
Alternating Current-Dielectrophoresis Collection and Chaining of Phytoplankton on Chip: Comparison of Individual Species and Artificial Communities


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Test Medium
2.2. DEP Experimental Setup and Parameter Optimization
2.3. DEP Behavior of Representative Phytoplankton Species
2.4. Effect of AC Field on Membrane Permeability of the Cells
2.5. Microscopy and Image Analysis
2.6. Cell Chaining Efficiency Determination
2.7. Measurements of Cell Size and Zeta Potential
2.8. Modelling of Chaining Efficiency
2.9. Statistical Analysis
3. Results
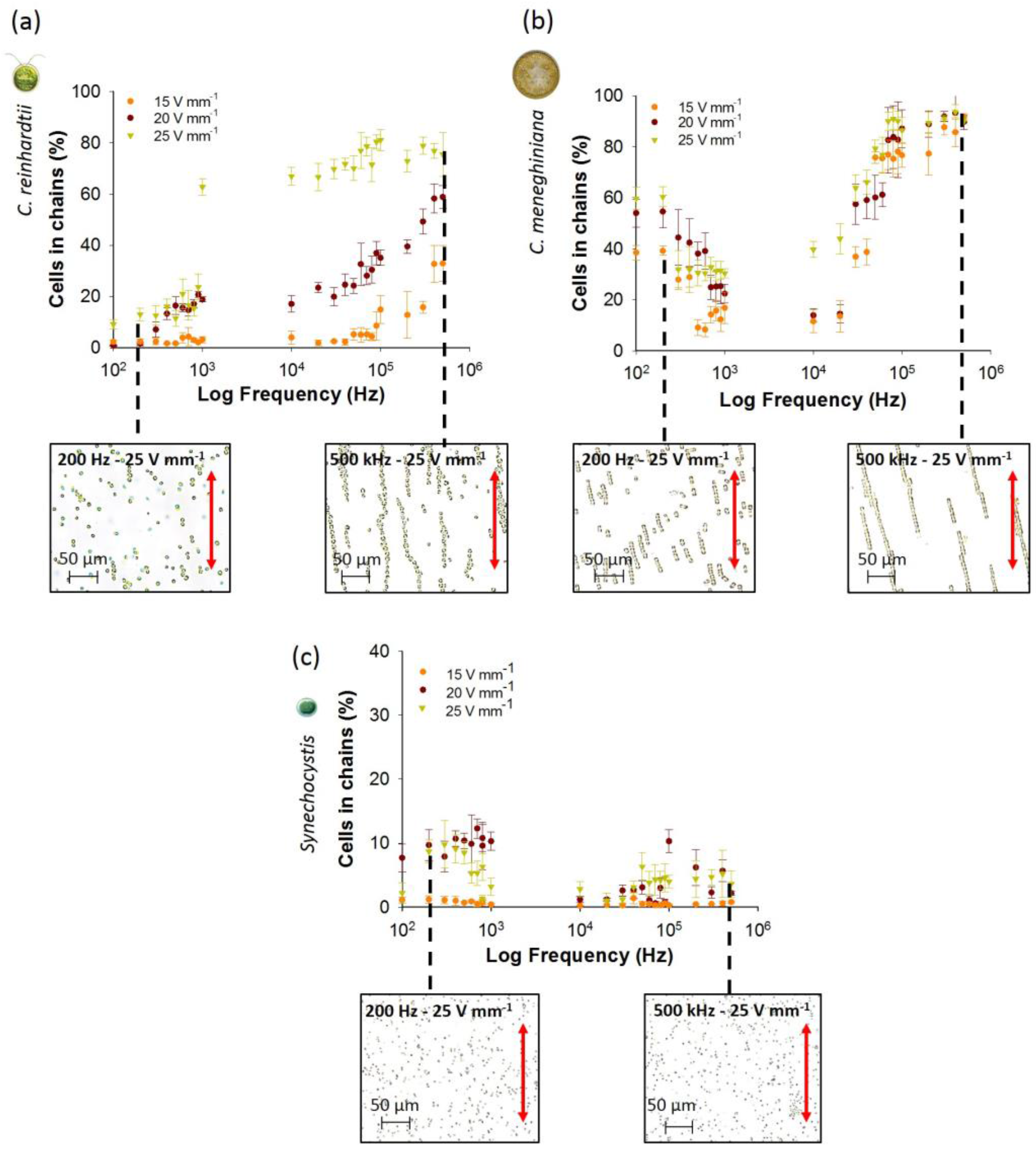
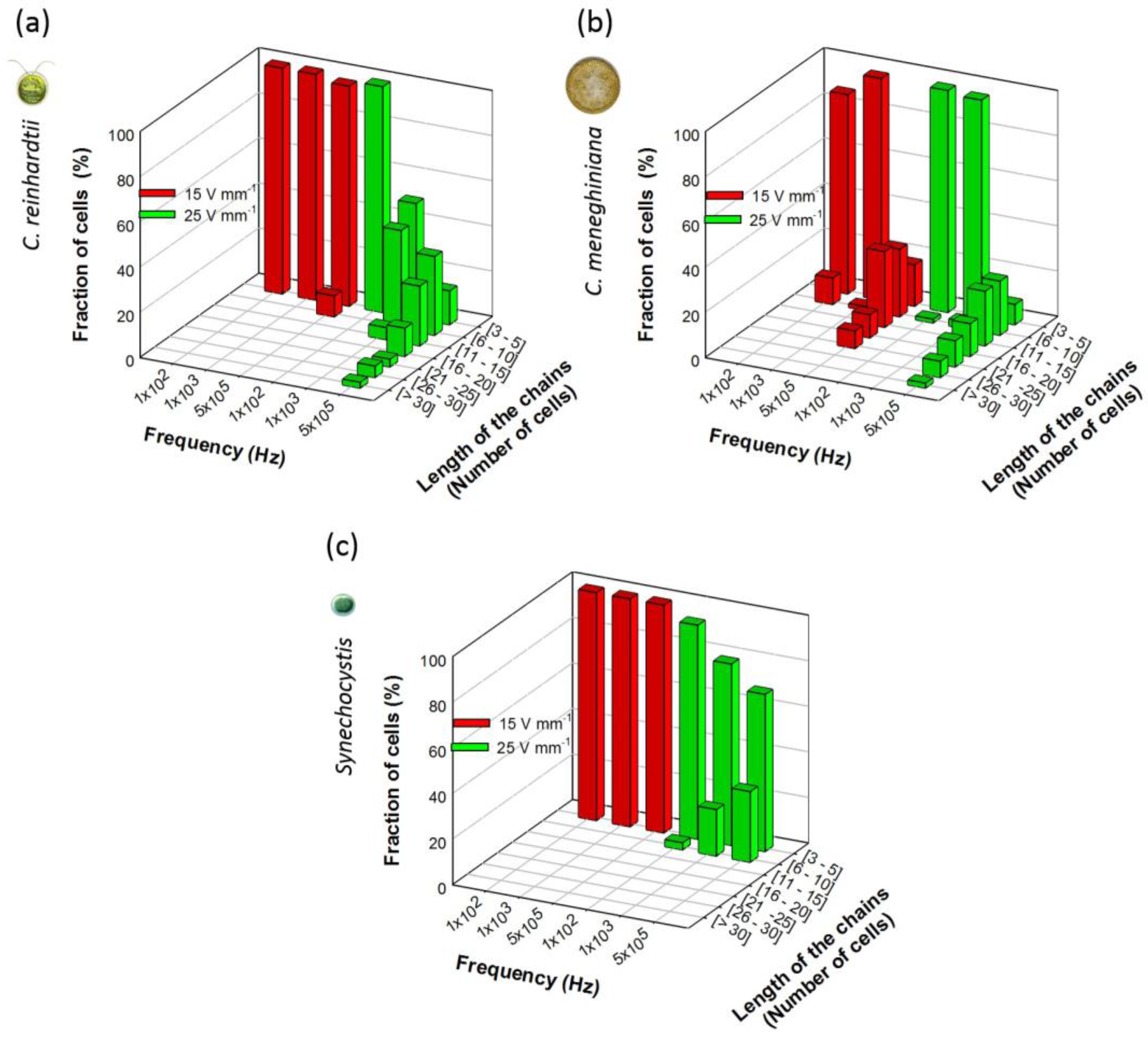
3.1. Collection and “Pearl” Chain Formation by the Individual Microorganisms
3.1.1. Effect of AC Field Frequency and Intensity on Chaining Efficiency and Chain Length
3.1.2. Effect of Cell Concentrations on Chaining Efficiency
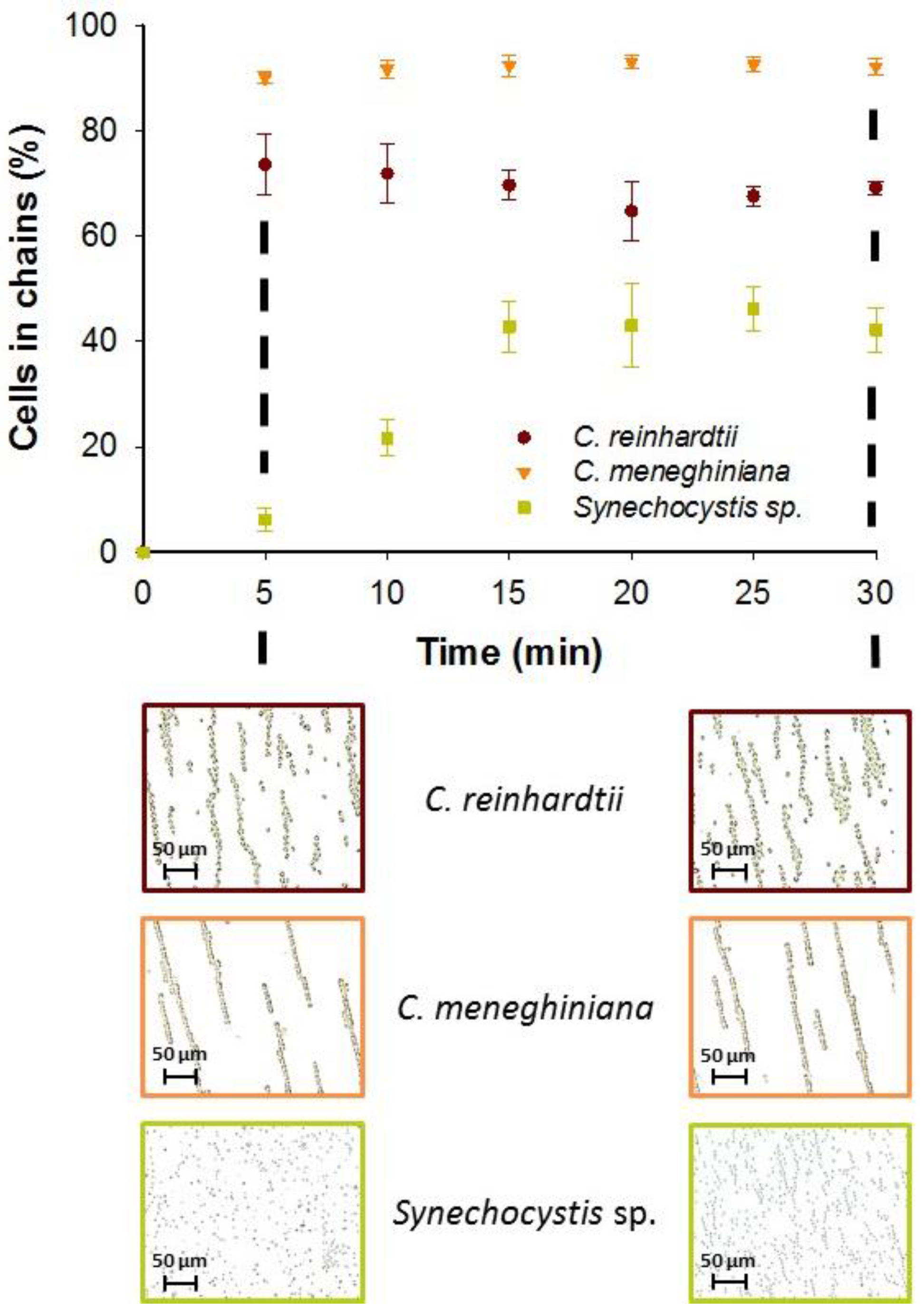
3.1.3. Effect of AC Field Duration on Chaining Efficiency
3.1.4. Effect of the AC Field on the Cell Viability
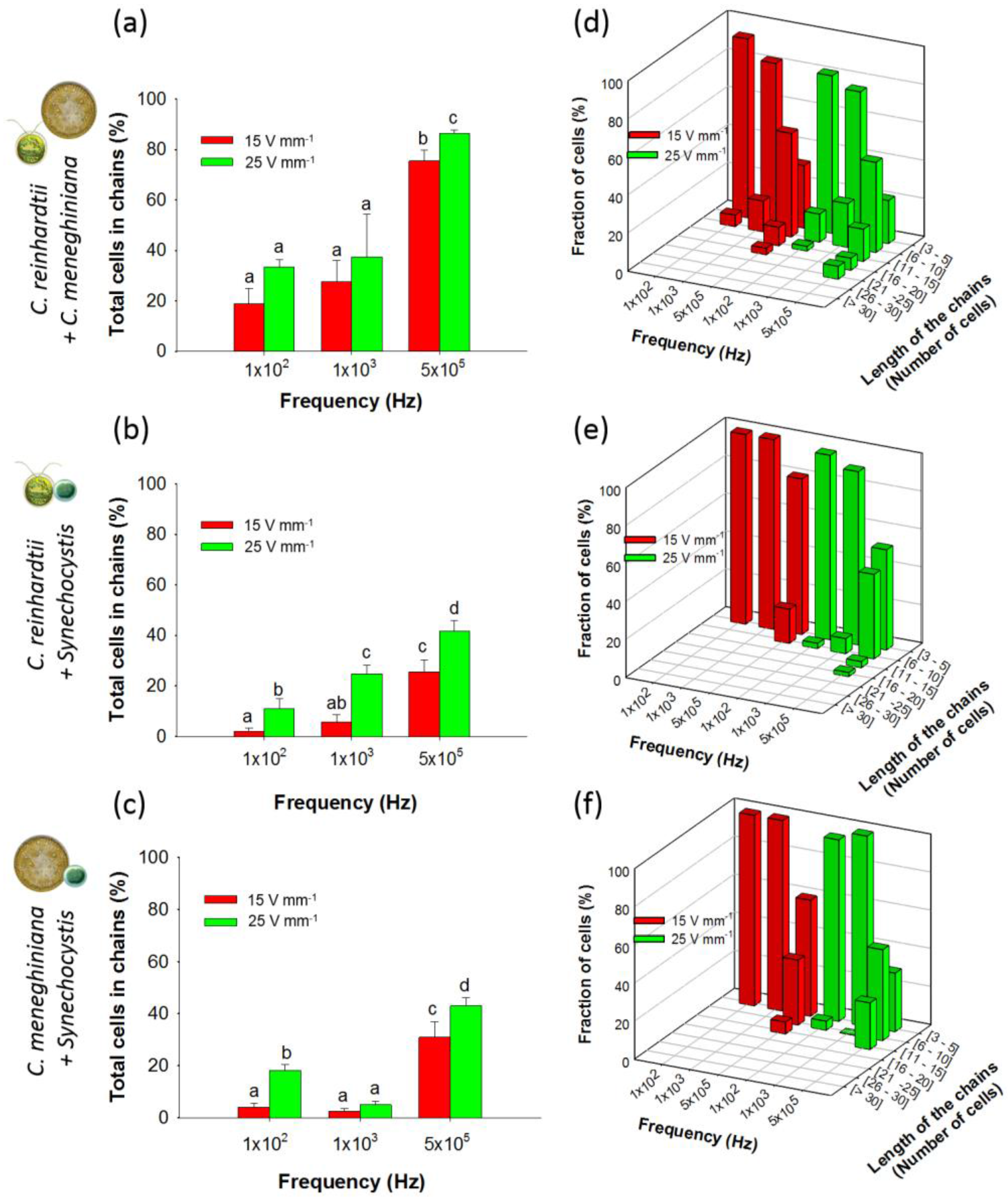
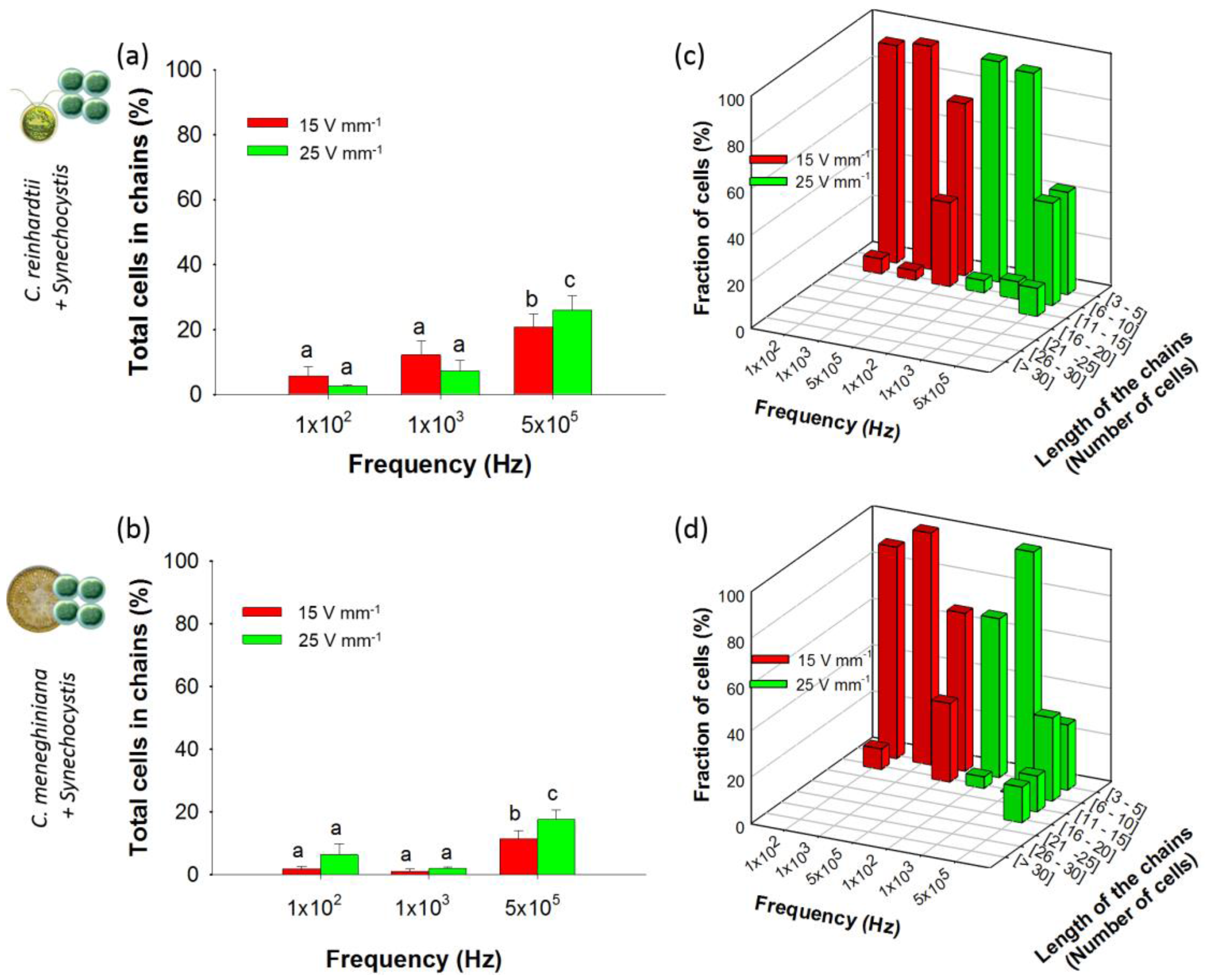
3.2. Collection and “Pearl” Chain Formation by Artificial Communities
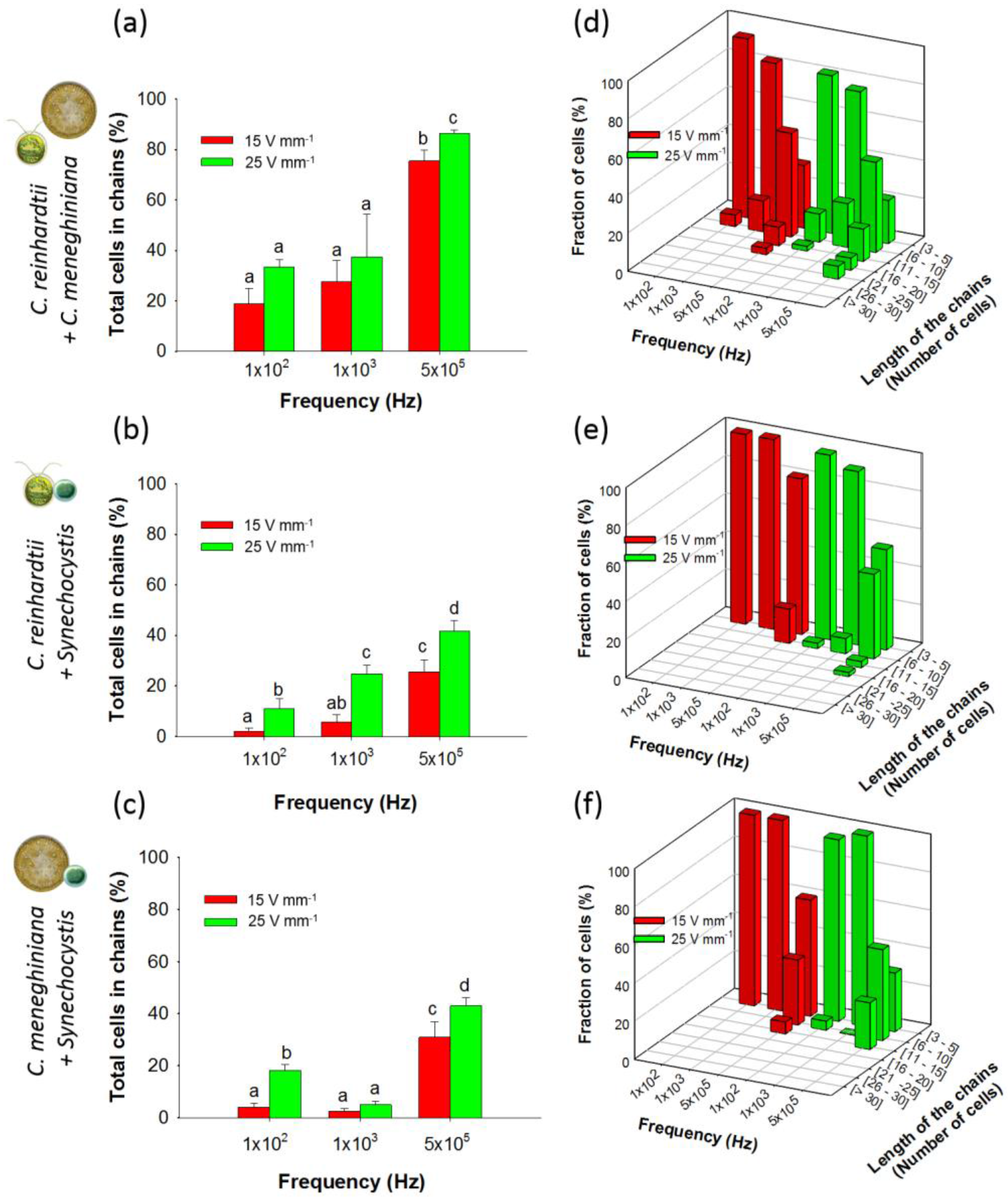
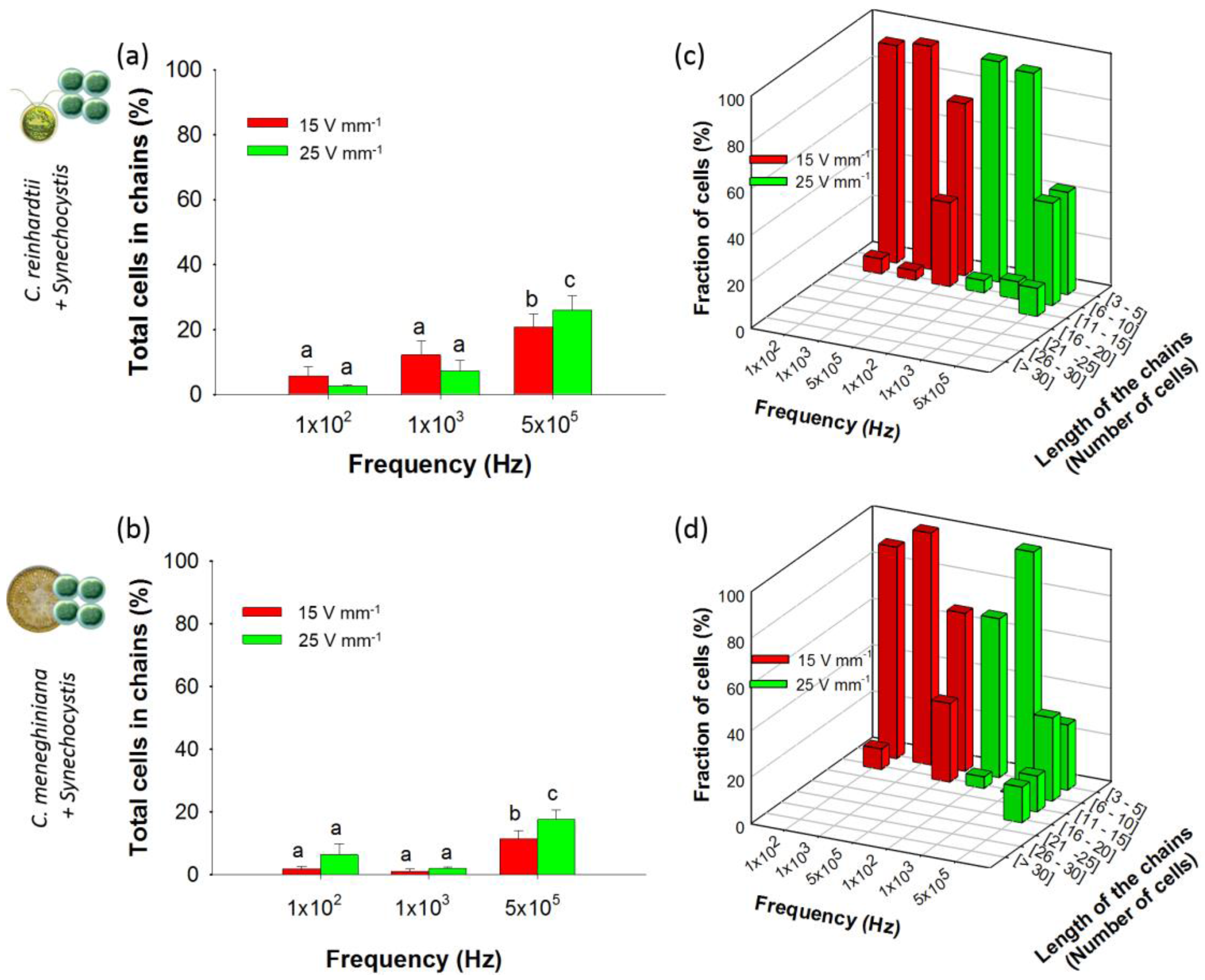
3.2.1. Chaining of Binary Artificial Communities in Equal Concentration
3.2.2. Chaining of Binary Artificial Communities Containing an Excess of Cyanobacteria
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Suthers, I.M.; Rissik, D. Plankton: A Guide to Their Ecology and Monitoring for Water Quality; CSIRO Publishing: Collingwood, Australia, 2008. [Google Scholar]
