
Monitoring Intact Viruses Using Aptamers

Abstract
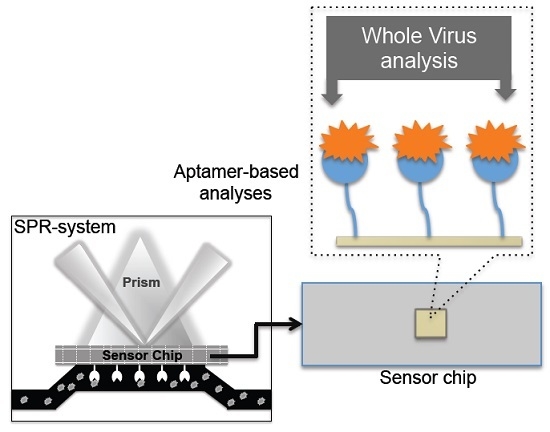
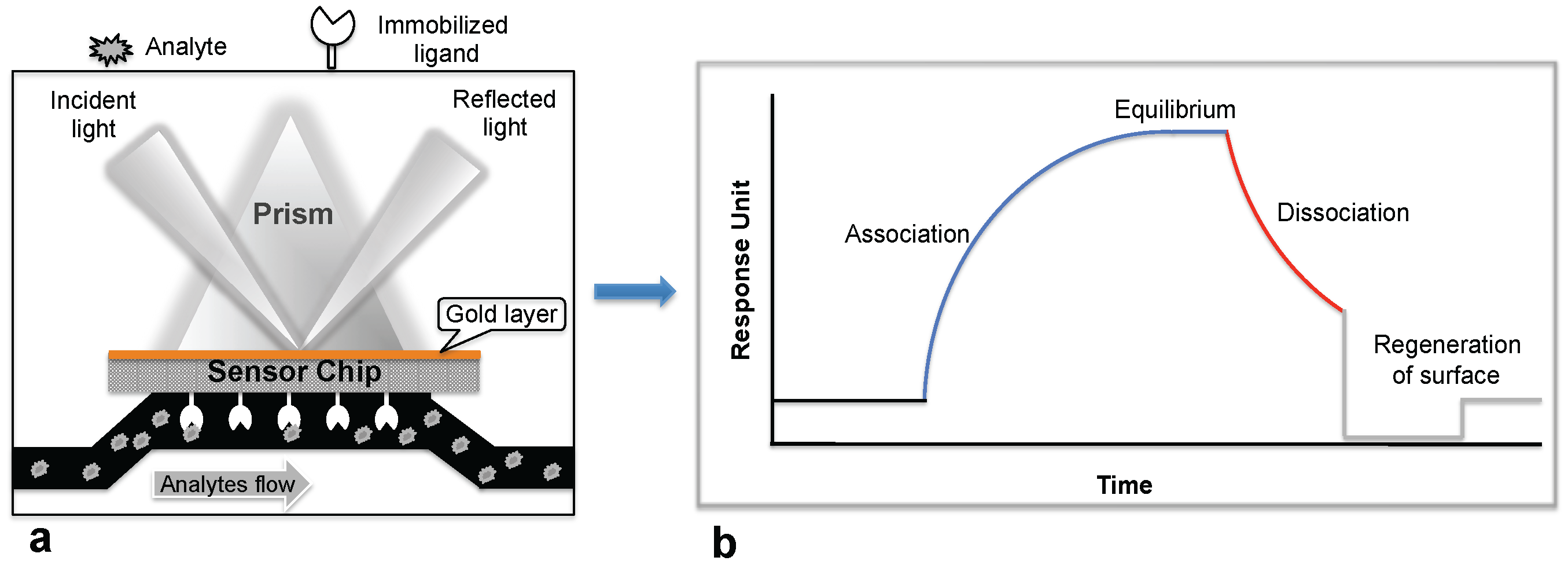
:
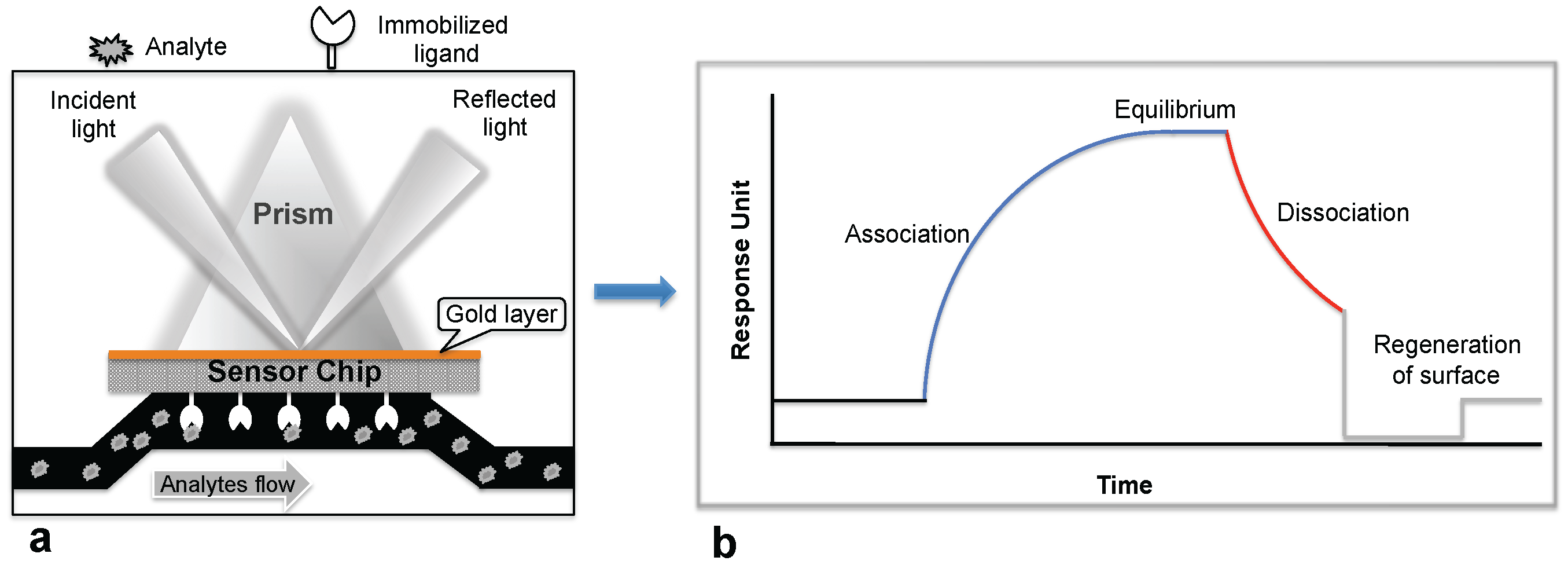
1. Introduction
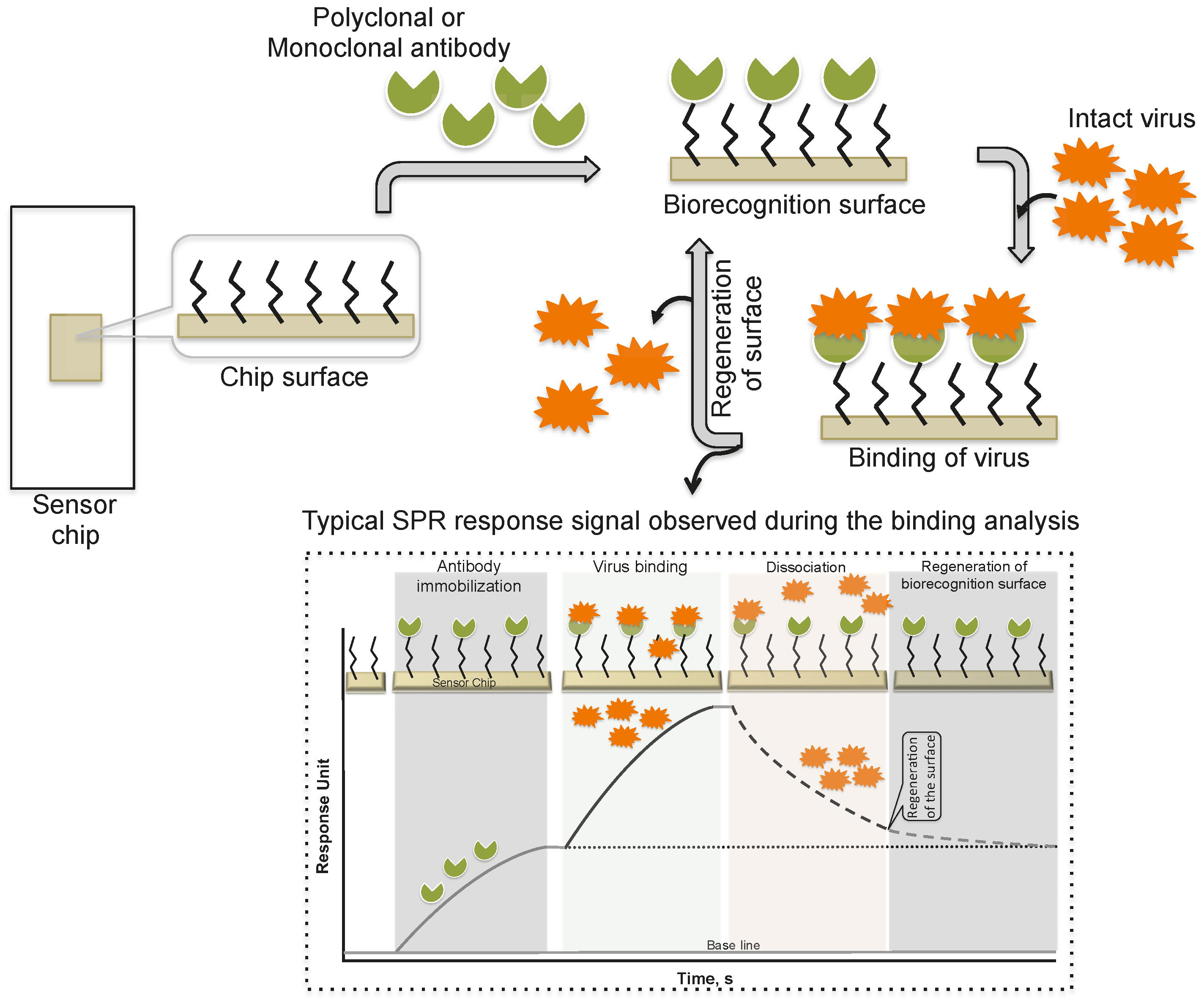
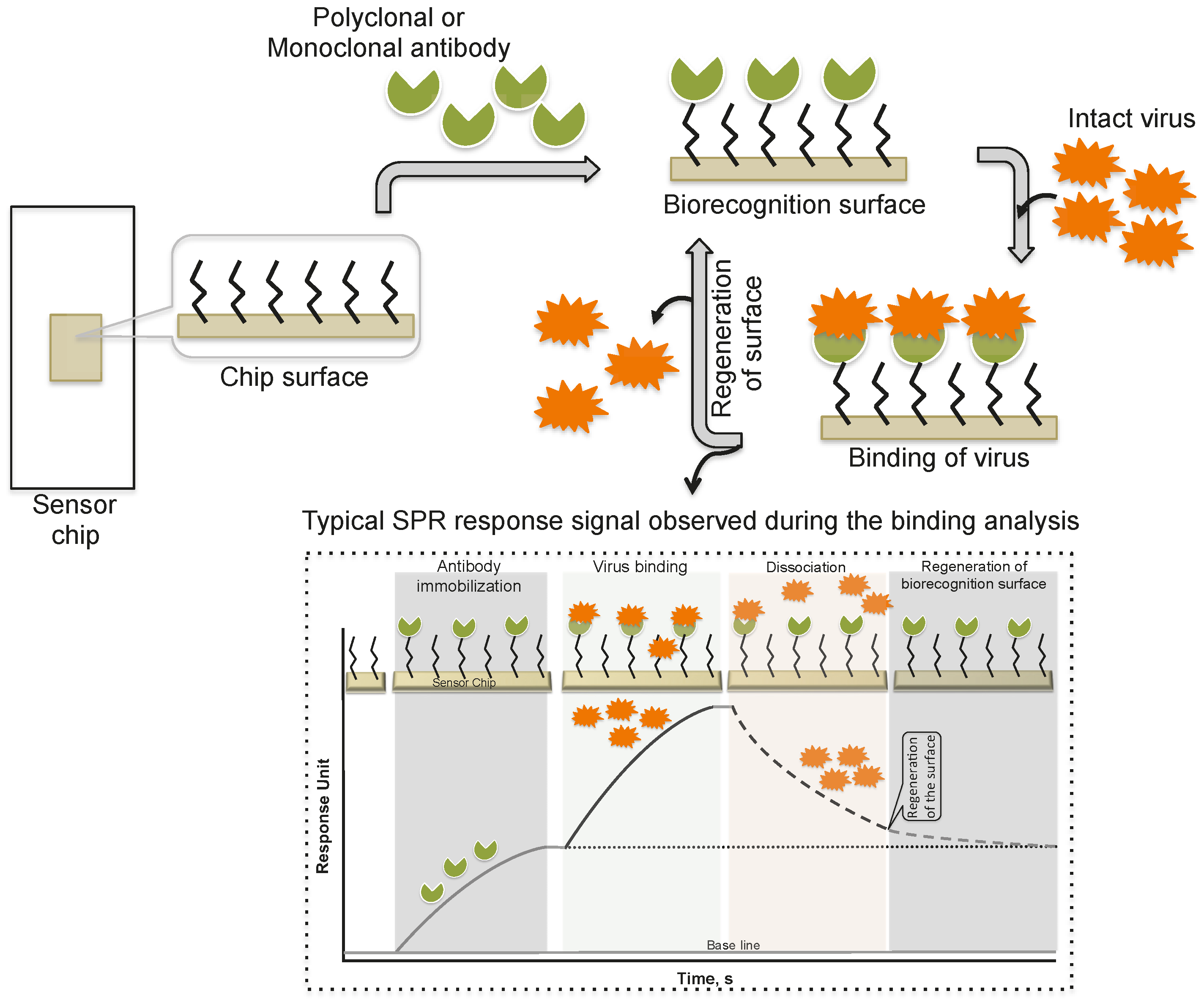
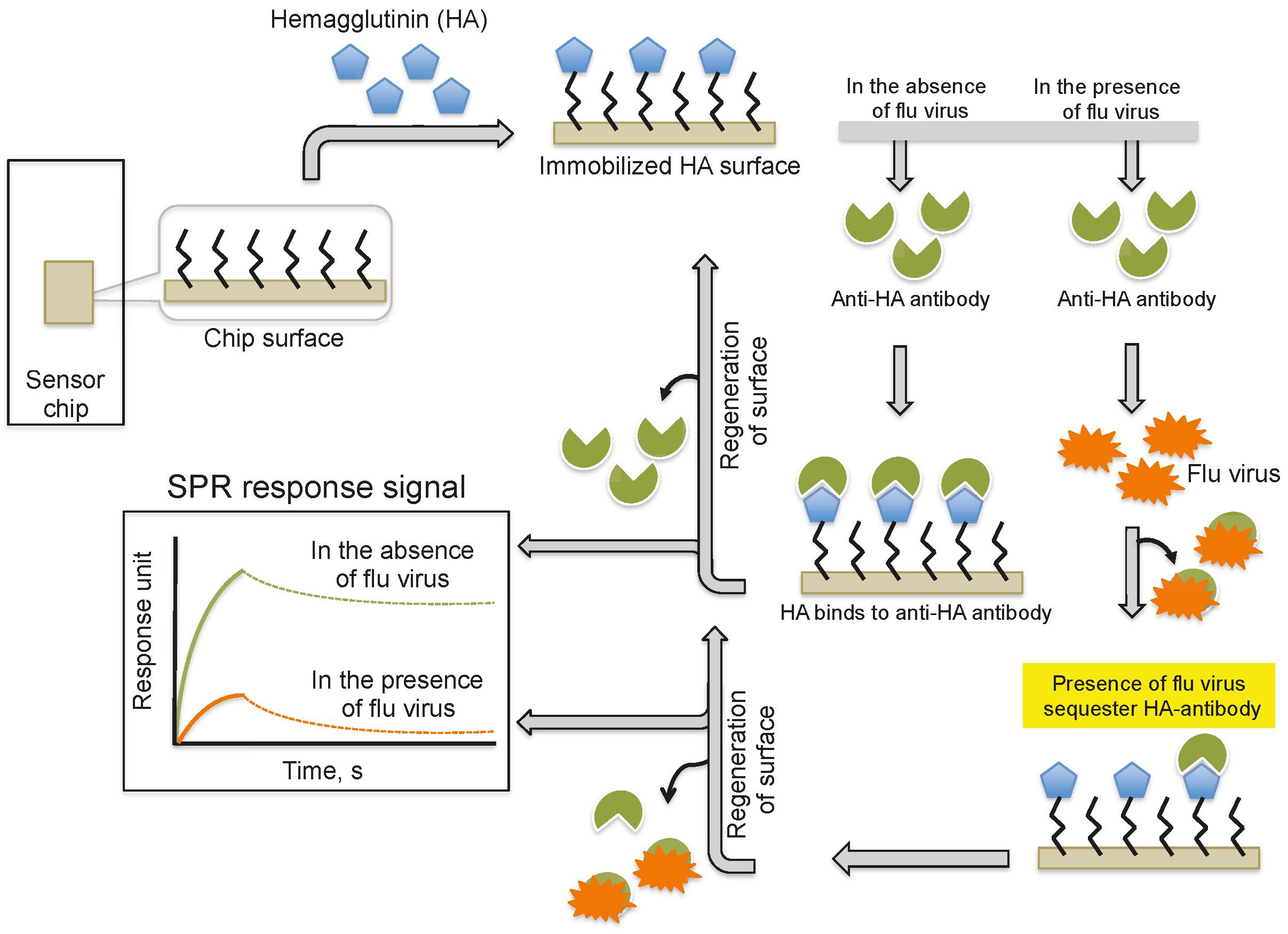
2. Monitoring Intact Viruses Using an Antibody as a Bioreceptor
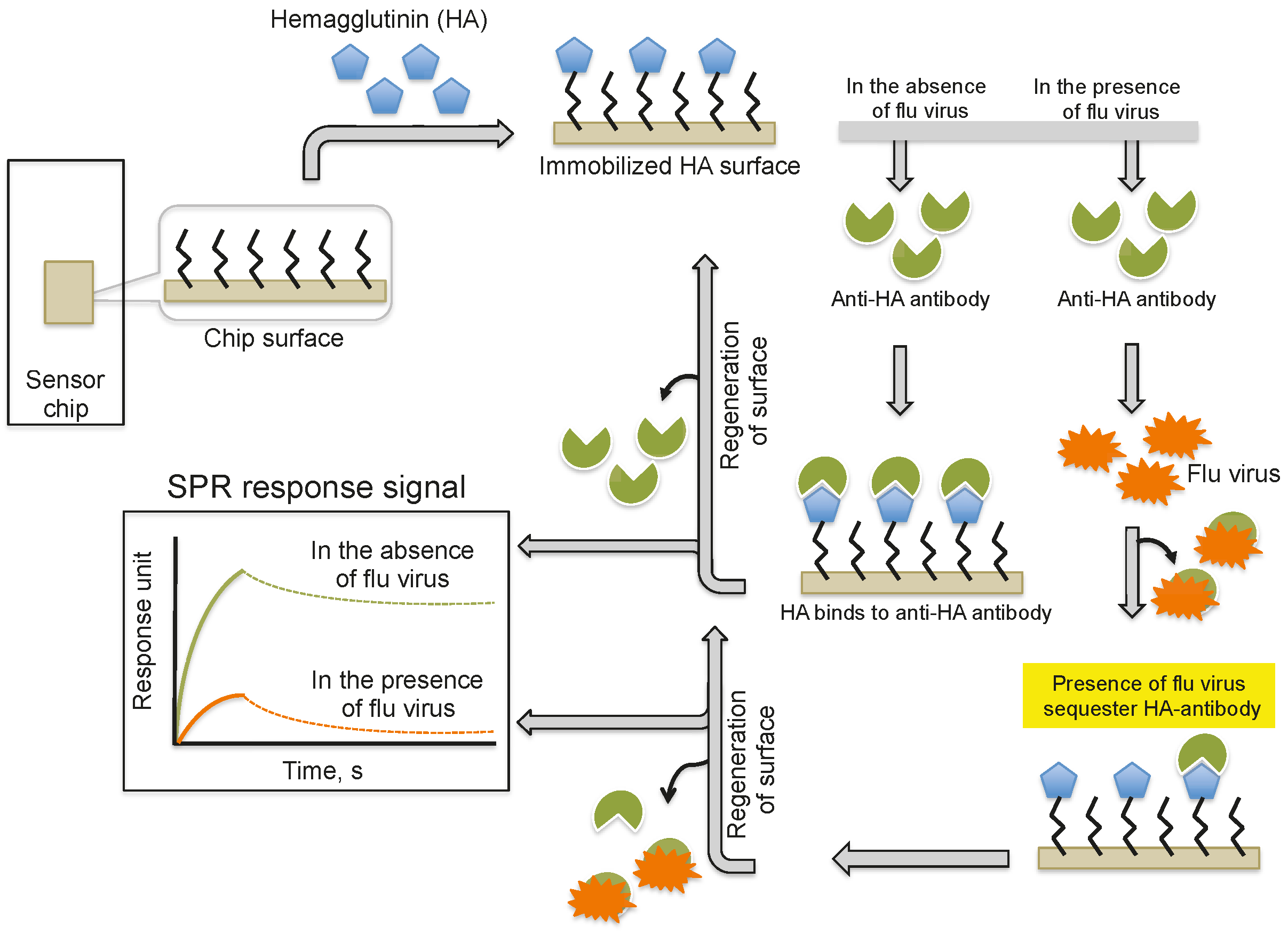
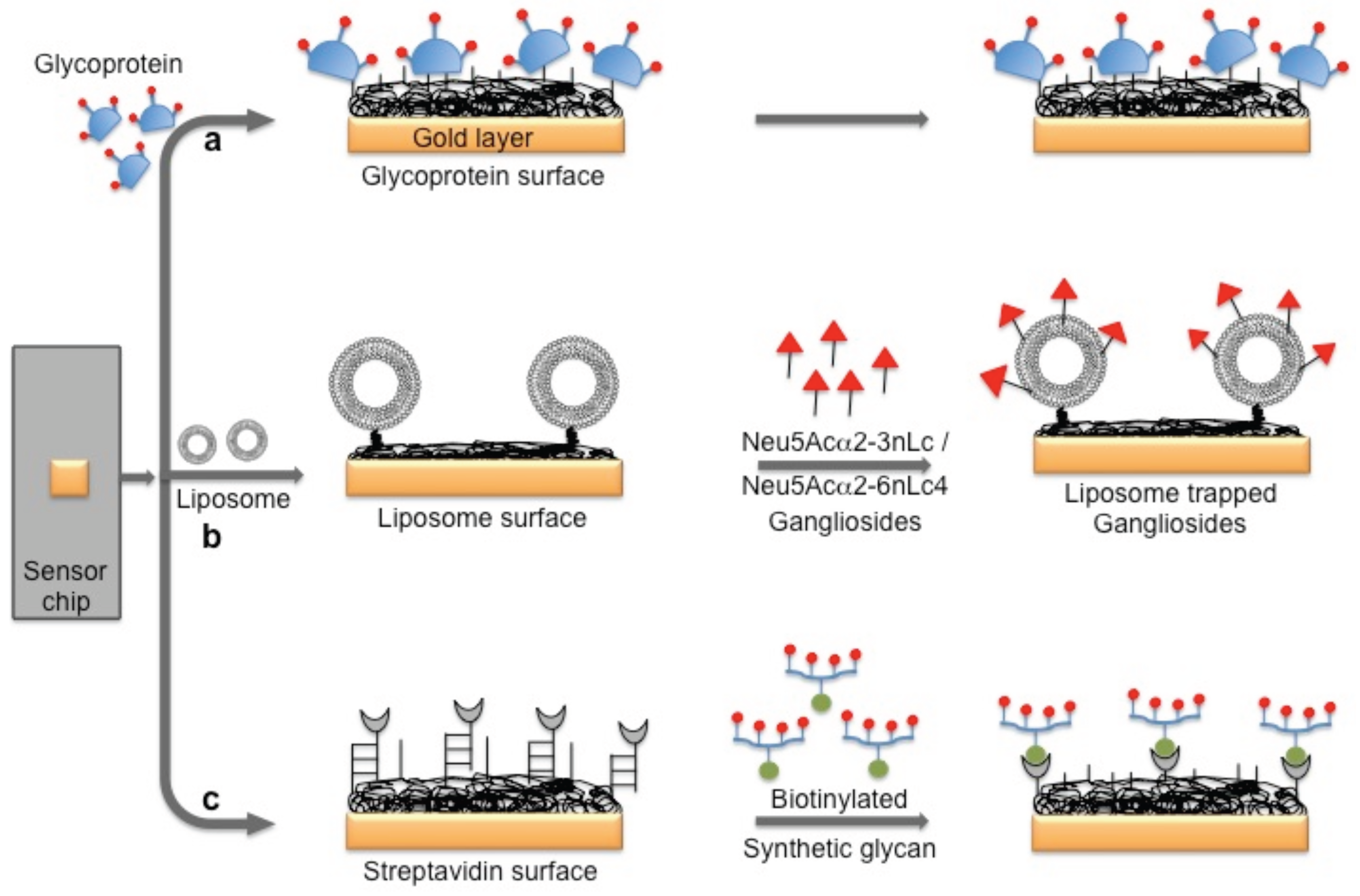
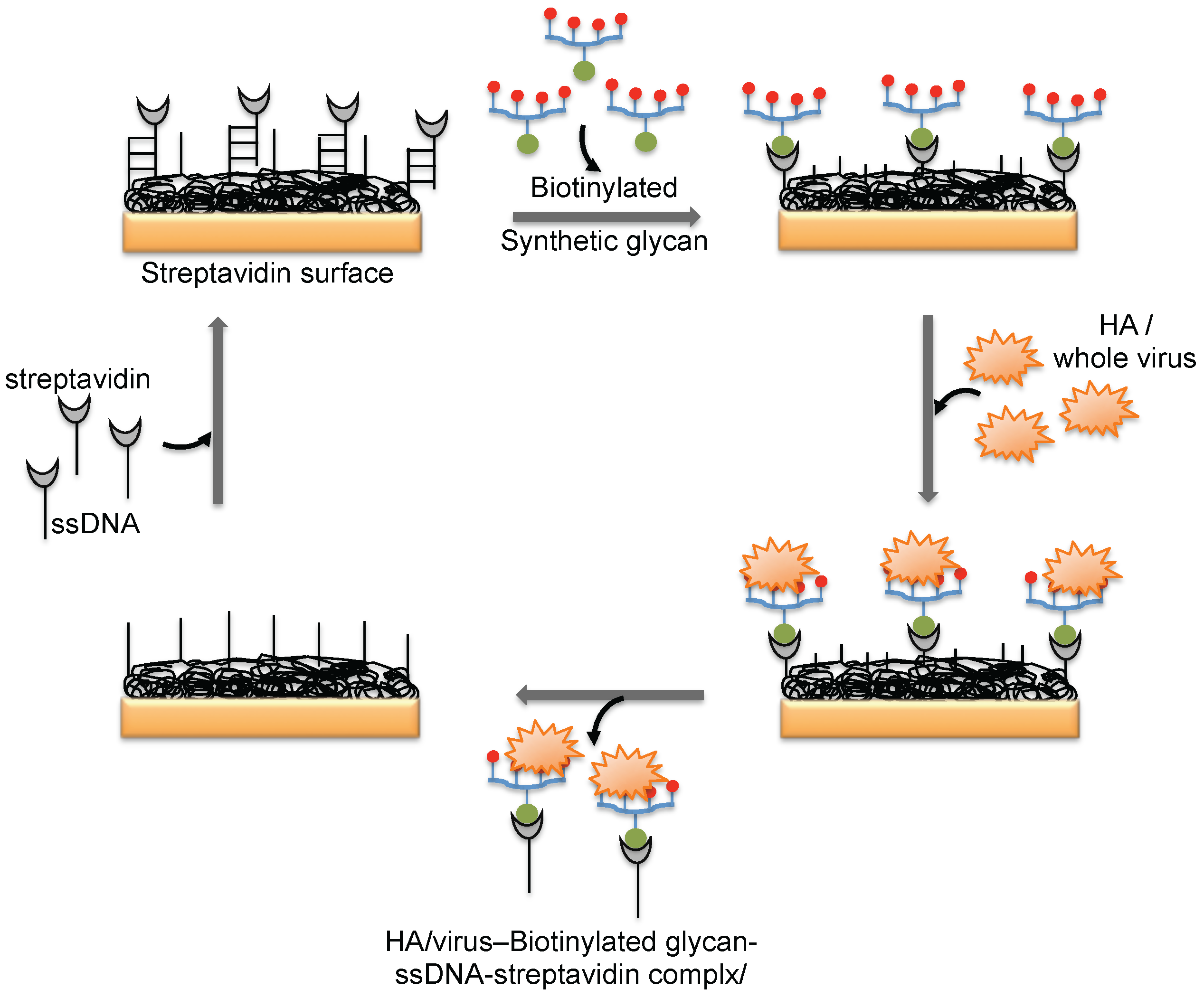
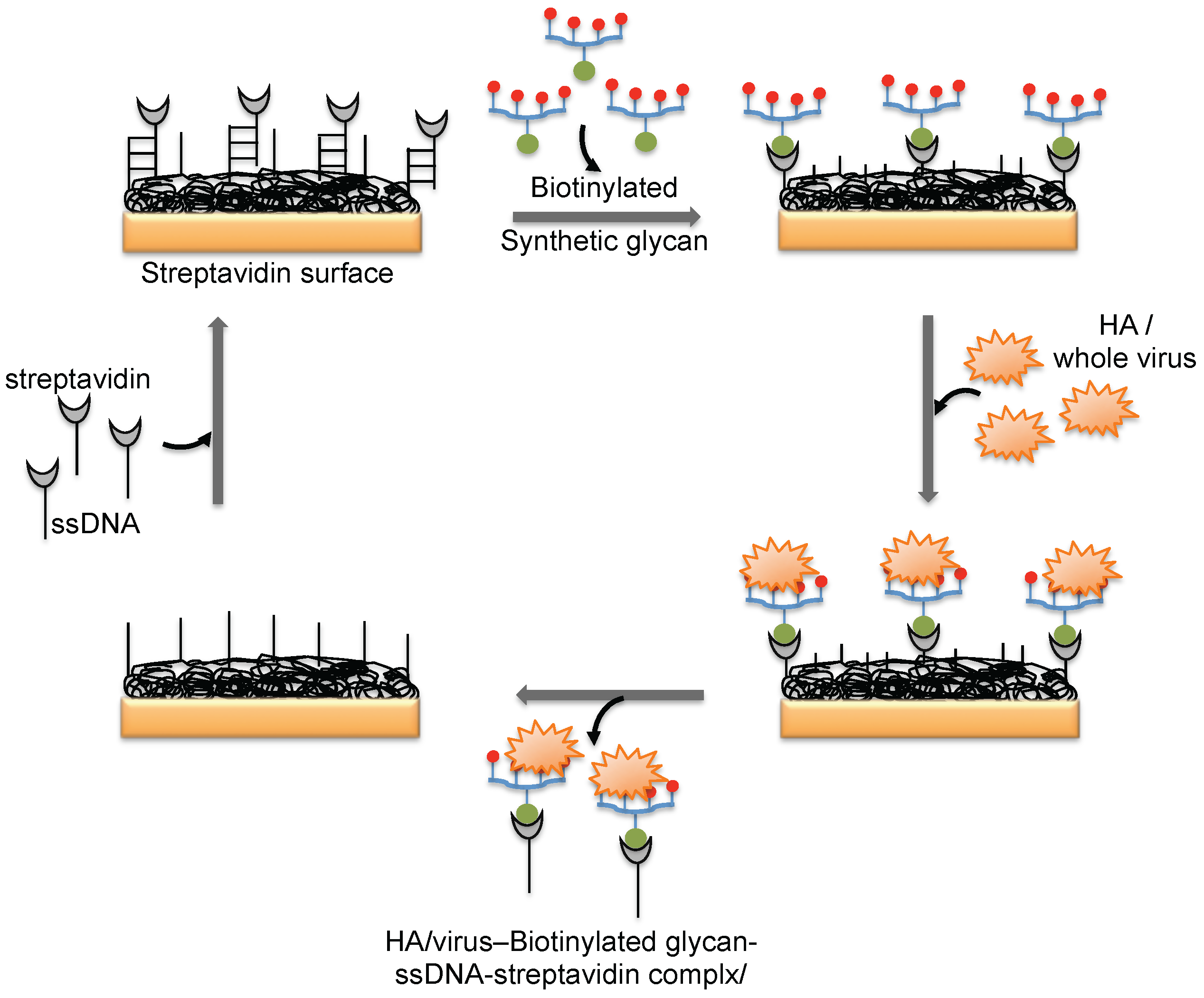
3. Monitoring Intact Viruses Using Glycan as Bioreceptor
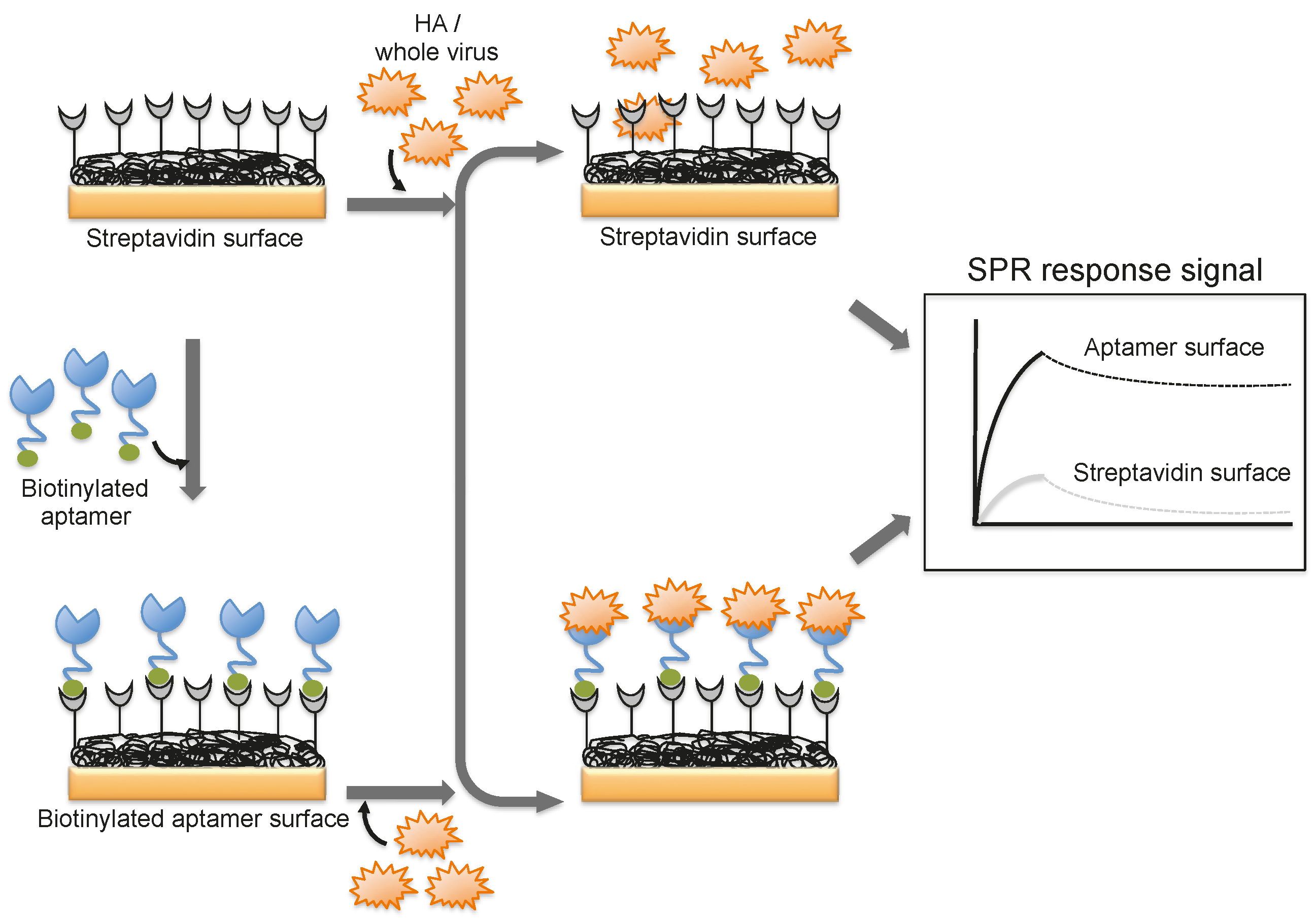
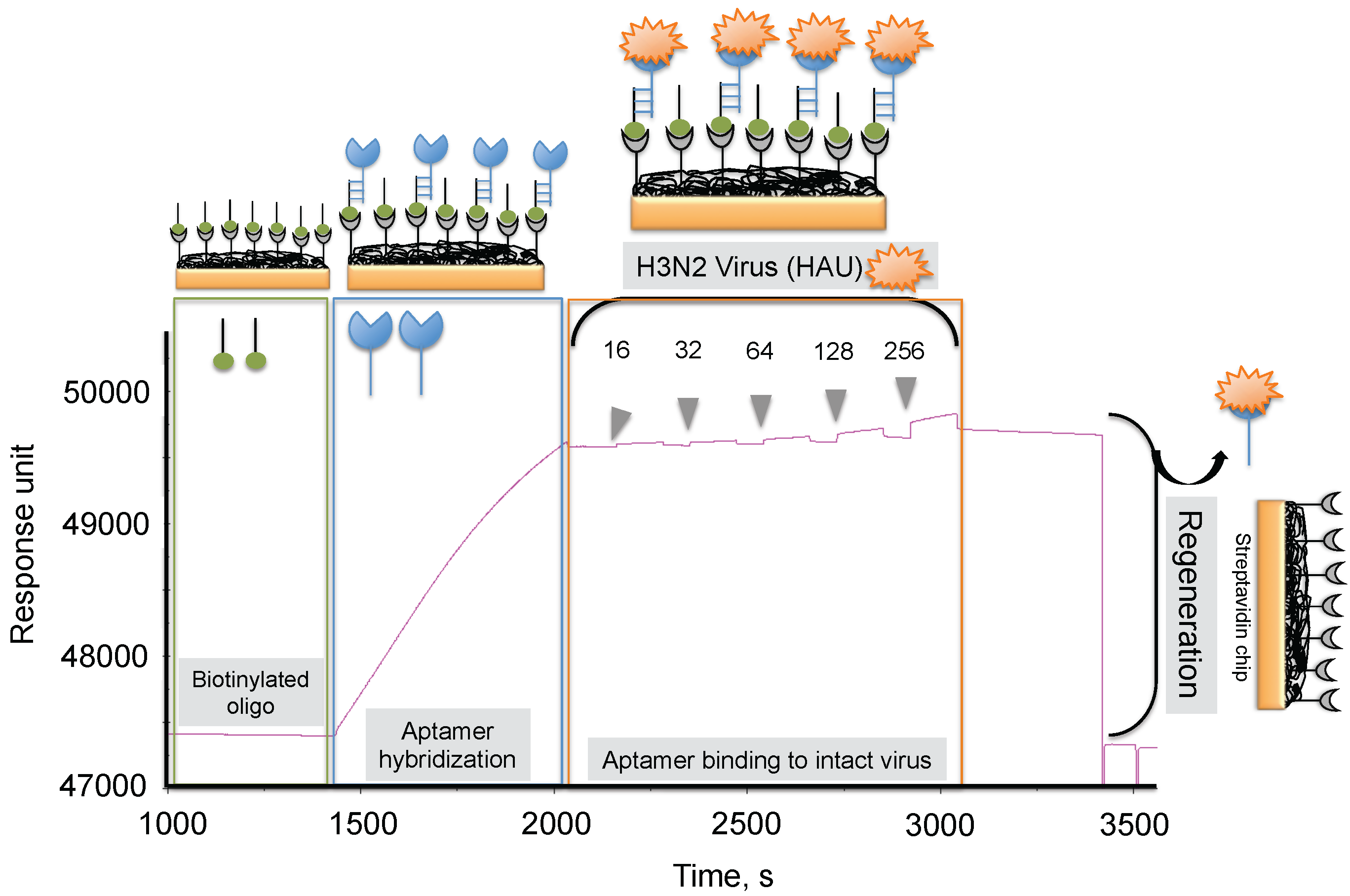
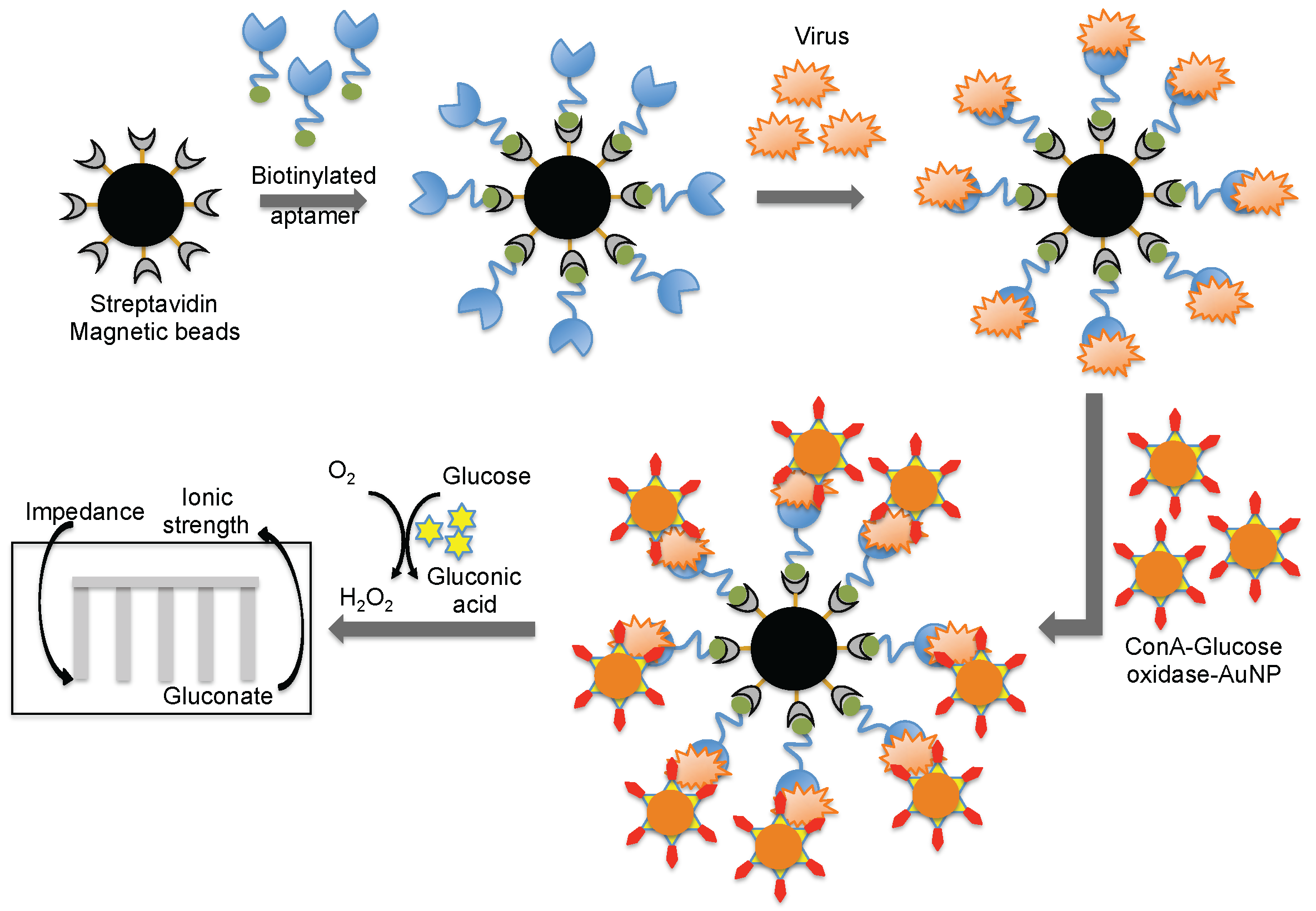
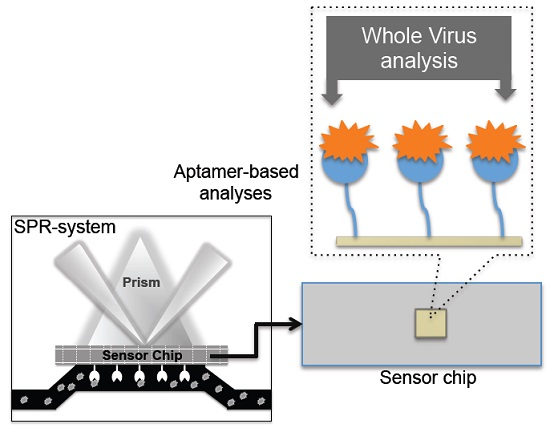
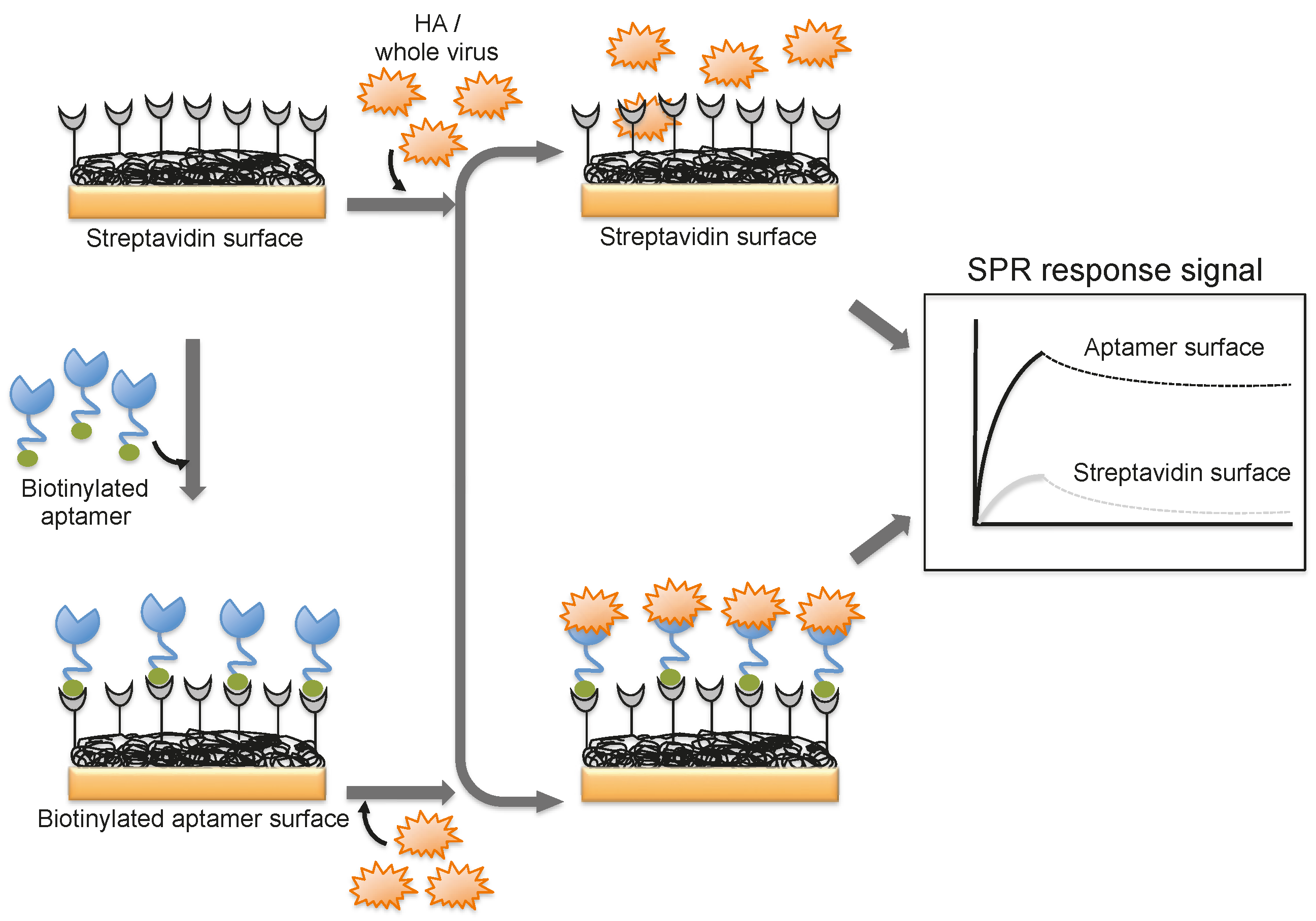
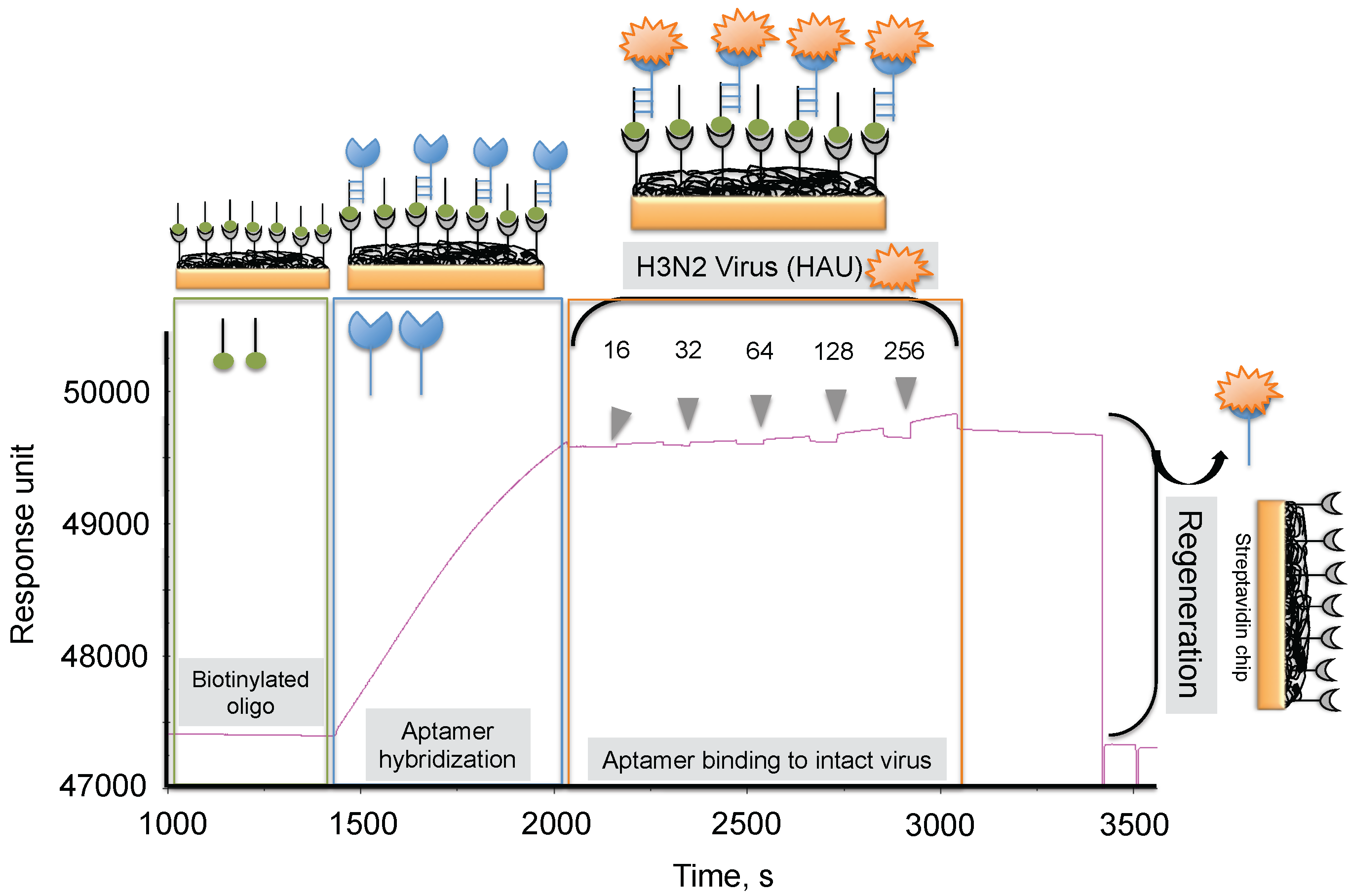
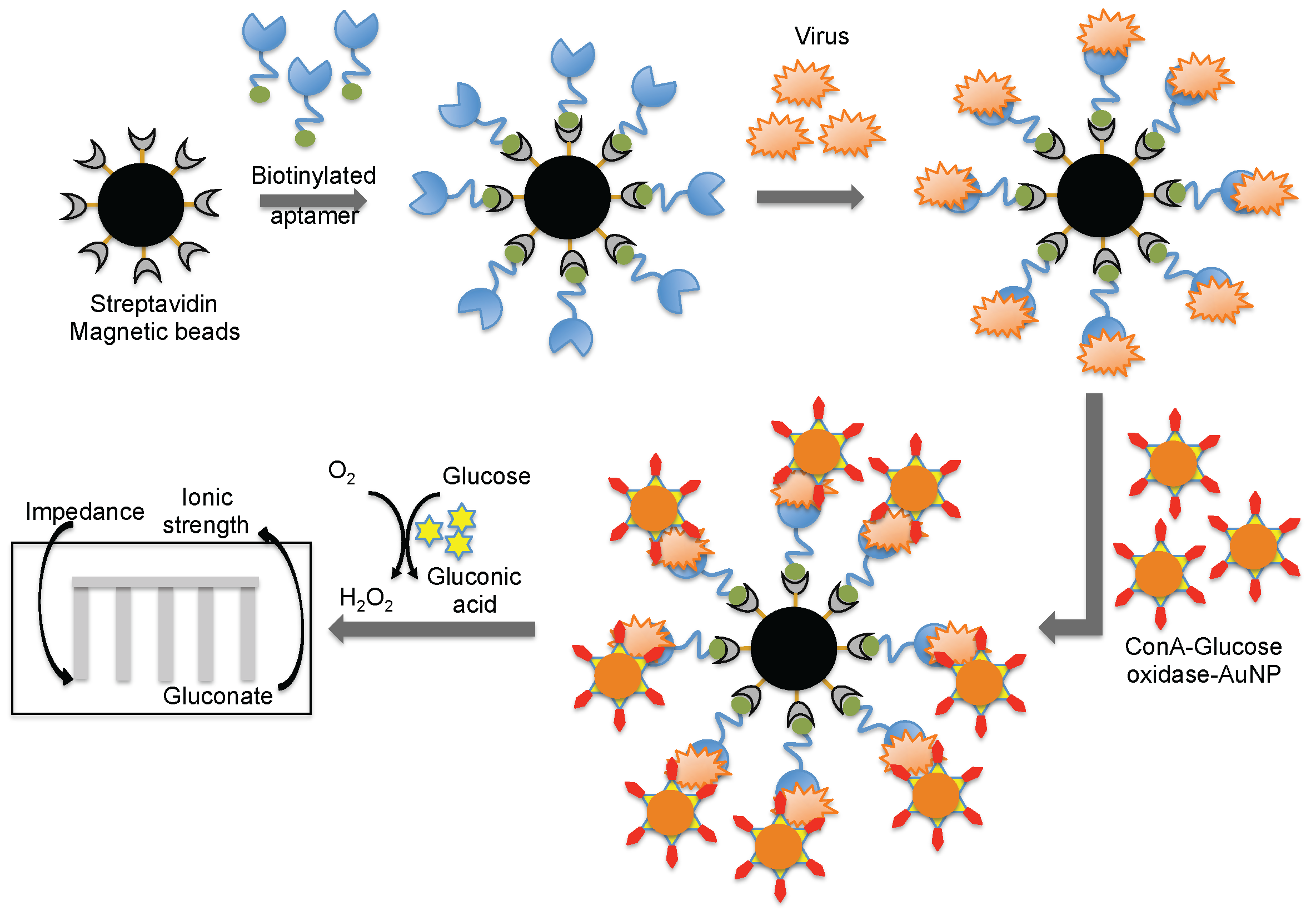
4. Monitoring Intact Viruses Using an Aptamer as a Bioreceptor
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mullis, K.B.; Faloona, F.A. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol. 1987, 155, 335–350. [Google Scholar] [PubMed]
