Highly Sensitive Whole-Cell Mercury Biosensors for Environmental Monitoring

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, Media, and Reagents
2.2. Plasmid Circuit Construction
2.3. Reporter Expression Assay
2.4. Environmental Samples from Madre de Dios, Peru
2.5. Data Analysis
2.5.1. Relative Concentration of the Reporter Protein
2.5.2. Promoter Activity
2.5.3. Dose–Response Curves
2.5.4. Calculation of Linear Range
2.5.5. Calculation of Limit of Detection (LOD)
2.5.6. Bacterial Growth
3. Results
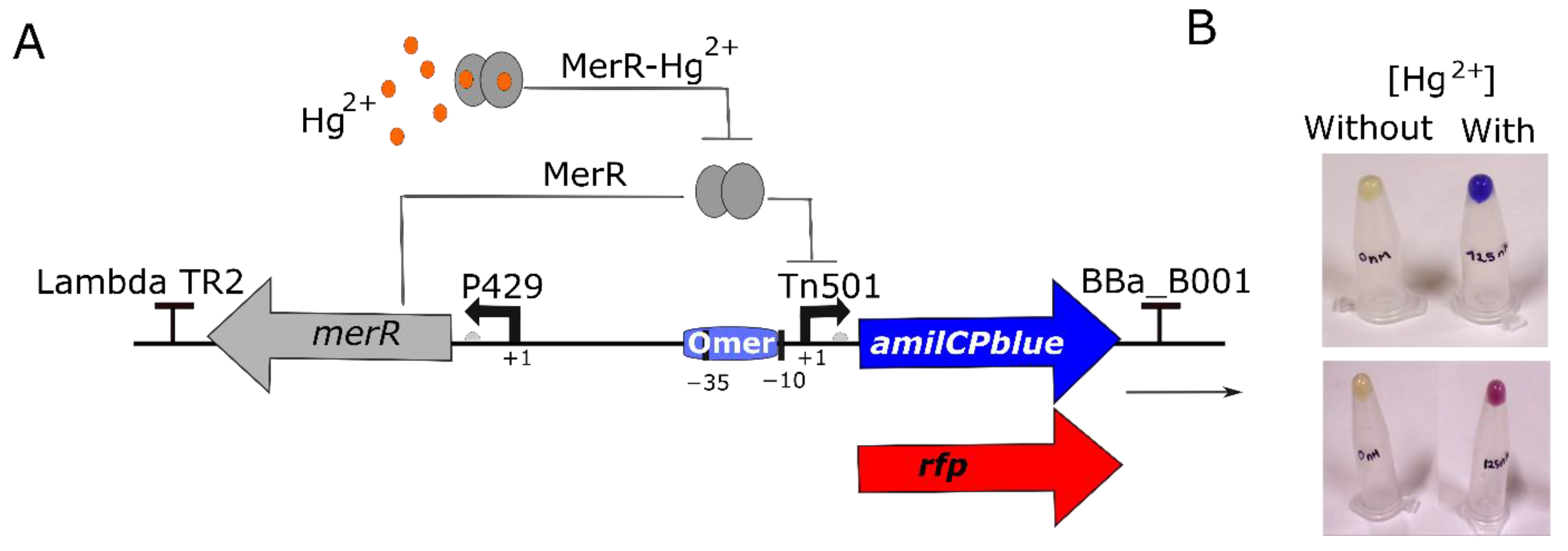
3.1. Design of a Synthetic Genetic Circuit for Mercury Detection
3.2. Testing the Biosensor Circuit
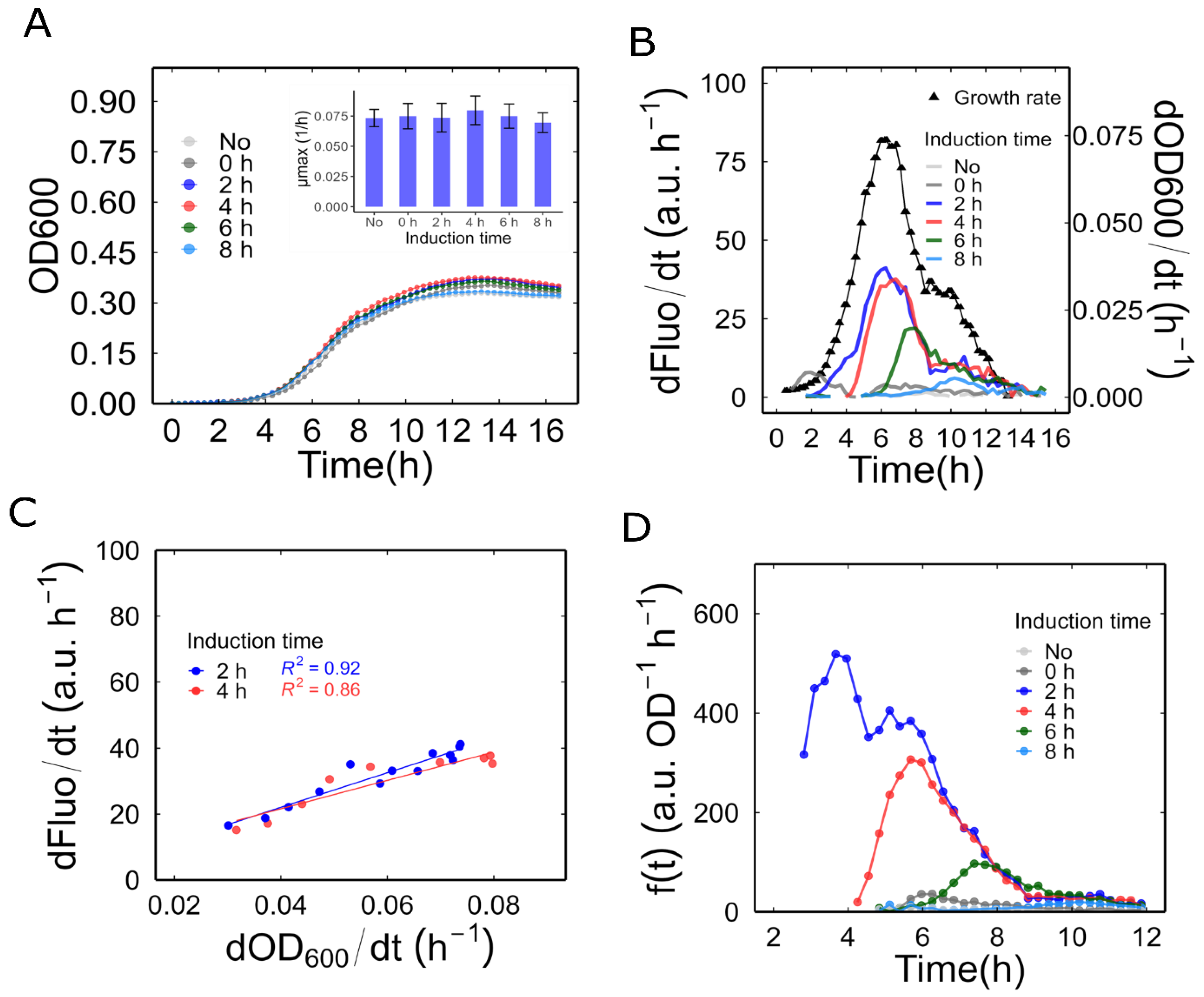
3.3. Reporter Expression Induced at Various Phases of Bacterial Growth
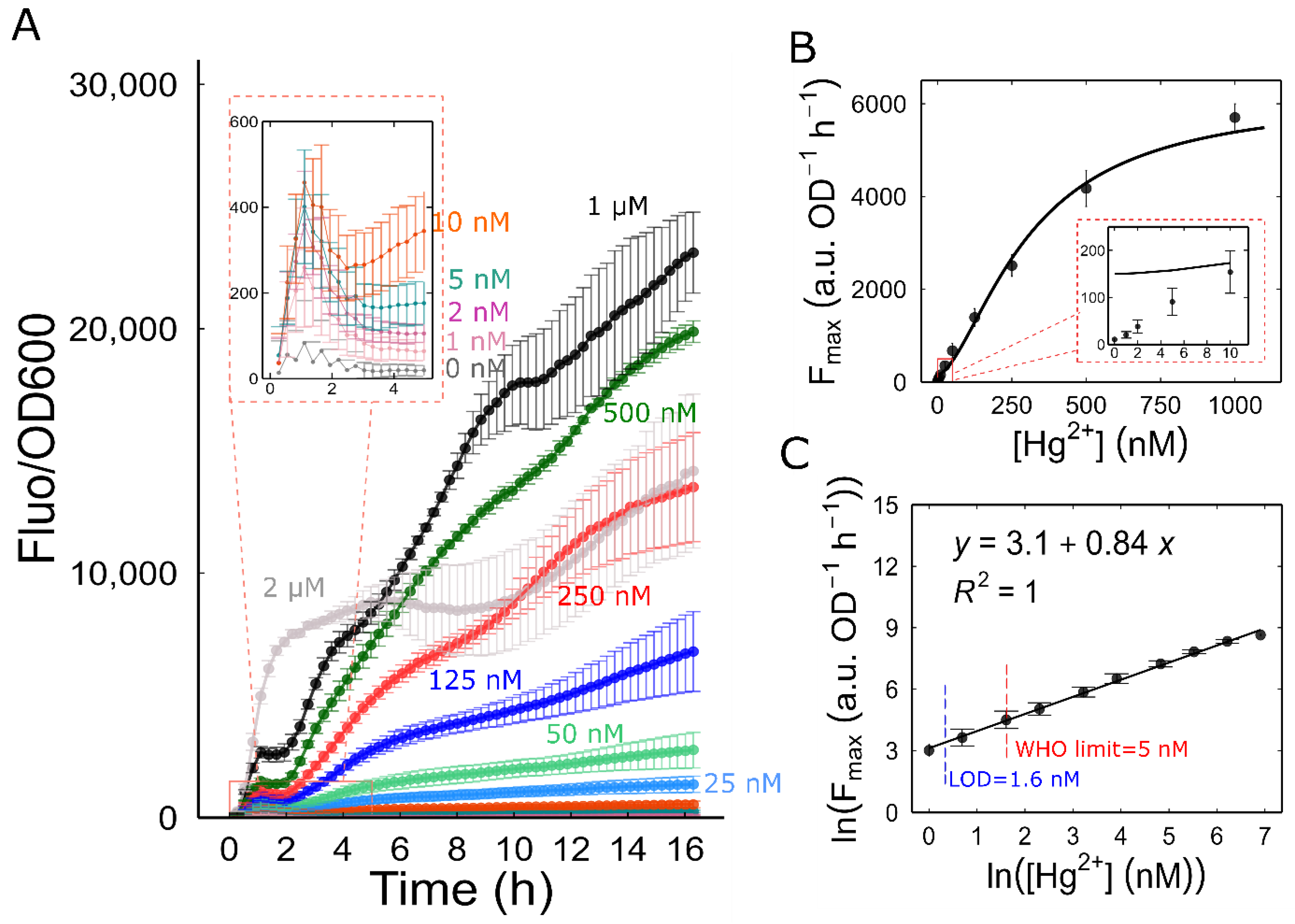
3.4. Characterization of Mer-RFP
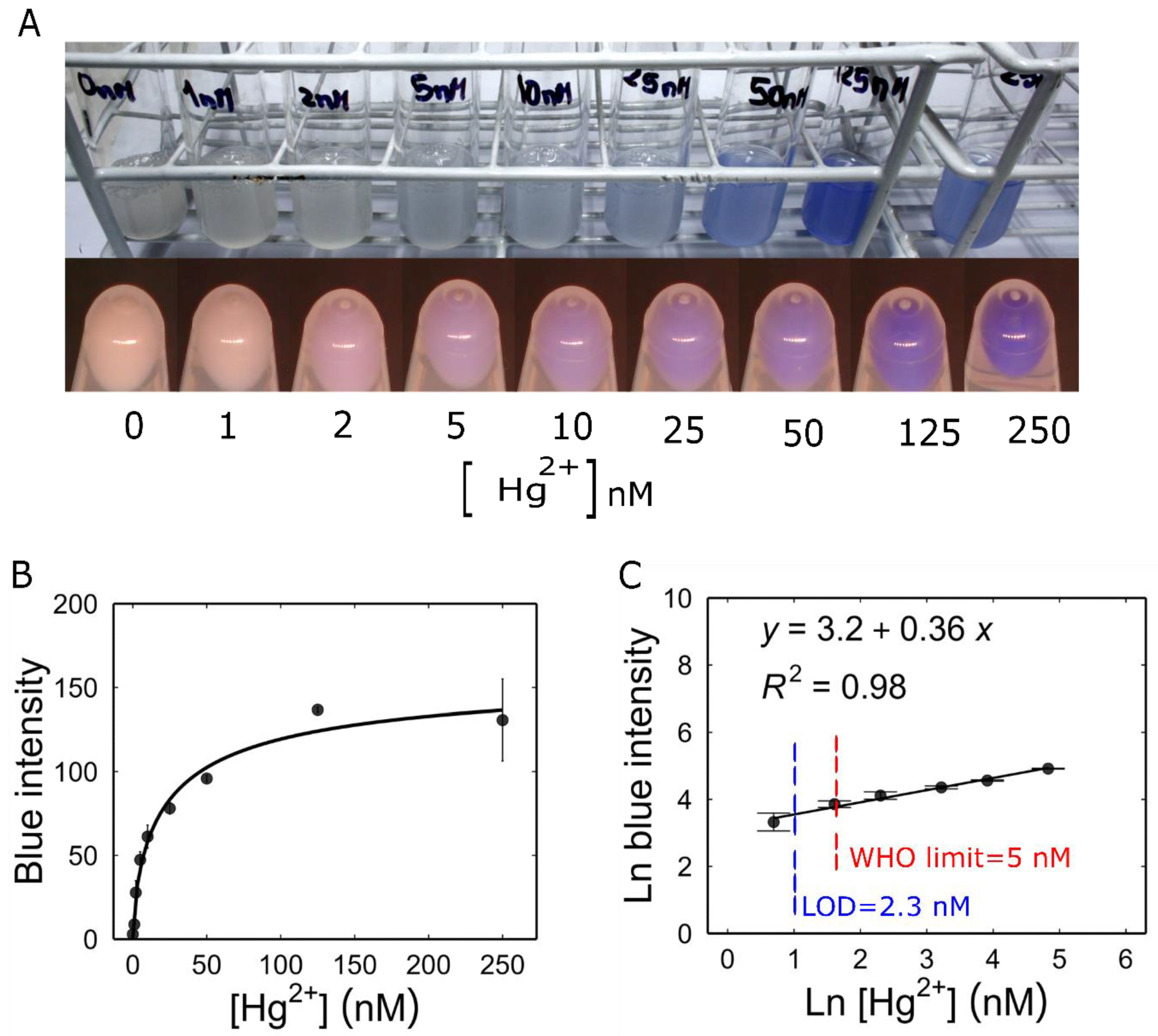
3.5. Characterization of Mer-Blue
3.6. Analysis of Environment Sample from Madre de Dios, Peru
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lamborg, C.; Bowman, K.; Hammerschmidt, C.; Gilmour, C.; Munson, K.; Selin, N.; Tseng, C.-M. Mercury in the Anthropocene Ocean. Oceanog 2014, 27, 76–87. [Google Scholar] [CrossRef]