- Willén, E. Phytoplankton in water quality assessment—An indicator concept. In Water Quality Measurements Series; John Wiley & Sons: Hoboken, NJ, USA, 2007; pp. 56–80. [Google Scholar]
- Thakur, R.K.; Jindal, R.; Singh, U.B.; Ahluwalia, A.S. Plankton diversity and water quality assessment of three freshwater lakes of mandi (Himachal Pradesh, India) with special reference to planktonic indicators. Environ. Monit. Assess. 2013, 185, 8355–8373. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Gopal, K. Biomonitoring of Water and Waste Water; Springer: Heidelberg, Germany, 2013. [Google Scholar]
- Su, L.A.; Jia, W.Z.; Hou, C.J.; Lei, Y. Microbial biosensors: A review. Biosens. Bioelectron. 2011, 26, 1788–1799. [Google Scholar] [CrossRef] [PubMed]
- Brayner, R.; Coute, A.; Livage, J.; Perrette, C.; Sicard, C. Micro-algal biosensors. Anal. Bioanal. Chem. 2011, 401, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Pasco, N.F.; Weld, R.J.; Hay, J.M.; Gooneratne, R. Development and applications of whole cell biosensors for ecotoxicity testing. Anal. Bioanal. Chem. 2011, 400, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Pethig, R. Review article-dielectrophoresis: Status of the theory, technology, and applications. Biomicrofluidics 2010, 4, 022811. [Google Scholar] [CrossRef] [PubMed]