- Mulder, J.; McKinney, N.; Christopherson, C.; Sninsky, J.; Greenfield, L.; Kwok, S. Rapid and simple PCR assay for quantitation of human immunodeficiency virus type 1 RNA in plasma: Application to acute retroviral infection. J. Clin. Microbiol. 1994, 32, 292–300. [Google Scholar] [PubMed]
- Berger, A.; Braner, J.; Doerr, H.W.; Weber, B. Quantification of viral load: Clinical relevance for human immunodeficiency virus, hepatitis B virus and hepatitis C virus infection. Intervirology 1998, 41, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Boeckh, M.; Boivin, G. Quantitation of cytomegalovirus: Methodologic aspects and clinical applications. Clin. Microbiol. Rev. 1998, 11, 533–554. [Google Scholar] [PubMed]
- Schofield, D.J.; Dimmock, N.J. Determination of affinities of a panel of IgGs and Fabs for whole enveloped (influenza A) virions using surface plasmon resonance. J. Virol. Methods 1996, 62, 33–42. [Google Scholar] [CrossRef]
- Hardy, S.A.; Dimmock, N.J. Valency of antibody binding to envelop virus particles as determined by surface plasmon resonance. J. Virol. 2003, 77, 1649–1552. [Google Scholar] [CrossRef] [PubMed]
- Boltovets, P.M.; Snopok, B.A.; Boyko, V.R.; Shevchenko, T.P.; Dyachenko, N.S.; Shirshov, Y.M. Detection of plat viurses using a surface plasmon resonance via complexing with specific antibodies. J. Virol. Methods. 