Non-Invasive On–Off Fluorescent Biosensor for Endothelial Cell Detection


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
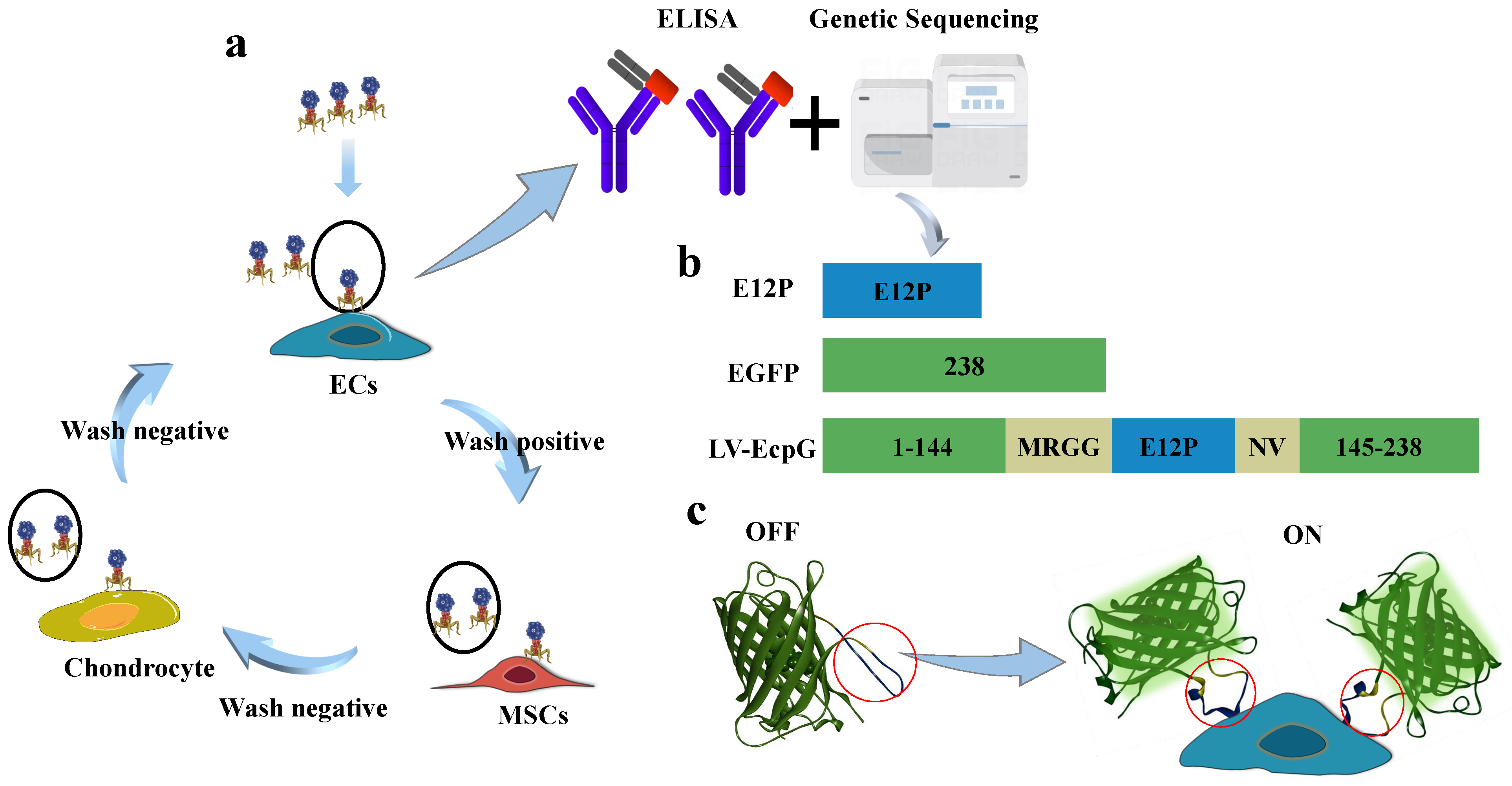