- Khoshmanesh, K.; Nahavandi, S.; Baratchi, S.; Mitchell, A.; Kalantar-Zadeh, K. Dielectrophoretic platforms for bio-microfluidic systems. Biosens. Bioelectron. 2011, 26, 1800–1814. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.; Morgan, H.; Green, N.G.; Castellanos, A. Ac electrokinetics: A review of forces in microelectrode structures. J. Phys. D Appl. Phys. 1998, 31, 2338–2353. [Google Scholar] [CrossRef]
- Jesus-Perez, N.M.; Lapizco-Encinas, B.H. Dielectrophoretic monitoring of microorganisms in environmental applications. Electrophoresis 2011, 32, 2331–2357. [Google Scholar] [CrossRef] [PubMed]
- Morgan, H.; Hughes, M.P.; Green, N.G. Separation of submicron bioparticles by dielectrophoresis. Biophys. J. 1999, 77, 516–525. [Google Scholar] [CrossRef]
- Suehiro, J.; Hamada, R.; Noutomi, D.; Shutou, M.; Hara, M. Selective detection of viable bacteria using dielectrophoretic impedance measurement method. J. Electrost. 2003, 57, 157–168. [Google Scholar] [CrossRef]
- Zhu, K.; Kaprelyants, A.S.; Salina, E.G.; Markx, G.H. Separation by dielectrophoresis of dormant and nondormant bacterial cells of mycobacterium smegmatis. Biomicrofluidics 2010, 4, 022809. [Google Scholar] [CrossRef] [PubMed]
- Allahrabbi, N.; Chia, Y.S.; Saifullah, M.S.; Lim, K.M.; Yung, L.Y. A hybrid dielectrophoretic system for trapping of microorganisms from water. Biomicrofluidics 2015, 9, 034110. [Google Scholar] [CrossRef] [PubMed]
- Suscillon, C.; Velev, O.D.; Slaveykova, V.I. Alternating current-dielectrophoresis driven on-chip collection and chaining of green microalgae in freshwaters. Biomicrofluidics 2013, 7, 24109. [Google Scholar] [CrossRef] [PubMed]
