clindamycin Differentially Inhibits Transcription of Exoprotein Genes in Staphylococcus aureus. Infect. Immun. 2001, 69, 2996–3003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilcher, K.; Andreoni, F.; Dengler Haunreiter, V.; Seidl, K.; Hasse, B.; Zinkernagel, A.S. Modulation of Staphylococcus aureus Biofilm Matrix by Subinhibitory Concentrations of Clindamycin. Antimicrob. Agents Chemother. 2016, 60, 5957–5967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IDF Europe Members—Portugal. Available online: https://idf.org/our-network/regions-members/europe/members/153-portugal.html (accessed on 27 November 2021).
- Heravi, F.S.; Zakrzewski, M.; Vickery, K.; Armstrong, D.G.; Hu, H. Bacterial Diversity of Diabetic Foot Ulcers: Current Status and Future Prospectives. J. Clin. Med. 2019, 8, 1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeks, B.Y.; Champlin, F.R.; Paulsen, D.B.; Scruggs, D.W.; Lawrence, M.L. Effects of Sub-Minimum Inhibitory Concentration Antibiotic Levels and Temperature on Growth Kinetics and Outer Membrane Protein Expression in Mannheimia haemolytica and Heamophilus somnus. Can. J. Vet. Res. 2005, 69, 1–10. [Google Scholar]
- Zhanel, G.G.; Hoban, D.J.; Harding, G.K. Subinhibitory Antimicrobial Concentrations: A Review of In Vitro and In Vivo Data. Can. J. Infect. Dis. 1992, 3, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Pratten, J.; Foster, S.J.; Chan, P.F.; Wilson, M.; Nair, S.P. Staphylococcus aureus Accessory Regulators: Expression within Biofilms and Effect on Adhesion. Microbes Infect. 2001, 3, 633–637. [Google Scholar] [CrossRef]
- Liu, D.; Li, Z.; Wang, G.; Li, T.; Zhang, L.; Tang, P. Virulence Analysis of Staphylococcus aureus in a Rabbit Model of Infected Full-Thickness Wound under Negative Pressure Wound Therapy. Int. J. Gen. Mol. Microbiol. 2018, 111, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Mottola, C.; Mendes, J.J.; Cristino, J.M.; Cavaco-Silva, P.; Tavares, L.; Oliveira, M. Polymicrobial Biofilms by Diabetic Foot Clinical Isolates. Folia Microbiol. 2016, 61, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.L.; Novick, R.P.; Kreiswirth, B.; Kornblum, J.; Schlievert, P. Cloning, Characterization, and Sequencing of an Accessory Gene Regulator (agr) in Staphylococcus aureus. J. Bacteriol. 1988, 170, 4365–4372. [Google Scholar] [CrossRef] [Green Version]
- Gómez, M.I.; O’Seaghdha, M.; Magargee, M.; Foster, T.J.; Prince, A.S. Staphylococcus aureus Protein A Activates TNFR1 Signaling through Conserved IgG Binding Domains. J. Biol. Chem. 2006, 281, 20190–20196. [Google Scholar] [CrossRef] [Green Version]
- Yanagihara, K.; Tashiro, M.; Fukuda, Y.; Ohno, H.; Higashiyama, Y.; Miyazaki, Y.; Hirakata, Y.; Tomono, K.; Mizuta, Y.; Tsukamoto, K.; et al. Effects of Short Interfering RNA against Methicillin-Resistant Staphylococcus aureus Coagulase In Vitro and In Vivo. J. Antimicrob. Chemother. 2006, 57, 122–126. [Google Scholar] [CrossRef]
- Vandenesch, F.; Kornblum, J.; Novick, R.P. A Temporal Signal, Independent of Agr, Is Required for Hla but Not Spa Transcription in Staphylococcus aureus. J. Bacteriol. 1991, 173, 6313–6320. [Google Scholar] [CrossRef] [Green Version]
