
SPR Biosensor Probing the Interactions between TIMP-3 and Heparin/GAGs



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Heparin Biochip

2.3. Measurement of Interaction between Heparin and TIMP-3 Using BIAcore
2.4. Solution Competition Study between Heparin on Chip Surface and Heparin-Derived Oligosaccharides in Solution Using SPR

2.5. Solution Competition Study between Heparin on Chip Surface and GAGs, Chemically Modified Heparin in Solution Using SPR
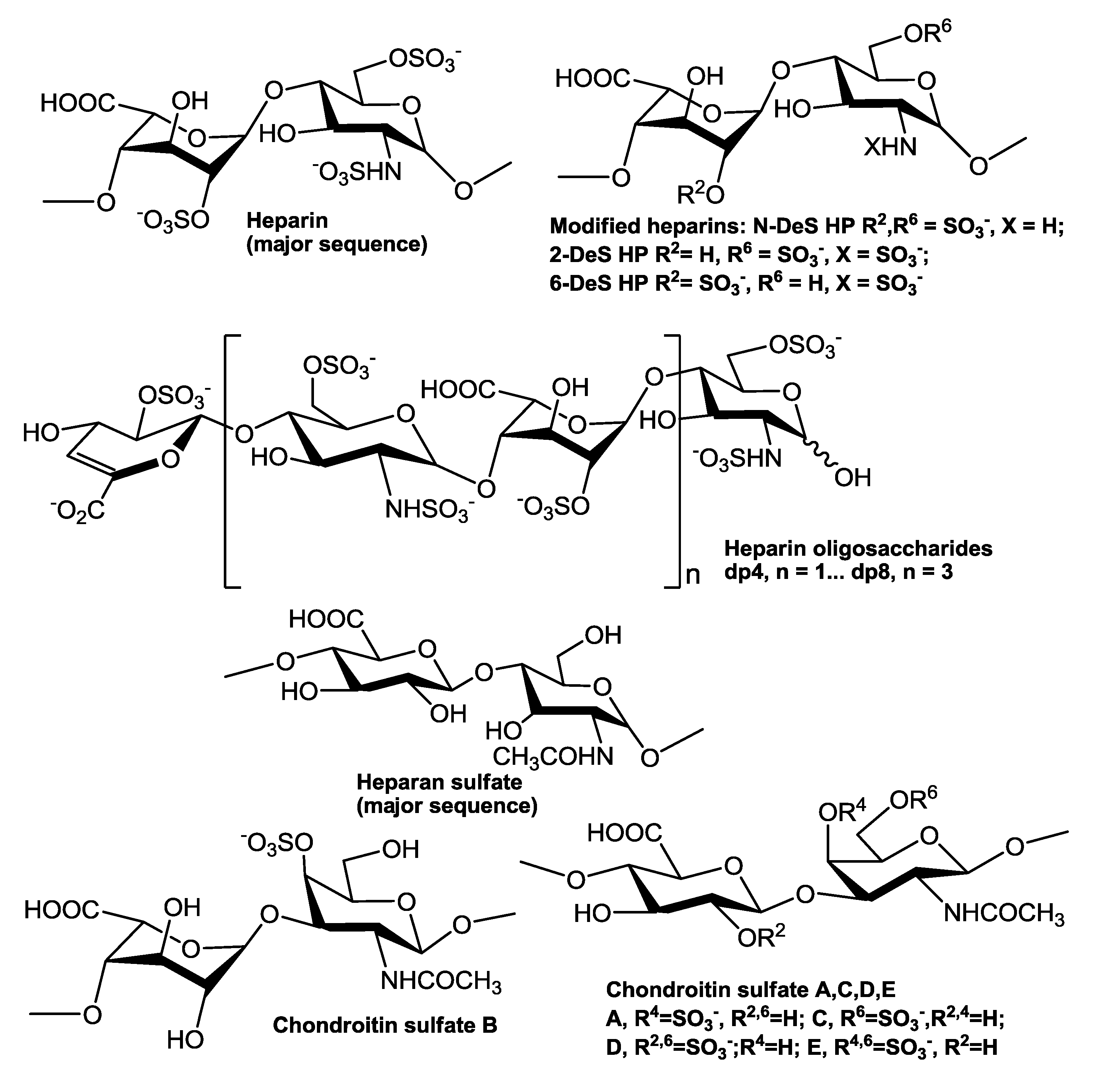
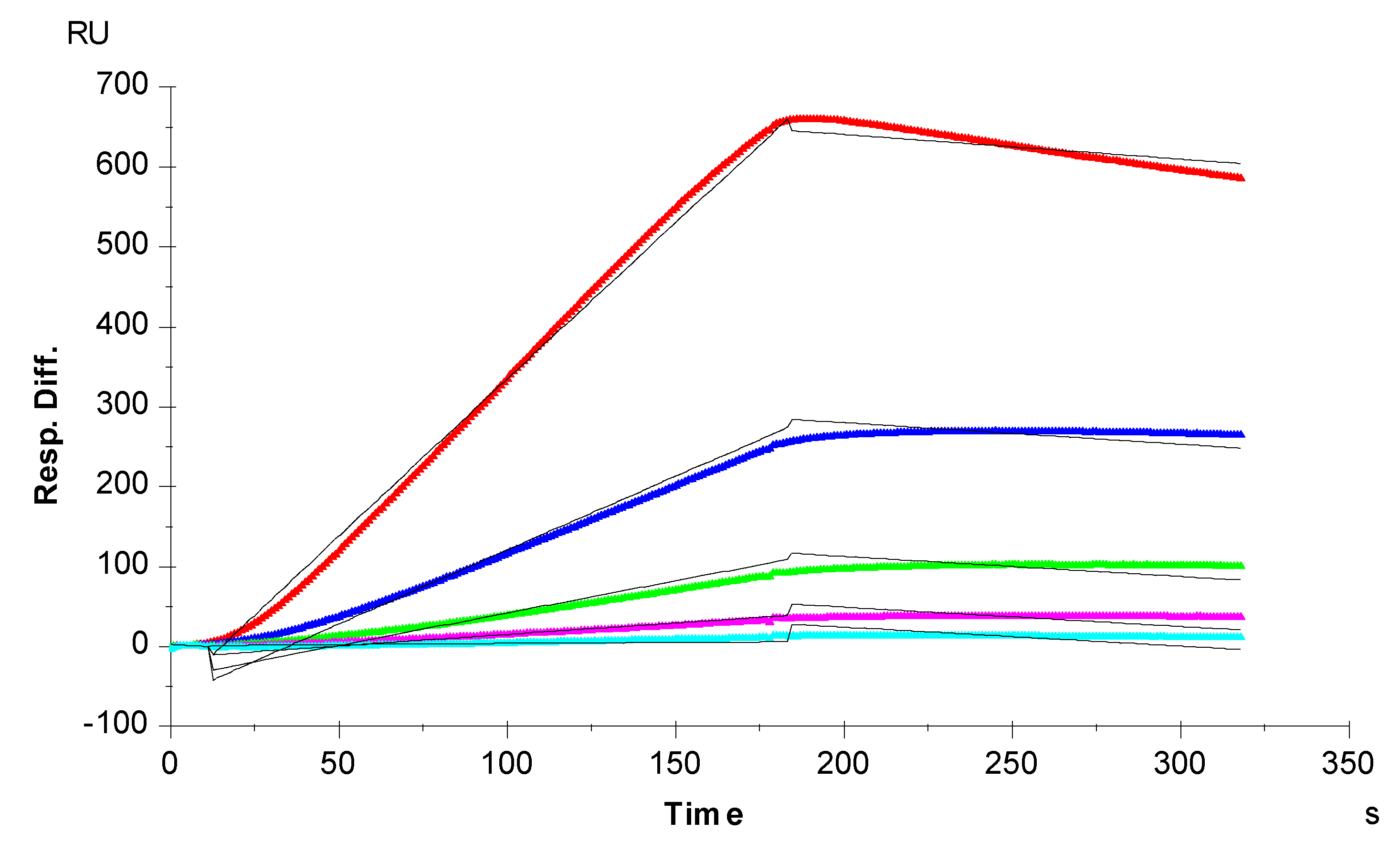
3. Results and Discussion
3.1. Kinetics Measurement of TIMP-3-Heparin Interactions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Interaction | ka (1/MS) | kd (1/S) | KD (M) |
|---|---|---|---|
| TIMP-3/Heparin | 1800 (±93) | 1.1 × 10−4 (±1.4 × 10−5) | 5.9 × 10−8 |
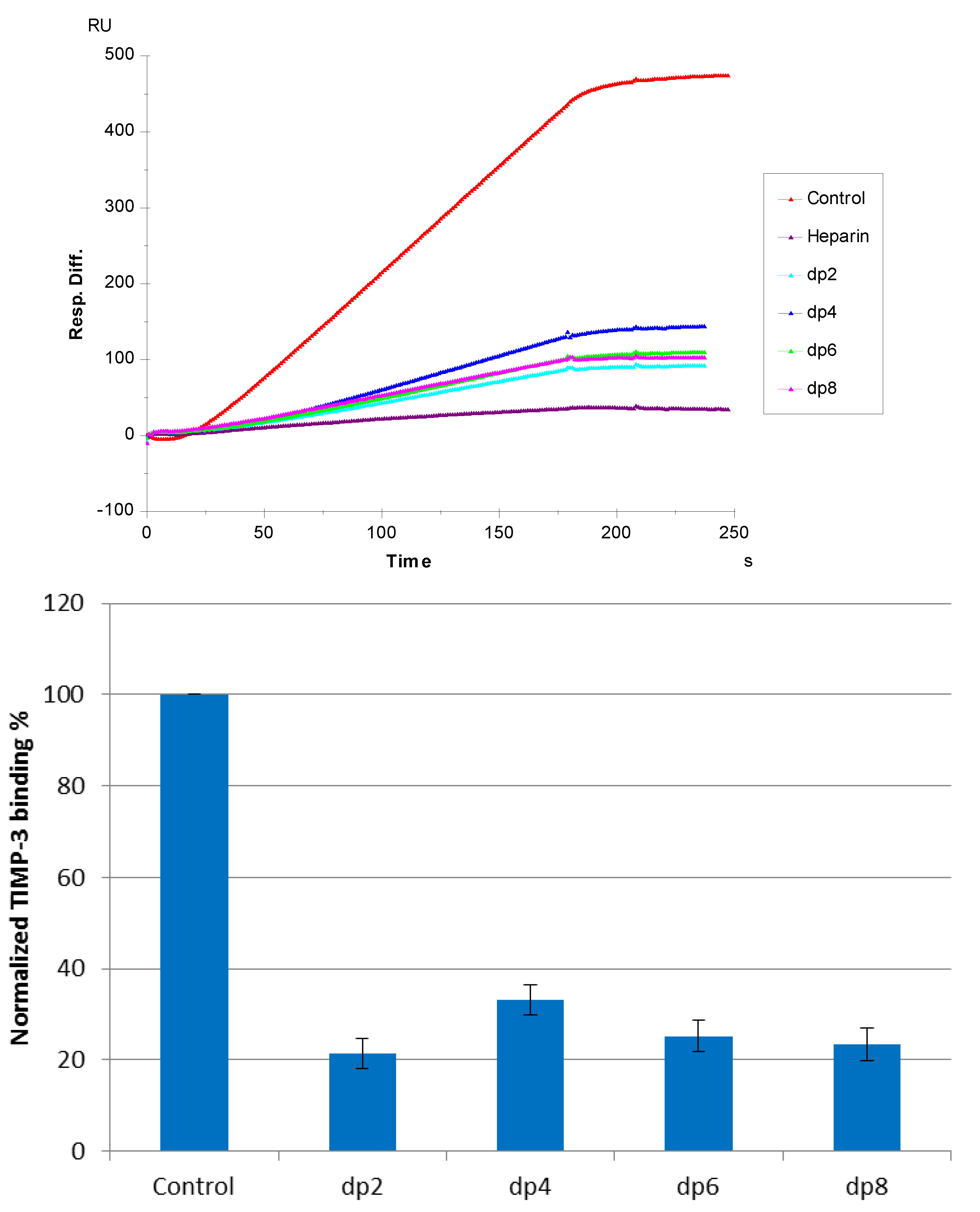
3.2. Solution Competition SPR Study Using Heparin-Derived Oligosaccharides
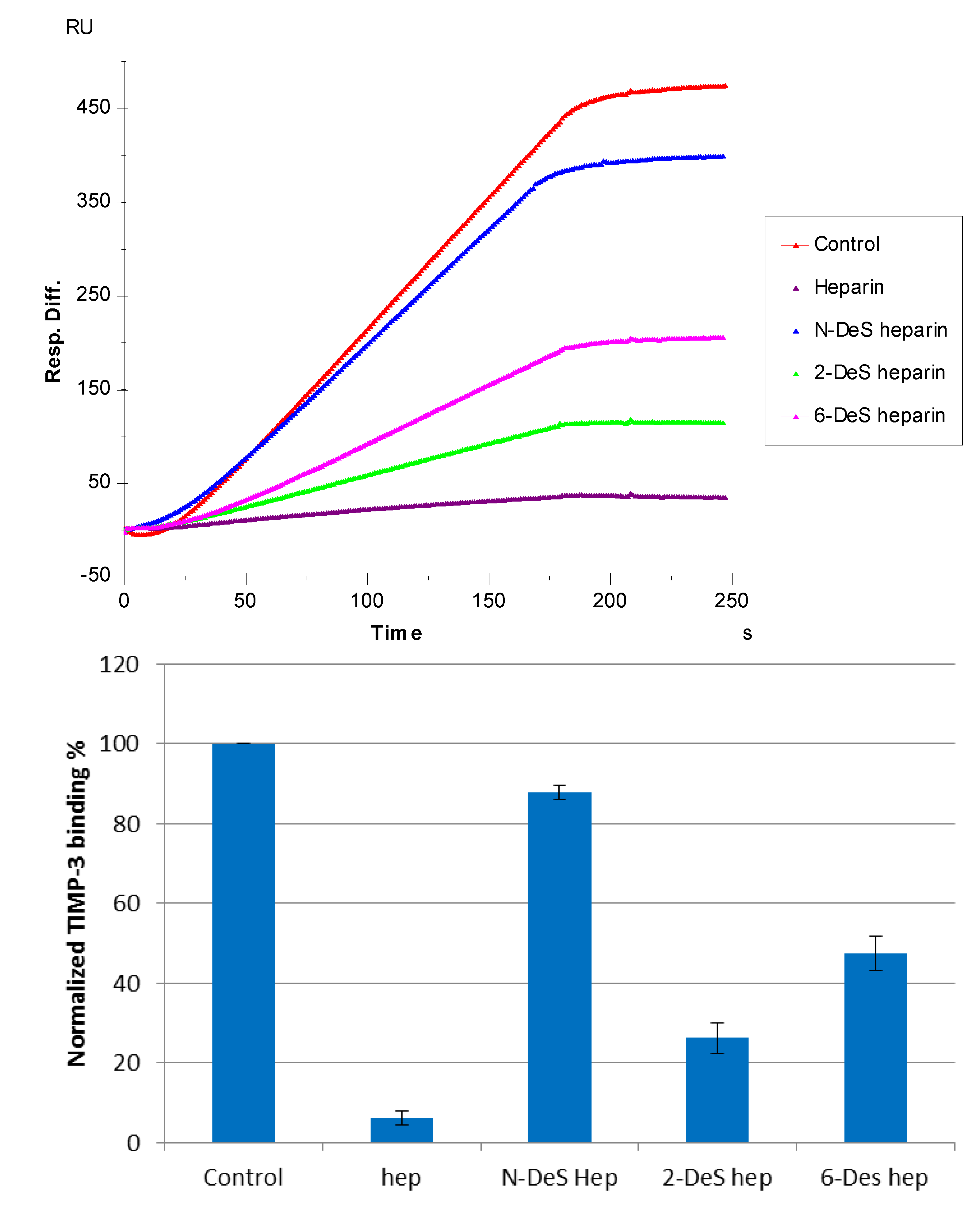
3.3. SPR Solution Competition Study of Different GAGs

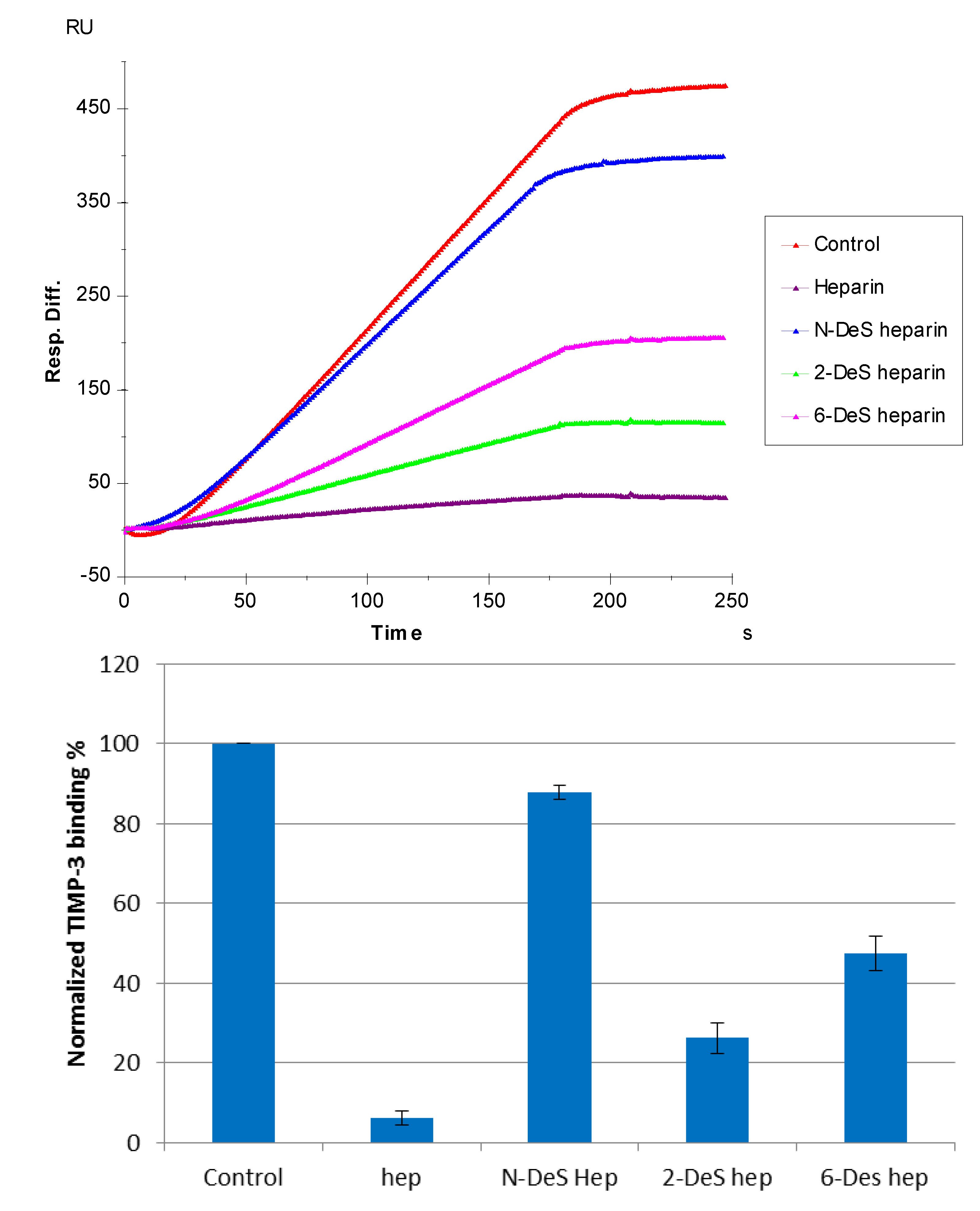
3.4. SPR Solution Competition Study of Different Chemically Modified Heparins
Conclusions
Abbreviations
| TIMP-3 | tissue inhibitor of metalloproteinases-3 |
| GAG | glycosaminoglycan |
| MMP | matrix metalloproteinase |
| SPR | surface plasmon resonance |
| HS | heparan sulfate |