- Li, F.; Ma, C.; Zhang, P. Mercury Deposition, Climate Change and Anthropogenic Activities: A Review. Front. Earth Sci. 2020, 8, 316. [Google Scholar] [CrossRef]
- Gautam, R.K.; Sharma, S.K.; Mahiya, S.; Chattopadhyaya, M.C. CHAPTER 1. Contamination of Heavy Metals in Aquatic Media: Transport, Toxicity and Technologies for Remediation. In Heavy Metals in Water; Royal Society of Chemistry: Washington, DC, USA, 2014; pp. 1–24. [Google Scholar]
- Rana, M.N.; Tangpong, J.; Rahman, M.M. Toxicodynamics of Lead, Cadmium, Mercury and Arsenic- induced kidney toxicity and treatment strategy: A mini review. Toxicol. Rep. 2018, 5, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, Y.; Wang, F.; Luo, Z.; Guo, S.; Strähle, U. Toxicity of mercury: Molecular evidence. Chemosphere 2020, 245, 125586. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Driscoll, C.T.; Eagles-Smith, C.A.; Eckley, C.S.; Gay, D.A.; Hsu-Kim, H.; Keane, S.E.; Kirk, J.L.; Mason, R.P.; Obrist, D.; et al. A Critical Time for Mercury Science to Inform Global Policy. Environ. Sci. Technol. 2018, 52, 9556–9561. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Shi, Z.; Long, Z.; Wu, P.; Zheng, C.; Hou, X. Determination and speciation of mercury in environmental and biological samples by analytical atomic spectrometry. Microchem. J. 2012, 103, 1–14. [Google Scholar] [CrossRef]
- Wyatt, L.; Ortiz, E.J.; Feingold, B.; Berky, A.; Diringer, S.; Morales, A.M.; Jurado, E.R.; Hsu-Kim, H.; Pan, W. Spatial, temporal, and dietary variables associated with elevated mercury exposure in peruvian riverine communities upstream and downstream of artisanal and small-scale gold mining. Int. J. Environ. Res. Public Health 2017, 14, 1582. [Google Scholar] [CrossRef] [PubMed]
- Lund, P.A.; Ford, S.J.; Brown, N.L. Transcriptional regulation of the mercury-resistance genes of transposon Tn501. J. Gen. Microbiol. 1986, 132, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Parkhill, J.; Brown, N.L. Site-specific insertion and deletion mutants in the mer promoter-operator region of Tn501; the nineteen base-pair spacer is essential for normal induction of the promoter by MerR. Nucleic Acids Res. 1990, 18, 5157–5162. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC332137/ (accessed on 20 February 2024). [CrossRef]
- Wang, D.; Huang, S.; Liu, P.; Liu, X.; He, Y.; Chen, W.; Hu, Q.; Wei, T.; Gan, J.; Ma, J.; et al. Structural Analysis of the Hg(II)-Regulatory Protein Tn501 MerR from Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 33391. [Google Scholar] [CrossRef]
- O’Halloran, T.V.; Frantz, B.; Shin, M.K.; Ralston, D.M.; Wright, J.G. The MerR heavy metal receptor mediates positive activation in a topologically novel transcription complex. Cell 1989, 56, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.L.; Stoyanov, J.V.; Kidd, S.P.; Hobman, J.L. The MerR family of transcriptional regulators. FEMS Microbiol. Rev. 2003, 27, 145–163. [Google Scholar] [CrossRef] [PubMed]
- Ivask, A.; Hakkila, K.; Virta, M. Detection of organomercurials with sensor bacteria. Anal. Chem. 2001, 73, 5168–5171. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.H.; Sørensen, S.J. Versatile biosensor vectors for detection and quantification of mercury. FEMS Microbiol. Lett. 2000, 193, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Du, R.; Xie, Z.; He, X.; Huang, K.; Luo, Y.; Xu, W. Using the promoters of MerR family proteins as “rheostats” to engineer whole-cell heavy metal biosensors with adjustable sensitivity. J. Biol. Eng. 2019, 13, 70. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, X.; Su, H.; Guo, M.; Liu, H. Advances in Synthetic-Biology-Based Whole-Cell Biosensors: Principles, Genetic Modules, and Applications in Food Safety. Int. J. Mol. Sci. 2023, 24, 7989. [Google Scholar] [CrossRef] [PubMed]
- Stocker, J.; Balluch, D.; Gsell, M.; Harms, H.; Feliciano, J.; Daunert, S.; Malik, K.A.; van der Meer, J.R. Development of a Set of Simple Bacterial Biosensors for Quantitative and Rapid Measurements of Arsenite and Arsenate in Potable Water. Environ. Sci. Technol. 2003, 37, 4743–4750. [Google Scholar] [CrossRef]
- Gui, Q.; Lawson, T.; Shan, S.; Yan, L.; Liu, Y. The Application of Whole Cell-Based Biosensors for Use in Environmental Analysis and in Medical Diagnostics. Sensors 2017, 17, 1623. [Google Scholar] [CrossRef]
- Lopreside, A.; Wan, X.; Michelini, E.; Roda, A.; Wang, B. Comprehensive Profiling of Diverse Genetic Reporters with Application to Whole-Cell and Cell-Free Biosensors. Anal. Chem. 2019, 91, 15284–15292. [Google Scholar] [CrossRef]
- fcoppens Deliberate Release of GMOs’, Belgian Biosafety Server. Available online: https://www.biosafety.be/content/deliberate-release-gmos (accessed on 26 April 2024).