- Siebman, C.; Velev, O.D.; Slaveykova, V.I. Two-dimensional algal collection and assembly by combining ac-dielectrophoresis with fluorescence detection for contaminant-induced oxidative stress sensing. Biosensors 2015, 5, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Velev, O.D.; Bhatt, K.H. On-chip micromanipulation and assembly of colloidal particles by electric fields. Soft Matter 2006, 2, 738–750. [Google Scholar] [CrossRef]
- Gupta, S.; Alargova, R.G.; Kilpatrick, P.K.; Velev, O.D. On-chip dielectrophoretic coassembly of live cells and particles into responsive biomaterials. Langmuir 2010, 26, 3441–3452. [Google Scholar] [CrossRef] [PubMed]
- Lumsdon, S.O.; Kaler, E.W.; Velev, O.D. Two-dimensional crystallization of microspheres by a coplanar ac electric field. Langmuir 2004, 20, 2108–2116. [Google Scholar] [CrossRef] [PubMed]
- Kadaksham, J.; Singh, P.; Aubry, N. Dielectrophoresis induced clustering regimes of viable yeast cells. Electrophoresis 2005, 26, 3738–3744. [Google Scholar] [CrossRef] [PubMed]
- Giner, V.; Sancho, M.; Lee, R.S.; Martinez, G.; Pethig, R. Transverse dipolar chaining in binary suspensions induced by rf fields. J. Phys. D Appl. Phys. 1999, 32, 1182–1186. [Google Scholar] [CrossRef]
- Pethig, R.; Markx, G.H. Applications of dielectrophoresis in biotechnology. Trends Biotechnol. 1997, 15, 426–432. [Google Scholar] [CrossRef]
- Gupta, S.; Alargova, R.G.; Kilpatrick, P.K.; Velev, O.D. On-chip electric field driven assembly of biocomposites from live cells and functionalized particles. Soft Matter 2008, 4, 726–730. [Google Scholar] [CrossRef]