| CS-A | chondroitin sulfate type A |
| CS-B | chondroitin sulfate type B |
| CS-C | chondroitin sulfate type C |
| CS-D | chondroitin sulfate type D |
| CS-E | chondroitin sulfate type E |
| SA | streptavidin |
| FC | flow-cell |
| RU | resonance unit |
| dp | degree of polymerization |
| ADAM | disintigrin and metalloprotease |
| ADMTS | ADAM with thrombospondin motifs |
| TNF-α | tumor necrosis factor-α |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Markey, F. What is SPR anyway? BIA. J. 1999, 6, 14–17. [Google Scholar]
- Jönsson, U.; Fägerstam, L.; Ivarsson, B.; Johnsson, B.; Karlsson, R.; Lundh, K.; Löfås, S.; Persson, B.; Roos, H.; Rönnber, I.; et al. Real-time biospecific interaction analysis using surface plasmon resonance and a sensor chip technology. BioTechniques 1991, 11, 620–627. [Google Scholar] [PubMed]
- Jason-Moller, L.; Murphy, M.; Bruno, J. Overview of Biacore systems and their applications. Curr. Protoc. Protein Sci. 2006, 19. [Google Scholar] [CrossRef]
- Navratilova, I.; Hopkins, A.L. Emerging role of surface plasmon resonance in fragment-based drug discovery. Future Med. Chem. 2011, 3, 1809–1820. [Google Scholar] [CrossRef] [PubMed]
- Rich, R.L.; Myszka, D.G. Spying on HIV with SPR. Trends Microbiol. 2003, 11, 124–133. [Google Scholar] [CrossRef]