- Lebeau, C.; Vandenesch, F.; Greenland, T.; Novick, R.P.; Etienne, J. Coagulase Expression in Staphylococcus aureus Is Positively and Negatively Modulated by an agr-Dependent Mechanism. J. Bacteriol. 1994, 176, 5534–5536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farnsworth, C.W.; Schott, E.M.; Jensen, S.E.; Zukoski, J.; Benvie, A.M.; Refaai, M.A.; Kates, S.L.; Schwarz, E.M.; Zuscik, M.J.; Gill, S.R.; et al. Adaptive Upregulation of Clumping Factor A (clfA) by Staphylococcus aureus in the Obese, Type 2 Diabetic Host Mediates Increased Virulence. Infect. Immun. 2017, 85, e01005-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josefsson, E.; Kubica, M.; Mydel, P.; Potempa, J.; Tarkowski, A. In Vivo Sortase A and Clumping Factor A MRNA Expression during Staphylococcus aureus Infection. Microb. Pathog. 2008, 44, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Pasztor, L.; Ziebandt, A.K.; Nega, M.; Schlag, M.; Haase, S.; Franz-Wachtel, M.; Madlung, J.; Nordheim, A.; Heinrichs, D.E.; Götz, F. Staphylococcal Major Autolysin (Atl) Is Involved in Excretion of Cytoplasmic Proteins. J. Biol. Chem. 2010, 285, 36794–36803. [Google Scholar] [CrossRef] [Green Version]
- Oshida, T.; Takano, M.; Sugai, M.; Suginaka, H.; Matsushita, T. Expression Analysis of the Autolysin Gene (atl) of Staphylococcus aureus. Microbiol. Immunol. 1998, 42, 655–659. [Google Scholar] [CrossRef] [Green Version]
- Atshan, S.S.; Shamsudin, M.N.; Karunanidhi, A.; van Belkum, A.; Lung, L.T.T.; Sekawi, Z.; Nathan, J.J.; Ling, K.H.; Seng, J.S.C.; Ali, A.M.; et al. Quantitative PCR Analysis of Genes Expressed during Biofilm Development of Methicillin Resistant Staphylococcus aureus (MRSA). Infect. Genet. Evol. 2013, 18, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.D.; Colton, E.; Ebert, M.; Anderson, J.M. Gene Expression during S. epidermidis Biofilm Formation on Biomaterials. J. Biomed. Mater. Res. 2012, 100, 2863–2869. [Google Scholar] [CrossRef]
- Smieja, M. Current Indications for the Use of Clindamycin: A Critical Review. Can. J. Infect. Dis. 1998, 9, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Meng, R.; Shi, C.; Liu, Z.; Huang, Y.; Zhao, Z.; Guo, N.; Yu, L. Analysis of the Gene Expression Profile of Staphylococcus aureus Treated with Nisin. Food Control 2016, 59, 499–506. [Google Scholar] [CrossRef]
- Malone, M.; Bjarnsholt, T.; McBain, A.J.; James, G.A.; Stoodley, P.; Leaper, D.; Tachi, M.; Schultz, G.; Swanson, T.; Wolcott, R.D. The Prevalence of Biofilms in Chronic Wounds: A Systematic Review and Meta-Analysis of Published Data. J. Wound Care 2017, 26, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andre, C.; de Jesus Pimentel-Filho, N.; de Almeida Costa, P.M.; Vanetti, M.C.D. Changes in the Composition and Architecture of Staphylococcal Biofilm by Nisin. Braz. J. Microbiol. 2019, 50, 1083–1090. [Google Scholar] [CrossRef]
- Kaplan, J.B. Antibiotic-Induced Biofilm Formation. Int. J. Artif. Organs 2011, 34, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, A.; Field, D.; Pérez-Ibarreche, M.; Warda, A.K.; Hill, C.; Ross, R.P. Vancomycin and Nisin A Are Effective against Biofilms of Multi-Drug Resistant Staphylococcus aureus Isolates from Human Milk. PLoS ONE 2020, 15, e0233284. [Google Scholar] [CrossRef]
- Majidpour, A.; Fathizadeh, S.; Afshar, M.; Rahbar, M.; Boustanshenas, M.; Heidarzadeh, M.; Arbabi, L.; Soleymanzadeh Moghadam, S. Dose-Dependent Effects of Common Antibiotics Used to Treat Staphylococcus aureus on Biofilm Formation. Iran. J. Pathol. 2017, 12, 362–370. [Google Scholar] [CrossRef]