- Wan, X.; Volpetti, F.; Petrova, E.; French, C.; Maerkl, S.J.; Wang, B. Cascaded amplifying circuits enable ultrasensitive cellular sensors for toxic metals. Nat. Chem. Biol. 2019, 15, 540–548. [Google Scholar] [CrossRef]
- Scott, M.; Gunderson, C.W.; Mateescu, E.M.; Zhang, Z.; Hwa, T. Interdependence of cell growth and gene expression: Origins and consequences. Science 2010, 330, 1099–1102. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.H.; Caputo, A.T.; French, N.G.; Peat, T.S.; Whitfield, J.; Warden, A.C.; Newman, J.; Scott, C. Over the rainbow: Structural characterization of the chromoproteins gfasPurple, amilCP, spisPink and eforRed. Acta Crystallogr. D Struct. Biol. 2022, 78, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.E.; Tour, O.; Palmer, A.E.; Steinbach, P.A.; Baird, G.S.; Zacharias, D.A.; Tsien, R.Y. A monomeric red fluorescent protein. Proc. Natl. Acad. Sci. USA 2002, 99, 7877–7882. [Google Scholar] [CrossRef] [PubMed]
- Liljeruhm, J.; Funk, S.K.; Tietscher, S.; Edlund, A.D.; Jamal, S.; Wistrand-Yuen, P.; Dyrhage, K.; Gynnå, A.; Ivermark, K.; Lövgren, J.; et al. Engineering a palette of eukaryotic chromoproteins for bacterial synthetic biology. J. Biol. Eng. 2018, 12, 8. [Google Scholar] [CrossRef]
- Boyer, F.; Besson, B.; Baptist, G.; Izard, J.; Pinel, C.; Ropers, D.; Geiselmann, J.; Jong, H.D.; Cnrs, M.; Fourier, U.J.; et al. WellReader: A MATLAB program for the analysis of fluorescence and luminescence reporter gene data. Bioinformatics 2010, 26, 1262–1263. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.K.; Guzman, C.D.; Taylor, N.D.; Raman, S.; Anderson, K.; Church, G.M. Synthetic biosensors for precise gene control and real-time monitoring of metabolites. Nucleic Acids Res. 2015, 43, 7648–7660. [Google Scholar] [CrossRef]
- Du, R.; Guo, M.; He, X.; Huang, K.; Luo, Y.; Xu, W. Feedback regulation mode of gene circuits directly affects the detection range and sensitivity of lead and mercury microbial biosensors. Anal. Chim. Acta 2019, 1084, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Prabowo, B.A.; Cabral, P.D.; Freitas, P.; Fernandes, E. The Challenges of Developing Biosensors for Clinical Assessment: A Review. Chemosensors 2021, 9, 299. [Google Scholar] [CrossRef]
- Wang, D.; Zheng, Y.; Fan, X.; Xu, L.; Pang, T.; Liu, T.; Liang, L.; Huang, S.; Xiao, Q.; Al, W.E.T.; et al. Visual detection of Hg2+ by manipulation of pyocyanin biosynthesis through the Hg2+ dependent transcriptional activator MerR in microbial cells. J. Biosci. Bioeng. 2020, 129, 223–228. [Google Scholar] [CrossRef]
- Brophy, J.A.N.; Voigt, C.A. Principles of genetic circuit design. Nat. Methods 2014, 11, 508–520. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO) and World Health Organization (WHO). General Standard for Contaminants and Toxins in Food and FeedCodex Alimentariusen.pdf. International Food Standards. 2019. Available online: https://www.fao.org/fao-who-codexalimentarius/sh-proxy/en/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FStandards%252FCXS%2B193-1995%252FCXS_193e.pdf (accessed on 10 March 2022).