- de Mol, N.J. Surface plasmon resonance for proteomics. Methods Mol. Biol. 2012, 800, 33–53. [Google Scholar] [PubMed]
- Fivash, M.; Towler, E.M.; Fisher, R.J. BIAcore for macromolecular interaction. Curr. Opin. Biotechnol. 1998, 9, 97–101. [Google Scholar] [CrossRef]
- Bergwerff, A.A.; van Knapen, F. Surface plasmon resonance biosensors for detection of pathogenic microorganisms: Strategies to secure food and environmental safety. J. AOAC. Int. 2006, 89, 826–831. [Google Scholar] [PubMed]
- Arima, Y.; Teramura, Y.; Takiguchi, H.; Kawano, K.; Kotera, H.; Iwata, H. Surface plasmon resonance and surface plasmon field-enhanced fluorescence spectroscopy for sensitive detection of tumor markers. Methods Mol. Biol. 2009, 503, 3–20. [Google Scholar] [PubMed]
- Thillaivinayagalingam, P.; Gommeaux, J.; McLoughlin, M.; Collins, D.; Newcombe, A.R. Biopharmaceutical production: Applications of surface plasmon resonance biosensors. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2010, 878, 149–153. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, J.M. Surface plasmon resonance: towards an understanding of the mechanisms of biological molecular recognition. Curr. Opin. Chem. Biol. 2001, 5, 572–577. [Google Scholar] [CrossRef]