- Hodille, E.; Rose, W.; Diep, B.A.; Goutelle, S.; Lina, G.; Dumitrescu, O. The Role of Antibiotics in Modulating Virulence in Staphylococcus aureus. Clin. Microbiol. Rev. 2017, 30, 887–917. [Google Scholar] [CrossRef] [Green Version]
- McAdow, M.; Missiakas, D.M.; Schneewind, O. Staphylococcus aureus Secretes Coagulase and von Willebrand Factor Binding Protein to Modify the Coagulation Casca de and Establish Host Infections. J. Innate Immun. 2012, 4, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Blickwede, M.; Wolz, C.; Valentin-Weigand, P.; Schwarz, S. Influence of Clindamycin on the Stability of Coa and FnbB Transcripts and Adherence Properties of Staphylococcus aureus Newman. FEMS Microbiol. Lett. 2005, 252, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Mendes, J.J.; Marques-Costa, A.; Vilela, C.; Neves, J.; Candeias, N.; Cavaco-Silva, P.; Melo-Cristino, J. Clinical and Bacteriological Survey of Diabetic Foot Infections in Lisbon. Diabetes Res. Clin. Pract. 2012, 95, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Mottola, C.; Semedo-Lemsaddek, T.; Mendes, J.J.; Melo-Cristino, J.; Tavares, L.; Cavaco-Silva, P.; Oliveira, M. Molecular Typing, Virulence Traits and Antimicrobial Resistance of Diabetic Foot Staphylococci. J. Biomed. Sci. 2016, 23, 33. [Google Scholar] [CrossRef] [Green Version]
- Mottola, C.; Matias, C.S.; Mendes, J.J.; Melo-Cristino, J.; Tavares, L.; Cavaco-Silva, P.; Oliveira, M. Susceptibility Patterns of Staphylococcus aureus Biofilms in Diabetic Foot Infections. BMC Microbiol. 2016, 16, 119. [Google Scholar] [CrossRef] [Green Version]
- Kearns, A.M.; Seiders, R.R.; Wheeler, J.; Freeman, R.; Steward, M. Rapid Detection of Methicillin-Resistant Staphylococci by Multiplex PCR. J. Hosp. Infect. 1999, 43, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Sperber, W.H.; Tatini, S.R. Interpretation of the Tube Coagulase Test for Identification of Staphylococcus aureus. Appl. Microbiol. 1975, 29, 502–505. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jesus, C.; Soares, R.; Cunha, E.; Grilo, M.; Tavares, L.; Oliveira, M. Influence of Nisin-Biogel at Subinhibitory Concentrations on Virulence Expression in Staphylococcus aureus Isolates from Diabetic Foot Infections. Antibiotics 2021, 10, 1501. https://doi.org/10.3390/antibiotics10121501
Jesus C, Soares R, Cunha E, Grilo M, Tavares L, Oliveira M. Influence of Nisin-Biogel at Subinhibitory Concentrations on Virulence Expression in Staphylococcus aureus Isolates from Diabetic Foot Infections. Antibiotics. 2021; 10(12):1501. https://doi.org/10.3390/antibiotics10121501
Chicago/Turabian StyleJesus, Carolina, Rui Soares, Eva Cunha, Miguel Grilo, Luís Tavares, and Manuela Oliveira. 2021. "Influence of Nisin-Biogel at Subinhibitory Concentrations on Virulence Expression in Staphylococcus aureus Isolates from Diabetic Foot Infections" Antibiotics 10, no. 12: 1501. https://doi.org/10.3390/antibiotics10121501
APA StyleJesus, C., Soares, R., Cunha, E., Grilo, M., Tavares, L., & Oliveira, M. (2021). Influence of Nisin-Biogel at Subinhibitory Concentrations on Virulence Expression in Staphylococcus aureus Isolates from Diabetic Foot Infections. Antibiotics, 10(12), 1501. https://doi.org/10.3390/antibiotics10121501