2.1. Phage Display Biopanning for Affinity Peptide Selection
2.2. ELISA
2.3. Plasmid Construction and Protein Expression
2.4. Cell Culture and Cell Differentiation
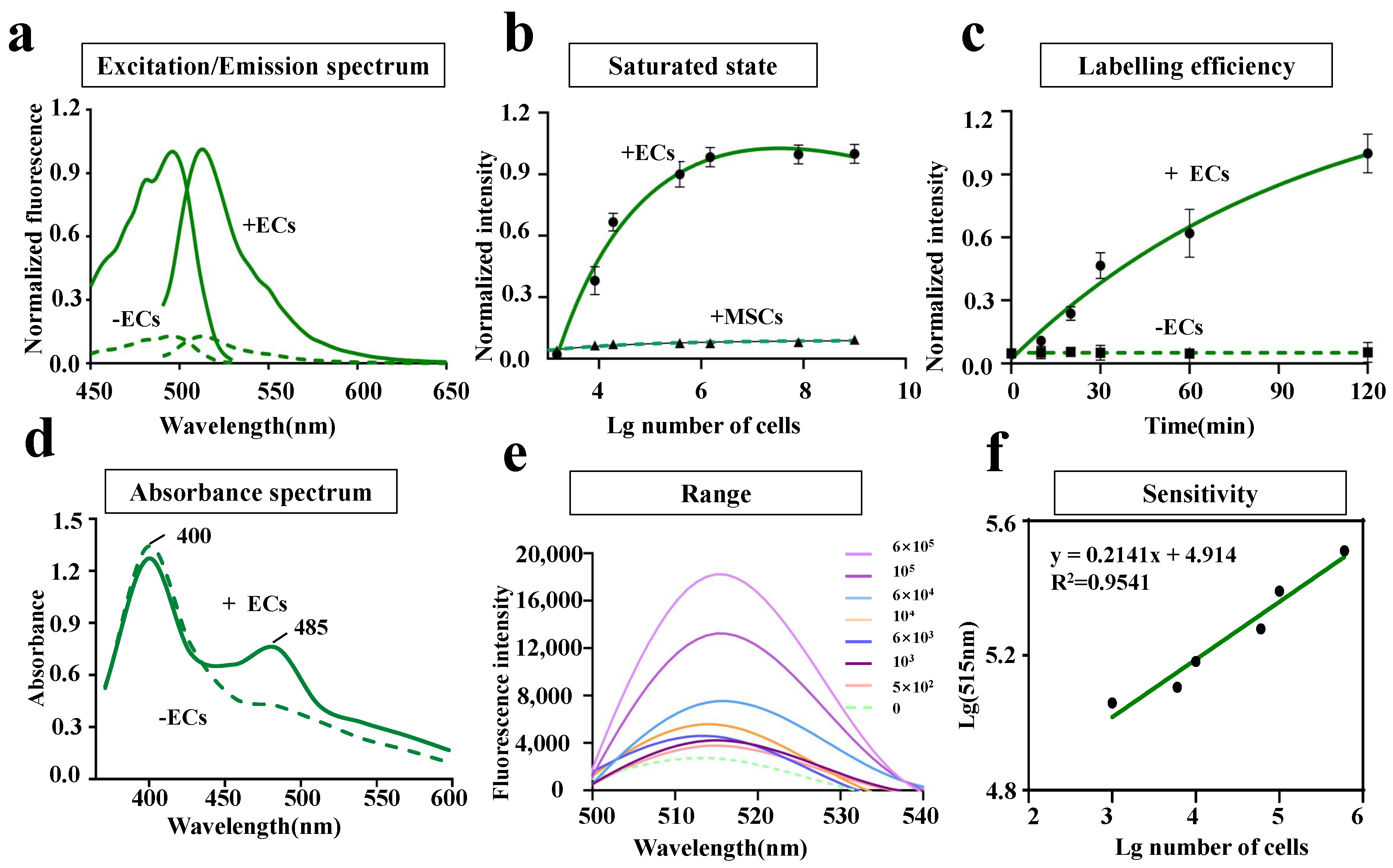
2.5. In Vitro Characterization of LV-EcpG Biosensor
2.6. Immunofluorescence