2004, 121, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Baac, H.; Hajos, J.P.; Lee, J.; Kim, D.; Kim, S.J.; Shuler, M.L. Antibody-based surface plasmon resonance detection of intact viral pathogen. Biotechnol. Bioeng. 2006, 94, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, C.E.; Abbas, S.; Bennemo, M.; Laesson, A.; Hamalainen, M.D.; Karlsson, A.F. A novel assay for influenza virus quantification using surface plasmon resonance. Vaccine 2010, 28, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Shan, X.; Patel, U.; Huang, X.; Lu, J.; Li, J.; Tao, N. Lable-free imaging, detection, and mass measurement of single viruses by surface plasmon resonance. Proc. Natl. Acad. Sci. USA 2010, 107, 16028–16032. [Google Scholar] [CrossRef] [PubMed]
- Chenail, G.; Brown, N.E.; Shea, A.; Feire, A.L.; Deng, G. Real-time analysis of antibody interactions with whole enveloped human cytomegalovirus using surface plasmon resonance. Anal. Biochem. 2011, 411, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Yakes, B.J.; Papafragkou, E.; Conrad, S.M.; Neil, J.D.; Ridpath, J.F.; Burkhardt, W.; Kulka, M.; De Grasse, S.L. Surface plasmon resonance biosensor for detection of feline calicivirus, a surrogate for norovirus. Int. J. Food Microbiol. 