- Flores-Rodriguez, N.; Markx, G.H. Improved levitation and trapping of particles by negative dielectrophoresis by the addition of amphoteric molecules. J. Phys. D Appl. Phys. 2004, 37, 353–361. [Google Scholar] [CrossRef]
- Khoshmanesh, K.; Baratchi, S.; Tovar-Lopez, F.J.; Nahavandi, S.; Wlodkowic, D.; Mitchell, A.; Kalantar-Zadeh, K. On-chip separation of lactobacillus bacteria from yeasts using dielectrophoresis. Microfluid. Nanofluid. 2012, 12, 597–606. [Google Scholar]
- Markx, G.H.; Huang, Y.; Zhou, X.F.; Pethig, R. Dielectrophoretic characterization and separation of microorganisms. Microbiology 1994, 140, 585–591. [Google Scholar] [CrossRef]
- Yang, L.; Banada, P.P.; Bhunia, A.K.; Bashir, R. Effects of dielectrophoresis on growth, viability and immuno-reactivity of listeria monocytogenes. J. Biol. Eng. 2008, 2, 1. [Google Scholar] [CrossRef]
- Bhatt, K.H.; Velev, O.D. Control and modeling of the dielectrophoretic assembly of on-chip nanoparticle wires. Langmuir 2004, 20, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Wanichapichart, P.; Bunthawin, S.; Kaewpaiboon, A.; Kanchanapoom, K. Determination of cell dielectric properties using dielectrophoretic technique. ScienceAsia 2002, 28, 113–119. [Google Scholar] [CrossRef]
- Park, S.; Zhang, Y.; Wang, T.H.; Yang, S. Continuous dielectrophoretic bacterial separation and concentration from physiological media of high conductivity. Lab Chip 2011, 11, 2893–2900. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.H.; Jones Stephen, H. The General Properties of Si, Ge, SiGe, SiO2 and Si3N4; Virginia Semiconductor: Fredericksburg, VA, USA, 2002. [Google Scholar]