2.7. Confocal Microscopy
2.8. Flow Cytometry Analysis
3. Results and Discussion
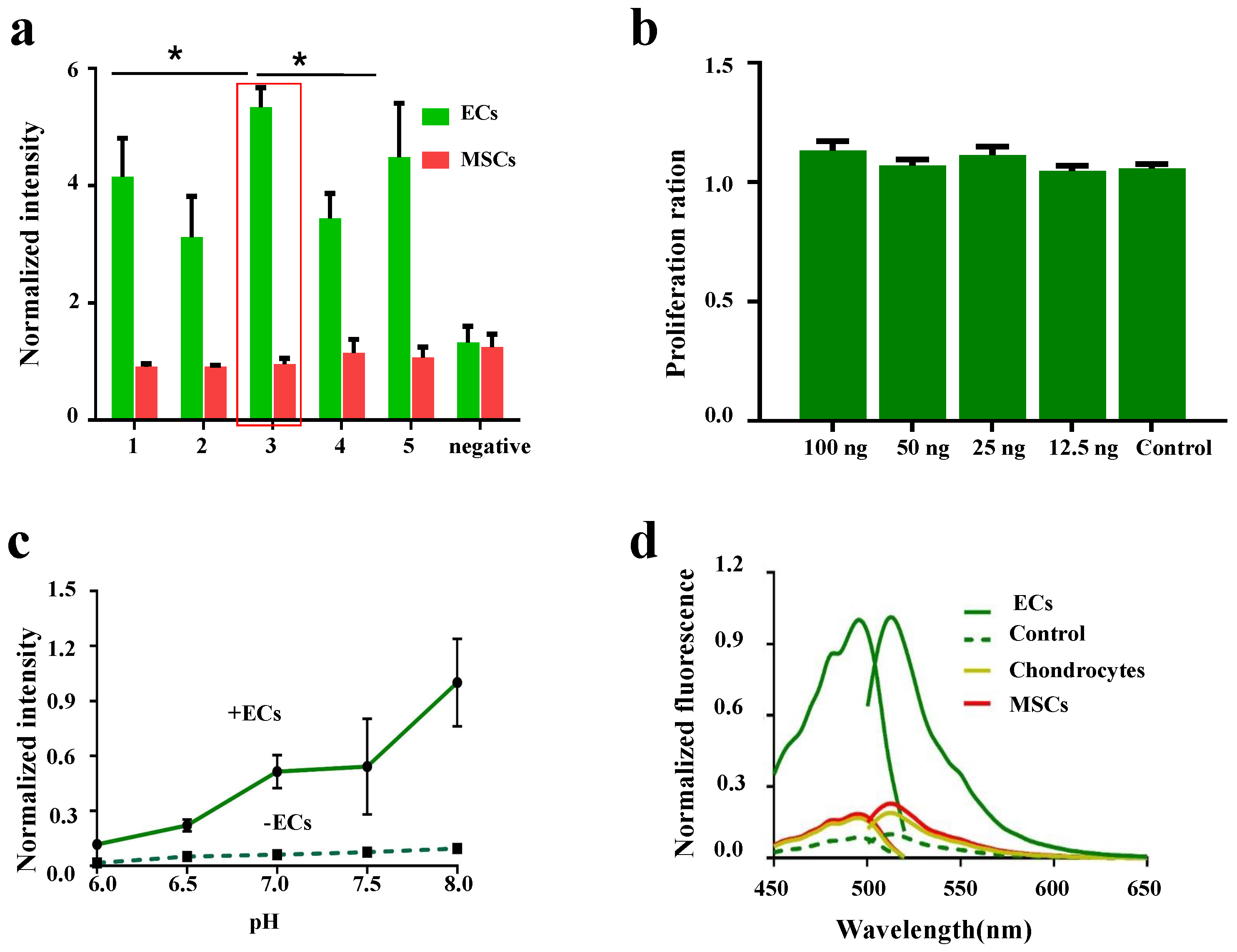
3.1. Design Strategies for LV-EcpG Biosensor