2013, 162, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Patolsky, F.; Zheng, G.; Hayden, O.; Lakadamyali, M.; Zhuang, X.; Lieber, C.M. Electrical detection of single viruses. Proc. Natl. Acad. Sci. USA 2004, 101, 14017–14022. [Google Scholar] [CrossRef] [PubMed]
- Ymeti, A.; Greve, J.; Lambeck, P.V.; Wink, T.; van Hovell, S.W.F.M.; Beumer, T.A.M.; Wijin, R.R.; Heideman, R.G.; Subramaniam, V.; Kanger, J.S. Fast, ultrasensitive virus detection suing a young interferometer sensor. Nano Lett. 2007, 7, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Hnaien, M.; Diouani, M.F.; Helali, S.; Hafaid, I.; Hassen, W.M.; Renault, N.J.; Ghram, A.; Abdelghani, A. Immobilization of specific antibody on SAM functionalized gold electrode for rabies virus detection by electrochemical impedance spectroscopy. Biochem. Eng. J. 2008, 39, 443–449. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Y.; Lassiter, K.; Li, Y.; Hargis, B.; Tung, S.; Berghman, L.; Bottje, W. Interdigitated array microelectrode based impedance immunosensor for detection of avian influenza virus H5N1. Talanta 2009, 79, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Shirale, D.; Bangar, M.A.; Park, M.; Yates, M.V.; Chen, W.; Myung, N.V.; Mulchandani, A. Label-free chemiresistive immunosensors for viruses. Environ. Sci. Technol. 