- Xue, T.; Cui, X.; Guan, W.; Wang, Q.; Liu, C.; Wang, H.; Qi, K.; Singh, D.J.; Zheng, W. Surface plasmon resonance technique for directly probing the interaction of DNA and graphene oxide and ultra-sensitive biosensing. Biosens. Bioelectron. 2014, 58, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Wang, Z.; Guan, W.; Hou, C.; Shi, Z.; Zheng, W.; Cui, X. Investigating the interaction of dye molecules with graphene oxide by using a surface plasmon resonance technique. RSC Adv. 2014, 4, 50789–50794. [Google Scholar] [CrossRef]
- Yu, W.H.; Yu, S.; Meng, Q.; Brew, K.; Woessner, J.F., Jr. TIMP-3 binds to sulfated glycosaminoglycans of the extracellular matrix. J. Biol. Chem. 2000, 275, 31226–31232. [Google Scholar] [CrossRef] [PubMed]
- Andreú, T.; Beckers, T.; Thoenes, E.; Hilgard, P.; von Melchner, H. Gene trapping identifies inhibitors of oncogenic transformation. The tissue inhibitor of metalloproteinases-3 (TIMP3) and collagen type I alpha2 (COL1A2) are epidermal growth factor-regulated growth repressors. J. Biol. Chem. 1998, 273, 13848–13854. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Murate, T.; Hayakawa, T. Multiple functions of tissue inhibitors of metalloproteinases (TIMPs): new aspects in hematopoiesis. Platelets. 1999, 10, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Capila, I.; Linhardt, R.J. Heparin-protein interactions. Angew. Chem. Int. Ed. 2002, 41, 391–412. [Google Scholar] [CrossRef]