- Wang, D.; Zheng, Y.; Xu, L.; Fan, X.; Wei, N.; Jin, N.; Huang, S.; Xiao, Q.; Wu, Z. Engineered cells for selective detection and remediation of Hg2+ based on transcription factor MerR regulated cell surface displayed systems. Biochem. Eng. J. 2019, 150, 107289. [Google Scholar] [CrossRef]
- Guo, Y.; Hui, C.y.; Liu, L.; Chen, M.P.; Huang, H.Y. Development of a bioavailable Hg(II) sensing system based on MerR-regulated visual pigment biosynthesis. Sci. Rep. 2021, 11, 13516. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Shen, Y.; Zou, Y.; Sun, P.; Wei, W.; Zhao, J.; Zhang, C. Engineering highly sensitive whole-cell mercury biosensors based on positive feedback loops from quorum-sensing systems. Analyst 2018, 143, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Din, G.; Hasan, F.; Conway, M.; Denney, B.; Ripp, S.; Shah, A.A. Engineering a bioluminescent bioreporter from an environmentally sourced mercury-resistant Enterobacter cloacae strain for the detection of bioavailable mercury. J. Appl. Microbiol. 2019, 127, 1125–1134. [Google Scholar] [CrossRef]
- Guo, M.; Wang, J.; Du, R.; Liu, Y.; Chi, J.; He, X.; Huang, K.; Luo, Y.; Xu, W. A test strip platform based on a whole-cell microbial biosensor for simultaneous on-site detection of total inorganic mercury pollutants in cosmetics without the need for predigestion. Biosens. Bioelectron. 2020, 150, 111899. [Google Scholar] [CrossRef]
- Mahbub, K.R.; Krishnan, K.; Naidu, R.; Megharaj, M. Development of a whole cell biosensor for the detection of inorganic mercury. Environ. Technol. Innov. 2017, 8, 64–70. [Google Scholar] [CrossRef]
- Priyadarshi, H.; Alam, A.; Gireesh-Babu, P.; Das, R.; Kishore, P.; Kumar, S.; Chaudhari, A. A GFP-based bacterial biosensor with chromosomally integrated sensing cassette for quantitative detection of Hg(II) in environment. J. Environ. Sci. China 2012, 24, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Cheng, H.; Ting, M.; Wen-Hui, Z.; Xian-Gui, L. A chromosomally based luminescent bioassay for mercury detection in red soil of China. Appl. Microbiol. Biotechnol. 2010, 87, 981–989. [Google Scholar] [CrossRef]
- Zhang, N.-X.; Guo, Y.; Li, H.; Yang, X.-Q.; Gao, C.-X.; Hui, C.-Y. Versatile artificial mer operons in Escherichia coli towards whole cell biosensing and adsorption of mercury. PLoS ONE 2021, 16, e0252190. [Google Scholar] [CrossRef]
- Lee, K.H.; Kim, D.M. In vitro use of cellular synthetic machinery for biosensing applications. Front. Pharmacol. 2019, 10, 455867. [Google Scholar] [CrossRef]
- Noireaux, V.; Liu, A.P. The New Age of Cell-Free Biology. Annu. Rev. Biomed. Eng. 2020, 22, 51–77. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Chavez, F.; Arce, A.; Adhikari, A.; Vadhin, S.; Pedroza-Garcia, J.A.; Gandini, C.; Ajioka, J.W.; Molloy, J.; Sanchez-Nieto, S.; Varner, J.D.; et al. Constructing Cell-Free Expression Systems for Low-Cost Access. ACS Synth. Biol. 2022, 11, 1114–1128. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, J.P.; Tamar, B. Cracking the Mercury Methylation Code. Science 2013, 339, 1280–1281. [Google Scholar]