- Kroger, N.; Poulsen, N. Diatoms-from cell wall biogenesis to nanotechnology. Annu. Rev. Genet. 2008, 42, 83–107. [Google Scholar] [CrossRef] [PubMed]
- Melvin, E.M.; Moore, B.R.; Gilchrist, K.H.; Grego, S.; Velev, O.D. On-chip collection of particles and cells by ac electroosmotic pumping and dielectrophoresis using asymmetric microelectrodes. Biomicrofluidics 2011, 5, 034113. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, K.H.; Grego, S.; Velev, O.D. An ac electrokinetic technique for collection and concentration of particles and cells on patterned electrodes. Langmuir 2005, 21, 6603–6612. [Google Scholar] [CrossRef] [PubMed]
- Ermolina, I.; Morgan, H. The electrokinetic properties of latex particles: Comparison of electrophoresis and dielectrophoresis. J. Colloid Interface Sci. 2005, 285, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Unni, H.N.; Hartono, D.; Yung, L.Y.L.; Ng, M.M.L.; Lee, H.P.; Khoo, B.C.; Lim, K.M. Characterization and separation of cryptosporidium and giardia cells using on-chip dielectrophoresis. Biomicrofluidics 2012, 6, 012805. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Morales, F.H.; Duarte, J.E.; Samitier-Marti, J. Bacterial handling under the influence of non-uniform electric fields: Dielectrophoretic and electrohydrodynamic effects. An. Acad. Bras. Cienc. 2008, 80, 627–638. [Google Scholar] [CrossRef]
- Green, N.G.; Morgan, H. Dielectrophoresis of submicrometer latex spheres. 1. Experimental results. J. Phys. Chem. B 1999, 103, 41–50. [Google Scholar] [CrossRef]