- Häcker, U.; Nybakken, K.; Perrimon, N. Heparan sulphate proteoglycans: the sweet side of development. Nat. Rev. Mol. Cell. Biol. 2005, 6, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Sasisekharan, R.; Raman, R.; Prabhakar, V. Glycomics approach to structure-function relationships of glycosaminoglycans. Annu. Rev. Biomed. Eng. 2006, 8, 181–231. [Google Scholar] [CrossRef] [PubMed]
- Ra, H.J.; Harju-Baker, S.; Zhang, F.; Linhardt, R.J.; Wilson, C.L.; Parks, W.C. Control of promatrilysin (MMP7) activation and substrate-specific activity by sulfated glycosaminoglycans. J. Biol. Chem. 2009, 284, 27924–27932. [Google Scholar] [CrossRef] [PubMed]
- Fulcher, Y.G.; Sanganna, G.R.R.; Frey, N.C.; Zhang, F.; Linhardt, R.J.; King, G.M.; Van Doren, S.R. Heparinoids activate a protease, secreted by mucosa and tumors, via tethering supplemented by allostery. ACS. Chem. Biol. 2014, 9, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Troeberg, L.; Lazenbatt, C.; Anower-E-Khuda, M.F.; Freeman, C.; Federov, O.; Habuchi, H.; Habuchi, O.; Kimata, K.; Nagase, H. Sulfated glycosaminoglycans control the extracellular trafficking and the activity of the metalloprotease inhibitor TIMP-3. Chem. Biol. 2014, 21, 1300–1309. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.E.; Buttle, D.J.; Short, R.D.; McArthur, S.L.; Steele, D.A.; Whittle, J.D. Glycosaminoglycan (GAG) binding surfaces for characterizing GAG-protein interactions. Biomaterials 2012, 33, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Yates, E.A.; Santini, F.; Guerrini, M.; Naggi, A.; Torri, G.; Casu, B. 1H and 13C NMR spectral assignments of the major sequences of twelve systematically modified heparin derivatives. Carbohydr. Res. 1996, 294, 15–27. [Google Scholar] [CrossRef]
- Wang, L.; Brown, J.R.; Varki, A.; Esko, J.D. Heparin’s anti-inflammatory effects require glucosamine 6-O-sulfation and are mediated by blockade of L- and P-selectins. J. Clin. Invest. 2002, 110, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Hernaiz, M.; Liu, J.; Rosenberg, R.D.; Linhardt, R.J. Enzymatic modification of heparan sulfate on a biochip promotes its interaction with antithrombin III. Biochem. Biophys. Res. Commun. 2000, 276, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; McLellan, J.S.; Ayala, A.M.; Leahy, D.J.; Linhardt, R.J. Kinetic and structural studies on interactions between heparin or heparan sulfate and proteins of the hedgehog signaling pathway. Biochemistry 2007, 46, 3933–3941. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liang, X.; Pu, D.; George, K.I.; Holland, P.J.; Walsh, S.T.; Linhardt, R.J. Biophysical characterization of glycosaminoglycan-IL-7 interactions using SPR. Biochimie 2012, 94, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Bode, W.; Fernandez-Catalan, C.; Grams, F.; Gomis-Rüth, F.X.; Nagase, H.; Tschesche, H.; Maskos, K. Insights into MMP-TIMP interactions. Ann. N. Y. Acad. Sci. 1999, 878, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Batra, J.; Robinson, J.; Soares, A.S.; Fields, A.P.; Radisky, D.C.; Radisky, E.S. Matrix metalloproteinase-10 (MMP-10) interaction with tissue inhibitors of metalloproteinases TIMP-1 and TIMP-2: binding studies and crystal structure. J. Biol. Chem. 2012, 287, 15935–15946. [Google Scholar] [CrossRef] [PubMed]
- Tocchi, A.; Parks, W.C. Functional interactions between matrix metalloproteinases and glycosaminoglycans. FEBS. J. 2013, 280, 2332–2341. [Google Scholar] [CrossRef] [PubMed]
- Brew, K.; Nagase, H. The tissue inhibitors of metalloproteinases (TIMPs): An ancient family with structural and functional diversity. Biochim. Biophys. Acta. 2010, 1803, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Amour, A.; Slocombe, P.M.; Webster, A.; Butler, M.; Knight, C.G.; Smith, B.J.; Stephens, P.E.; Shelley, C.; Hutton, M.; Knäuper, V.; Docherty, A.J.P.; Murphy, G. TNF-Alpha converting enzyme (TACE) is inhibited by TIMP-3. FEBS Lett. 1998, 435, 39–44. [Google Scholar] [CrossRef]