2010, 44, 9030–9035. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Lin, J.; Lassiter, K.; Srinivasan, B.; Lin, L.; Lu, H.; Tung, S.; Hargis, B.; Bottje, W.; Berghman, L.; Li, Y. Evaluation study of a portable impedance biosensor for detection of avian influenza virus. J. Virol. Methods 2011, 178, 52–58. [Google Scholar] [CrossRef] [PubMed]
- McClellan, M.S.; Domier, L.L.; Bailey, R.C. Label-free virus detection using silicon photonic microring resonators. Biosens. Bioelectron. 2012, 31, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Lum, J.; Wang, R.; Lassiter, K.; Srinivasan, B.; Abi-Ghanem, D.; Berghman, L.; Hargis, B.; Tung, S.; Lu, H.; Li, Y. Rapid detection of avian influenza H5N1 virus suing impedance measurement of immune-reaction coupled with RBC amplification. Biosens. Bioelectron. 2012, 38, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.C.B.; Awazu, K.; Fujimaki, M.; Shimizu, K.; Shima, T. Observations of immune-gold conjugates on influenza viruses suing Waveguide-mode sensors. PLoS ONE 2013, 8, e69121. [Google Scholar] [CrossRef]
- Baca, J.T.; Severns, V.; Lovato, D.; Branch, D.W.; Larson, R.S. Rapid detection of Ebola virus with a reagent-free point-of-care biosensor. Sensors 2015, 15, 8605–8614. [Google Scholar] [CrossRef] [PubMed]