- Green, N.G.; Ramos, A.; Morgan, H.; Castellanos, A. Sub-micrometre ac electrokinetics: Particle dynamics under the influence of dielectrophoresis and electrohydrodynamics. Inst. Phys. Conf. Ser. 1999, 163, 89–92. [Google Scholar]
- Donato, S.S.; Chu, V.; Prazeres, D.M.; Conde, J.P. Metabolic viability of Echerichia coli trapped by dielectrophoresis in microfluidics. Electrophoresis 2013, 34, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Bharti, B.; Findenegg, G.H.; Velev, O.D. Co-assembly of oppositely charged particles into linear clusters and chains of controllable length. Sci. Rep. 2012, 2, 1004. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Value | Units | |||
|---|---|---|---|---|---|
| C. reinhardtii | Synechocystis sp. | C. meneghiniana | |||
| Relative cytoplasm dielectric constant | Ɛ2 | 1.33 × 10−9 a | 5.40 × 10−10 b | 1.33 × 10−9 a | C2·J−1·m−1 |
| Cytoplasm conductivity | σ2 | 8.00 × 10−3 a | 1.90 × 10−1 b | 8.00 × 10−3 a | S·m−1 |
| Membrane capacitance | cm | 1.42 × 10−2 a | 5.45 × 10−2 b | 1.42 × 10−2 a | F·m−2 |
| Relative cell wall dielectric constant | Ɛ1 | 6.20 × 10−10 a | 5.31 × 10−10 b | 3.45 × 10−11 c | C2·J−1·m−1 |
| Cell wall conductivity | σ1 | 5.00 × 10−2 a | 6.80 × 10−1 b | 1.00 × 10−20 c | S·m−1 |
| Inner radius | R | 6.00 × 10−6 a | 1.83 × 10−6 b | 8.36 × 10−6 a | m |
| Outer radius d | R0 | 6.50 × 10−6 | 1.96 × 10−6 | 8.86 × 10−6 | m |
| Zeta potential d | ζ | −2.03 × 10−2 | −1.82 × 10−2 | −1.49 × 10−2 | V |
| Double layer thickness e | Δ | 3.00 × 10−9 | 3.00 × 10−9 | 3.00 × 10−9 | m |
| Debye length e | κ−1 | 9.60 × 10−9 | 9.60 × 10−9 | 9.60 × 10−9 | m |