- Diringer, S.E.; Feingold, B.J.; Ortiz, E.J.; Gallis, J.A.; Araújo-Flores, J.M.; Berky, A.; Pan, W.K.Y.; Hsu-Kim, H. River transport of mercury from artisanal and small-scale gold mining and risks for dietary mercury exposure in Madre de Dios, Peru. Environ. Sci. Process. Impacts 2015, 17, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Griffin, H.G.; Foster, T.J.; Silver, S.; Misra, T.K. Cloning and DNA sequence of the mercuric- and organomercurial-resistance determinants of plasmid pDU1358. Proc. Natl. Acad. Sci. USA 1987, 84, 3112–3116. [Google Scholar] [CrossRef]
- Nucifora, G.; Chu, L.; Silver, S.; Misra, T.K. Mercury operon regulation by the merR gene of the organomercurial resistance system of plasmid pDU1358. J. Bacteriol. 1989, 171, 4241–4247. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC210196/ (accessed on 12 March 2024). [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SF-ICP-MS | Mer-RFP | Mer-Blue | ||||||
|---|---|---|---|---|---|---|---|---|
| Place | Status | Type of Sample | Std. Dev. | Std. Dev. | [Hg] nM | Std. Dev. | [Hg] nM | Std. Dev. |
| Laberinto district | Active Pond | Water | 0.48 | ±0.12 | <3.2 * | <4.6 * | ||
| Water–Sediment | 2.72 | ±0.31 | <3.2 * | <4.6 * | ||||
| Inactive pond | Water | 0.68 | ±0.18 | <3.2 * | <4.6 * | |||
| Water–Sediment | 0.62 | ±0.04 | <3.2 * | <4.6 * | ||||
| Isla de los Monos | Active pond | Water | 0.60 | ±0.24 | <3.2 * | <4.6 * | ||
| Water–Sediment | 2.37 | ±0.05 | <3.2 * | <4.6 * | ||||
| Reaction bucket | Recently mixed | Water | 25.13 | ±0.38 | 52.3 * | ±11.4 | <45.9 * | ±4.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zevallos-Aliaga, D.; De Graeve, S.; Obando-Chávez, P.; Vaccari, N.A.; Gao, Y.; Peeters, T.; Guerra, D.G. Highly Sensitive Whole-Cell Mercury Biosensors for Environmental Monitoring. Biosensors 2024, 14, 246. https://doi.org/10.3390/bios14050246
Zevallos-Aliaga D, De Graeve S, Obando-Chávez P, Vaccari NA, Gao Y, Peeters T, Guerra DG. Highly Sensitive Whole-Cell Mercury Biosensors for Environmental Monitoring. Biosensors. 2024; 14(5):246. https://doi.org/10.3390/bios14050246
Chicago/Turabian StyleZevallos-Aliaga, Dahlin, Stijn De Graeve, Pamela Obando-Chávez, Nicolás A. Vaccari, Yue Gao, Tom Peeters, and Daniel G. Guerra. 2024. "Highly Sensitive Whole-Cell Mercury Biosensors for Environmental Monitoring" Biosensors 14, no. 5: 246. https://doi.org/10.3390/bios14050246
APA StyleZevallos-Aliaga, D., De Graeve, S., Obando-Chávez, P., Vaccari, N. A., Gao, Y., Peeters, T., & Guerra, D. G. (2024). Highly Sensitive Whole-Cell Mercury Biosensors for Environmental Monitoring. Biosensors, 14(5), 246. https://doi.org/10.3390/bios14050246