- Baker, M. Blame it on the antibodies. Nature 2015, 521, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Barton, E.S.; Connolly, J.L.; Forrest, J.C.; Chappell, J.D.; Dermody, T.S. Utilization of sialic acid as a coreceptor enhances reovirus attachment by multiple adhesion strengthening. J. Biol. Sci. 2001, 276, 2200–2211. [Google Scholar]
- Shinya, K.; Ebina, M.; Yamada, S.; Ono, M.; Kasai, N.; Kawaoka, Y. Avian flu: Influenza virus receptors in the human airway. Nature 2006, 440, 435–436. [Google Scholar] [CrossRef] [PubMed]
- Hidari, K.I.P.J.; Shimada, S.; Suzuki, Y.; Suzuki, T. Binding kinetics of influenza viruses to sialic acid-containing carbohydrates. Glycoconj. J. 2007, 24, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, E.; Mizuno, H.; Kumar, P.K.R. Monitoring influenza hemagglutinin and glycan interactions using surface plasmon resonance. Biosens. Bioelectron. 2012, 32, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, E.; Mizuno, H.; Kumar, P.K.R. Influenza virus surveillance using surface plasmon resonance. Virulence 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, E.; Gopinath, S.C.B.; Tanaka, M.; Kumar, P.K.R. Monitoring the whole influenza virus using a surface plasmon resonance sensor. Unpublished data.