- Wayne, G.J.; Deng, S.J.; Amour, A.; Borman, S.; Matico, R.; Carter, H.L.; Murphy, G. TIMP-3 inhibition of ADAMTS-4 (Aggrecanase-1) is regulated by interactions between aggrecan and the C-terminal domain of ADAMTS-4. J. Biol. Chem. 2007, 282, 20991–20998. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.M.; Ge, G.; Lim, N.H.; Nagase, H.; Greenspan, D.S. TIMP-3 inhibits the procollagen N-proteinase ADAMTS-2. Biochem. J. 2006, 398, 515–519. [Google Scholar] [PubMed]
- Troeberg, L.; Fushimi, K.; Khokha, R.; Emonard, H.; Ghosh, P.; Nagase, H. Calcium pentosan polysulfate is a multifaceted exosite inhibitor of aggrecanases. FASEB J. 2008, 22, 3515–3524. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Lee, K.B.; Linhardt, R.J. SPR Biosensor Probing the Interactions between TIMP-3 and Heparin/GAGs. Biosensors 2015, 5, 500-512. https://doi.org/10.3390/bios5030500
Zhang F, Lee KB, Linhardt RJ. SPR Biosensor Probing the Interactions between TIMP-3 and Heparin/GAGs. Biosensors. 2015; 5(3):500-512. https://doi.org/10.3390/bios5030500
Chicago/Turabian StyleZhang, Fuming, Kyung Bok Lee, and Robert J. Linhardt. 2015. "SPR Biosensor Probing the Interactions between TIMP-3 and Heparin/GAGs" Biosensors 5, no. 3: 500-512. https://doi.org/10.3390/bios5030500
APA StyleZhang, F., Lee, K. B., & Linhardt, R. J. (2015). SPR Biosensor Probing the Interactions between TIMP-3 and Heparin/GAGs. Biosensors, 5(3), 500-512. https://doi.org/10.3390/bios5030500