| Relative medium dielectric constant e | Ɛm | 7.08 × 10−10 | 7.08 × 10−10 | 7.08 × 10−10 | C2·J−1·m−1 |
| Medium conductivity d | σm | 3.20 × 10−1 | 3.20 × 10−1 | 3.20 × 10−1 | S·m−1 |
| Angular frequency e | ω | variable | variable | variable | Rad·s−1 |
| Elementary charge e | e | 1.60 × 10−19 | 1.60 × 10−19 | 1.60 × 10−19 | C |
| Boltzmann constant e | k | 1.38 × 10−23 | 1.38 × 10−23 | 1.38 × 10−23 | J·K−1 |
| Absolute temperature e | T | 298 | 298 | 298 | K |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siebman, C.; Velev, O.D.; Slaveykova, V.I. Alternating Current-Dielectrophoresis Collection and Chaining of Phytoplankton on Chip: Comparison of Individual Species and Artificial Communities. Biosensors 2017, 7, 4. https://doi.org/10.3390/bios7010004
Siebman C, Velev OD, Slaveykova VI. Alternating Current-Dielectrophoresis Collection and Chaining of Phytoplankton on Chip: Comparison of Individual Species and Artificial Communities. Biosensors. 2017; 7(1):4. https://doi.org/10.3390/bios7010004
Chicago/Turabian StyleSiebman, Coralie, Orlin D. Velev, and Vera I. Slaveykova. 2017. "Alternating Current-Dielectrophoresis Collection and Chaining of Phytoplankton on Chip: Comparison of Individual Species and Artificial Communities" Biosensors 7, no. 1: 4. https://doi.org/10.3390/bios7010004
APA StyleSiebman, C., Velev, O. D., & Slaveykova, V. I. (2017). Alternating Current-Dielectrophoresis Collection and Chaining of Phytoplankton on Chip: Comparison of Individual Species and Artificial Communities. Biosensors, 7(1), 4. https://doi.org/10.3390/bios7010004