- Hushegyi, A.; Pihikova, D.; Bertok, T.; Adam, V.; Kizek, R.; Tkac, J. Ultrasensitive detection of influenza viruses with a glycan-based impedimetric biosensor. Biosens. Bioelectron. 2016, 79, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.; Polisky, B.; Uhlenbeck, O.; Yarus, M. Diversity of oligonucleotide functions. Annu. Rev. Biochem. 1995, 64, 763–797. [Google Scholar] [CrossRef] [PubMed]
- Osborne, S.E.; Ellington, A.D. Nucleic acid selection and the challenge of combinatorial chemistry. Chem. Rev. 1999, 97, 349–370. [Google Scholar] [CrossRef]
- Wilson, D.S.; Szostak, J.W. In vitro selection of functional nucleic acids. Annu. Rev. Biochem. 1999, 68, 611–647. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.J.; Lee, J.W.; Ellington, A.D. Applications of aptamers as sensors. Annu. Rev. Anal. Chem. 2009, 2, 241–264. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.C.B.; Misono, T.S.; Kumar, P.K.R. Prospects of ligand-induced aptamers. Crit. Rev. Anal. Chem. 2008, 38, 34–47. [Google Scholar] [CrossRef]
- Keefe, A.D.; Pai, S.; Ellington, A.D. Aptamers as therapeutics. Nat. Rev. Drug Discov. 2010, 9, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, C.A.; Cai, W.; Hong, H. Applications of aptamers in targeted imaging: Sate of the art. Curr. Top. Med. Chem. 2015, 15, 1138–1152. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.C.B. Antiviral aptamers. Arch. Virol. 2007, 152, 2137–2157. [Google Scholar] [CrossRef] [PubMed]
- Shum, K.T.; Zhou, J.; Rossi, J.J. Aptamer-based therapeutics: New approaches to combat human viral diseases. Pharmaceuticals 2013, 6, 1507–1542. [Google Scholar] [CrossRef] [PubMed]
- Misono, T.S.; Kumar, P.K.R. Selection of RNA aptamers against human influenza virus hemagglutinin using surface plasmon resonance. Anal. Biochem. 2005, 342, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.C.B.; Misono, T.S.; Kawasaki, K.; Mizuno, T.; Imai, M.; Odagiri, T.; Kumar, P.K.R. An RNA aptamer that distinguishes between closely related human influenza viruses and inhibits haemagglutinin-mediated membrane fusion. J. Gen. Virol. 2006, 87, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.C.B.; Sakamaki, Y.; Kawasaki, K.; Kumar, P.K.R. An efficient RNA aptamer against human influenza B virus hemagglutinin. J. Biochem. 2006, 139, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Kim, S.; Yoon, H.; Kim, K.B.; Kalme, S.S.; Oh, S.; Song, C.S.; Kim, D.E. Selection of an antiviral RNA aptamer against hemagglutinin of the subtype H5 avian influenza virus. Nucleic Acid Ther. 2011, 21, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhao, J.; Jiang, T.; Kwon, Y.M.; Lu, H.; Jiao, P.; Liao, M.; Li, Y. Selection and characterization of DNA aptamers for use in detection of avian influenza virus H5N1. J. Virol. Methods 2013, 189, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.C.B.; Kumar, P.K.R. Aptamers that bind to the hemagglutinin of the recent pandemic influenza virus H1N1 and efficiently inhibit agglutinin. Acta. Biomater. 2013, 9, 8932–8941. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, E.; Kumar, P.K.R. An Aptamer that binds efficiently to the hemagglutinins of highly pathogenic avian influenza viruses (H5N1 and H7N7) and inhibits hemagglutinn-glycan interactions. Acta. Biomater. 2014, 10, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.C.B.; Hayashi, K.; Kumar, P.K.R. Aptamer that binds to the gD protein of herpes simplex virus 1 and efficiently inhibits viral entry. J. Virol. 2012, 86, 6732–6744. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Wang, R.; Hargis, B.; Lu, H.; Li, Y. A SPR aptasensor for detection of avian influenza virus H5N1. Sensors 2012, 12, 12506–12518. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.H.; Kayhan, B.; Ben-Yedidia, T.; Arnon, R. A DNA aptamer prevents influenza infection by blocking the receptor binding region of the viral hemagglutinin. J. Biol. Chem. 2004, 279, 48410–48419. [Google Scholar] [CrossRef] [PubMed]
- Kiilerich-Pedersen, K.; Dapra, J.; Cherre, S.; Rozlosnik, N. High sensitivity point-of-care for direct virus diagnosis. Biosens. Bioelectron. 2013, 49, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Callaway, Z.; Lum, J.; Lin, J.; Li, Y. Exploiting enzyme catalysis in ultra-low ion strength media for impedance biosensing of avain influenza virus using a bare interdigitated electrode. Anal. Chem. 2013, 86, 1965–1971. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, Y. Hydrogel based QCM aptasensor for detection of avian influenza virus. Biosens. Bioelectron. 2013, 42, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Butler, D. Flu surveillance lacking. Nature 2012, 483, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Wandtke, T.; Wozniak, J.; Kopinski, P. Apatmers in diagnostics and treatment of viral infections. Viruses 2015, 7, 751–780. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Hunag, J.J.; Ding, J.; Liu, J. Aptamer-based biosensors for biomedical diagnostics. Analyst 2014, 139, 2627–2640. [Google Scholar] [CrossRef] [PubMed]
- Soelberg, S.D.; Chinowsky, T.; Geiss, G.; Spinelli, C.B.; Stevens, R.; Near, S.; Kauffman, P.; Yee, S.; Furlong, C.E. A potable surface plasmon resonance sensor system for real-time monitoring of small to large analytes. J. Ind. Microbiol. Biotechnol. 2005, 32, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Chinowsky, T.; Soelberg, S.D.; Baker, P.; Swanson, N.R.; Kauffman, P.; Mactitis, A.; Grow, M.S.; Atmar, R.; Yee, S.S.; Furlong, C.E. Portable 24-analyte surface plasmon resonance instruments for rapid, versatile biodetection. Biosens. Bioelectron. 2007, 22, 2268–2275. [Google Scholar] [CrossRef] [PubMed]
- Seattle Sensor Systems. Available online: http://www.seattlesensors.com/products.html (accessed on 2 August 2016).
- SPR Micro (KMAC). Available online: http://www.kmac.com/eng/products.php?cid=sprmicro (accessed on 2 August 2016).
- Biosuplar 6 (Analytical Micro-Systems/Mivitec). Available online: http://www.biosuplar.de (accessed on 2 August 2016).
- Bruno, J.G.; Carrillo, M.P.; Richarte, A.M.; Phillips, T.; Anfrews, C.; Lee, J.S. Development, screening, and analysis of DNA aptamer libraries potentially useful for diagnosis and passive immunity of arboviruses. BMC Res. 2012, 5, e633. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, Y.; Hu, B.; Ma, Z.Y.; Huang, H.P.; Yu, Y.; Liu, S.P.; Lu, M.J.; Yang, D.L. Development of HBsAg-binding aptamers that bind HepG2.2.15 cells via HBV surface antigen. Virol. Sin. 2010, 25, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Jee, M.H.; Kwon, O.S.; Keum, S.J.; Jang, S.K. Infectivity of hepatitis C virus correlates with the amount of envelope protein E2: Development of a new aptamer-based assay system suitable for measuring the infectious titer of HCV. Virology 2013, 439, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiang, H.; Liu, F. In vitro selection of novel RNA ligands that bind human cytomegalovirus and block viral infection. RNA 2000, 6, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Sayer, N.; Ibrahim, J.; Turner, K.; Tahiri-Alaoui, A.; James, W. Structural characterization of a 2'F-RNA aptamer that binds a HIV-1 SU glycoprotein, gp120. Biochem. Biophys. Res. Commun. 2002, 293, 924–931. [Google Scholar] [CrossRef]
- Allen, P.; Collins, B.; Brown, D.; Hostomsky, Z.; Gold, L. A specific RNA structural motif mediates high affinity binding by the HIV-1 nucleocapsid protein (NCp7). Virology 1996, 226, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.D.; Bunka, D.H.; Forzan, M.; Spear, P.G.; Stockley, P.G.; McGowan, I.; James, W. Generation of neutralizing aptamers against herpes simplex virus type 2: Potential components of multivalent microbicides. J. Gen. Virol. 2011, 92, 1493–1499. [Google Scholar] [CrossRef] [PubMed]
- Kiilerich-Pedersen, K.; Daprà, J.; Cherré, S.; Rozlosnik, N. High sensitivity point-of-care device for direct virus diagnostics. Biosens. Bioelectron. 2013, 49, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Shiratori, I.; Akitomi, J.; Boltz, D.A.; Horii, K.; Furuichi, M.; Waga, I. Selection of DNA aptamers that bind to influenza A viruses with high affinity and broad subtype specificity. Biochem. Biophys. Res. Commun. 2014, 443, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Musafia, B.; Oren-Banaroya, R.; Noiman, S. Designing anti-influenza aptamers: Novel quantitative structure activity relationship approach gives insights into aptamer–virus interaction. PLoS ONE 2015, 9, e97696. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.Q.; Ren, Q.; Wei, H.P.; Chen, Z.; Deng, J.Y.; Zhang, Z.P.; Zhang, X.E. Quantum dot-aptamer nanoprobes for recognizing and labeling influenza a virus particles. Nanoscale 2011, 3, 2454–2457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, Z.; Jiang, F.; Fu, P.; Shen, J.; Wu, W.; Li, J. Two DNA aptamers against avian influenza H9N2 virus prevent viral infection in cells. PLoS ONE 2015, 10, e0123060. [Google Scholar] [CrossRef] [PubMed]
- Lakshmipriya, T.; Fujimaki, M.; Gopinath, S.C.; Awazu, K. Generation of anti-influenza aptamers using the systematic evolution of ligands by exponential enrichment for sensing applications. Langmuir 2013, 29, 15107–15115. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.R.; Hu, G.Q.; Zhang, T.; Yang, Y.J.; Zhao, L.L.; Qi, Y.L.; Wang, H.L.; Gao, Y.W.; Yang, S.T.; Xia, X.Z. Isolation of ssDNA aptamers that inhibit rabies virus. Int. Immunopharmacol. 2012, 14, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Craven, R.C.; Qiu, Q.; Wilson, C.B.; Wills, J.W.; Golovine, S.; Wang, J.F. Isolation of virus-neutralizing RNAs from a large pool of random sequences. Proc. Natl. Acad. Sci. USA 1995, 92, 11509–11513. [Google Scholar] [CrossRef] [PubMed]
- Labib, M.; Zamay, A.S.; Muharemagic, D.; Chechik, A.V.; Bell, J.C.; Berezovski, M.V. Aptamer-based viability impedimetric sensor for viruses. Anal. Chem. 2012, 84, 1813–1816. [Google Scholar] [CrossRef] [PubMed]
- Parekh, P.; Tang, Z.; Turner, P.C.; Moyer, R.W.; Tan, W. Aptamers recognize glycosylated hemagglutinin expressed on the surface of vaccinia virus-infected cells. Anal. Chem. 2010, 82, 8642–8649. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Parekh, P.; Turner, P.; Moyer, R.W.; Tan, W. Generating aptamers for recognition of virus-infected cells. Clin. Chem. 2009, 55, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Lautner, G.; Balogh, Z.; Bardoczy, V.; Meszaros, T.; Gyurcsanyi, R.E. Aptamer-based biochips for label-free detection of plant virus coat proteins by SPR imaging. Analyst 2010, 135, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Houser-Scott, F.; Ansel-McKinney, P.; Cai, J.M.; Gehrke, L. In vitro genetic selection analysis of alfalfa mosaic virus coat protein binding to 3’-terminal AUGS repeats in the viral RNAs. J. Virol. 1997, 71, 2310–2319. [Google Scholar] [PubMed]
- Schneider, D.; Tuerk, C.; Gold, L. Selection of high affinity RNA ligands to the bacteriophage R17 coat protein. J. Mol. Biol. 1992, 228, 862–869. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Approximate Size (nm) | Sensor Chip | Buffer | Bioreceptor | Reference |
|---|---|---|---|---|---|
| A/fowl plague/Rostock/34 (H7N1) | 120 | CM5 | PBS | Monoclonal antibody | [5] |
| A/Puerto Rico/8/34 (H1N1) | 120 | SA | HBS-EP | Monoclonal antibody | [6] |
| Tobacco mosaic virus | 180 | Custom | Carbonate | Polyclonal antibody | [7] |
| Autograph californica multiple nuclear polyhedrosis virus | 240 | Custom | PBS | Monoclonal antibody | [8] |
| B/Brisbane/3/2007 | 120 | CM5 | HBS-EP+ | Polyclonal antibody | [9] |
| A/Solomon Islands/3/2006 (H1N1) | 120 | CM5 | HBS-EP+ | Polyclonal antibody | [9] |
| A/PR/8/34 (H1N1) | 120 | CM5 | HBS-EP+ | Polyclonal antibody | [9] |
| A/Wsiconsin/67/2005 (H3N2) | 120 | CM5 | HBS-EP+ | Polyclonal antibody | [9] |
| A/PR/8/34 (H1N1) | 120 | Custom | PBS | Monoclonal antibody | [10] |
| Human cytomegalovirus | 230 | Custom | PBS | Monoclonal antibody | [10] |
| Human cytomegalovirus | 230 | CM3/CM5 | PBS + 0.05% Tween20 | Monoclonal antibody | [11] |
| Feline calicivirus (F-9 strain, VR-782) | 30 | CM3 | HBS-EP+ | Polyclonal antibody | [12] |
| Virus | Detection Method | Bioreceptor | Reference |
|---|---|---|---|
| Influenza virus A | Nanowire field effect transistors | Monoclonal Antibody | [13] |
| Herpes simplex virus-1 | Interferometer sensor | Monoclonal Antibody | [14] |
| Rabies virus | Impedance spectroscopy | Polyclonal Antibody | [15] |
| Avian Influneza virus [A/Scotland 59 (H5N1)] | Microelectrode based Impedance spectroscopy | Polyclonal Antibody | [16] |
| Bacteriophages T7/ MS2 | Nanowire electrochemical | Monoclonal Antibody | [17] |
| Avian Influneza virus [A/ck/PA/87 (H5N2)] | Impedance spectroscopy | Monoclonal Antibody | [18] |
| Bean pod mottle virus | Photonic microring resonators | Monoclonal Antibody | [19] |
| Avian Influneza virus [A/Scotland 59 (H5N1)] | Impedance biosensor | Monoclonal Antibody | [20] |
| A/Udon/307/1972 (H3N2) | Waveguide-mode sensor | Monoclonal Antibody | [21] |
| A/Brisbane/10/2007 (H3N2) | Waveguide-mode sensor | Monoclonal Antibody | [22] |
| Ebola virus (Zaire) | Surface acoustic wave | Monoclonal Antibody | [23] |
| Virus | Apamer Target | Reference |
|---|---|---|
| Chikungunya/Dengue/West Nile | Viral Envelop proteins | [ 61] |
| HBV | Surface antigen | [62] |
| HCV | E2 Glycoprotein | [63] |
| Human cytomegalovirus | Whole virus | [64] |
| HIV | Gp120 | [65] |
| HIV | Nucleocapsid | [66] |
| HSV-1 | gD | [47] |
| HSV-2 | gD | [67] |
| Influenza A (H1N1) | ||
| California/2007/1999 | HA | [45] |
| PR/8/34 | HA | [24,68] |
| Brisbane/59/07 | HA | [69] |
| California/04/09 | HA | [69] |
| Singapore/6/86 | HA | [69] |
| Georgia/20/06 | HA | [69] |
| Perth/265/09 | HA | [70] |
| Influenza A (H2N2) | ||
| Japan/57 | HA | [24] |
| Influenza A (H3N2) | ||
| Panama/2007/1999 | HA | [40,41] |
| Brisbane/10/07 | HA | [69] |
| Wisconsin/67/05 | HA | [69] |
| Moscow/10/99 | HA | [69] |
| Texas/77 | HA | [24,70] |
| Port Chalmers/1/73 | HA | [24] |
| Guizhou/54/89 | HA | [25] |
| Influenza A (H5N1) | ||
| Vietnam/1203/2004 | HA | [71] |
| Vietam/1194/2004 | HA | [46] |
| Indonesia/05/2005 | HA | [46] |
| Anhui/1/05 | HA | [69] |
| Influenza A (H7N7) | ||
| Netherlands/219/2003 | HA | [46] |
| Influenza A (H9N2) | ||
| Beijing/1/01 | HA | [72] |
| Hebei/3/98 | HA | [72] |
| Influenza B | ||
| Johannesburg/05/99 | HA | [42] |
| Tokio/53/99 | HA | [73] |
| Jilin/20/03 | HA | [73] |
| Rabies Virus | Whole virus | [74] |
| Rous sarcoma virus | Whole virus | [75] |
| Vaccinia | Whole virus/HA | [76,77] |
| Surface protein | [78] | |
| Other viruses | ||
| Apple stem pitting virus | Coat protein | [79] |
| Alpha mosaic virus | Coat protein | [80] |
| Bacteriophage R17 | Coat protein | [81] |
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, P.K.R. Monitoring Intact Viruses Using Aptamers. Biosensors 2016, 6, 40. https://doi.org/10.3390/bios6030040
Kumar PKR. Monitoring Intact Viruses Using Aptamers. Biosensors. 2016; 6(3):40. https://doi.org/10.3390/bios6030040
Chicago/Turabian StyleKumar, Penmetcha K. R. 2016. "Monitoring Intact Viruses Using Aptamers" Biosensors 6, no. 3: 40. https://doi.org/10.3390/bios6030040
APA StyleKumar, P. K. R. (2016). Monitoring Intact Viruses Using Aptamers. Biosensors, 6(3), 40. https://doi.org/10.3390/bios6030040