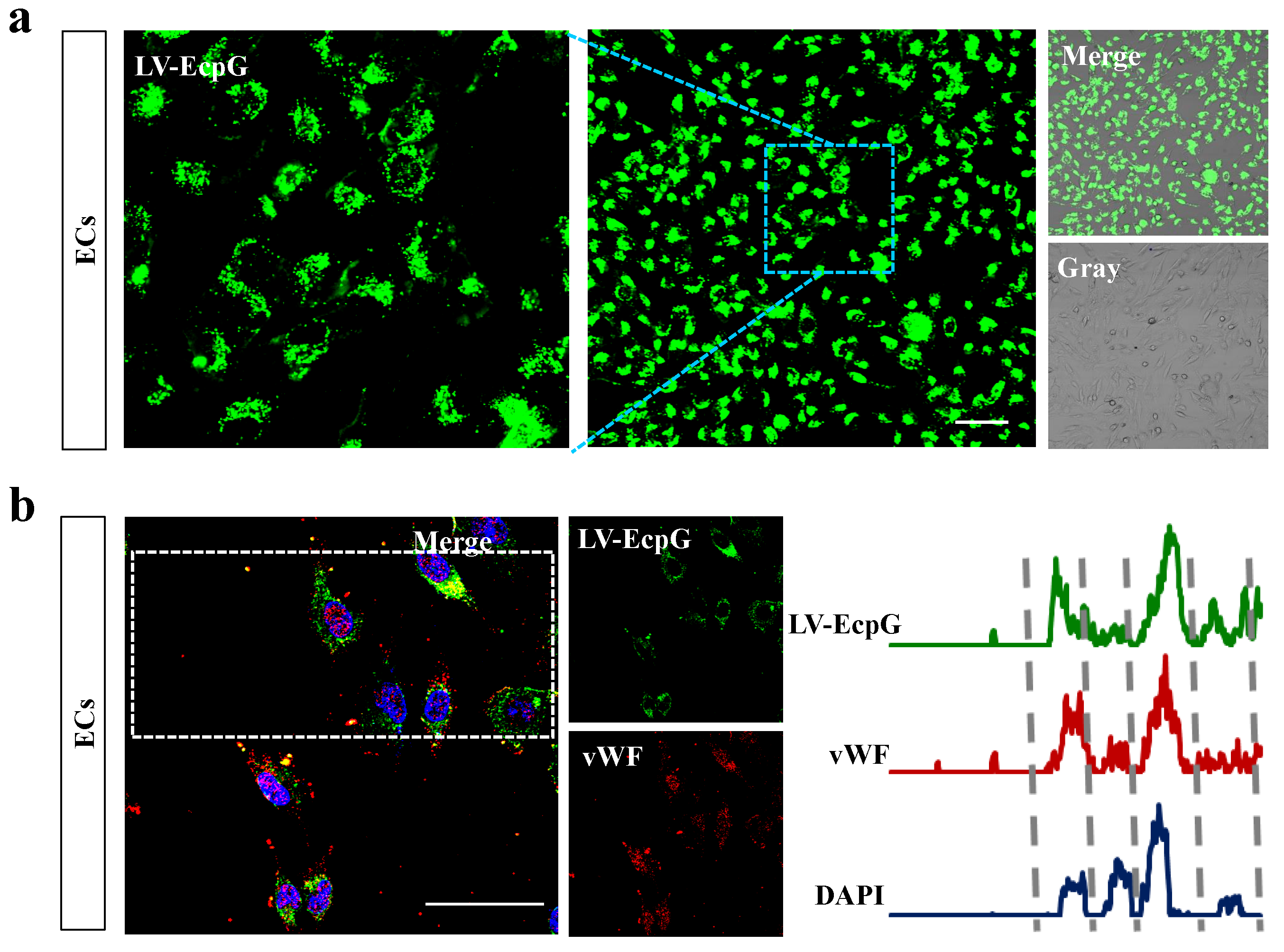
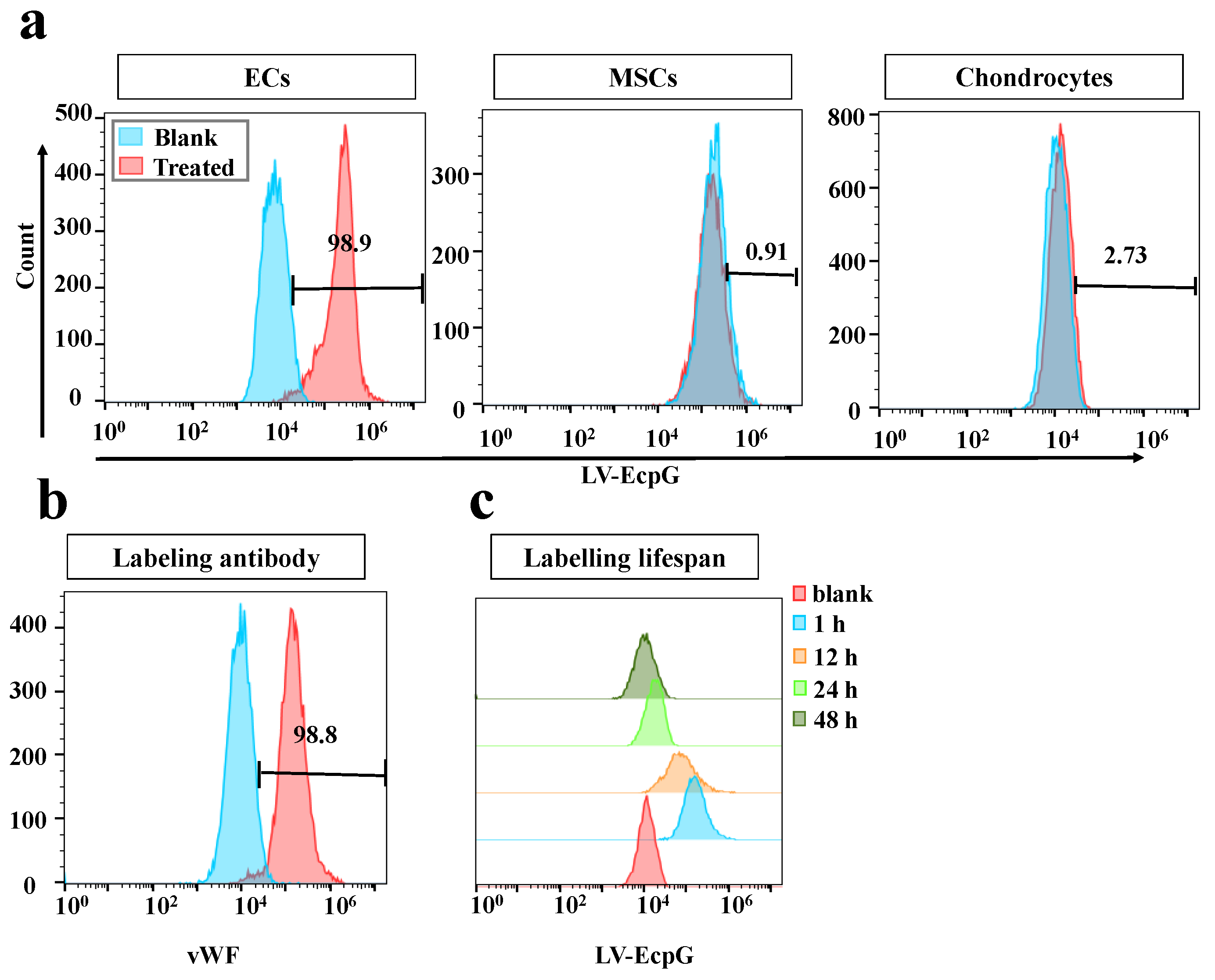
3.2. Characterization of LV-EcpG Biosensor in Cultured Cells
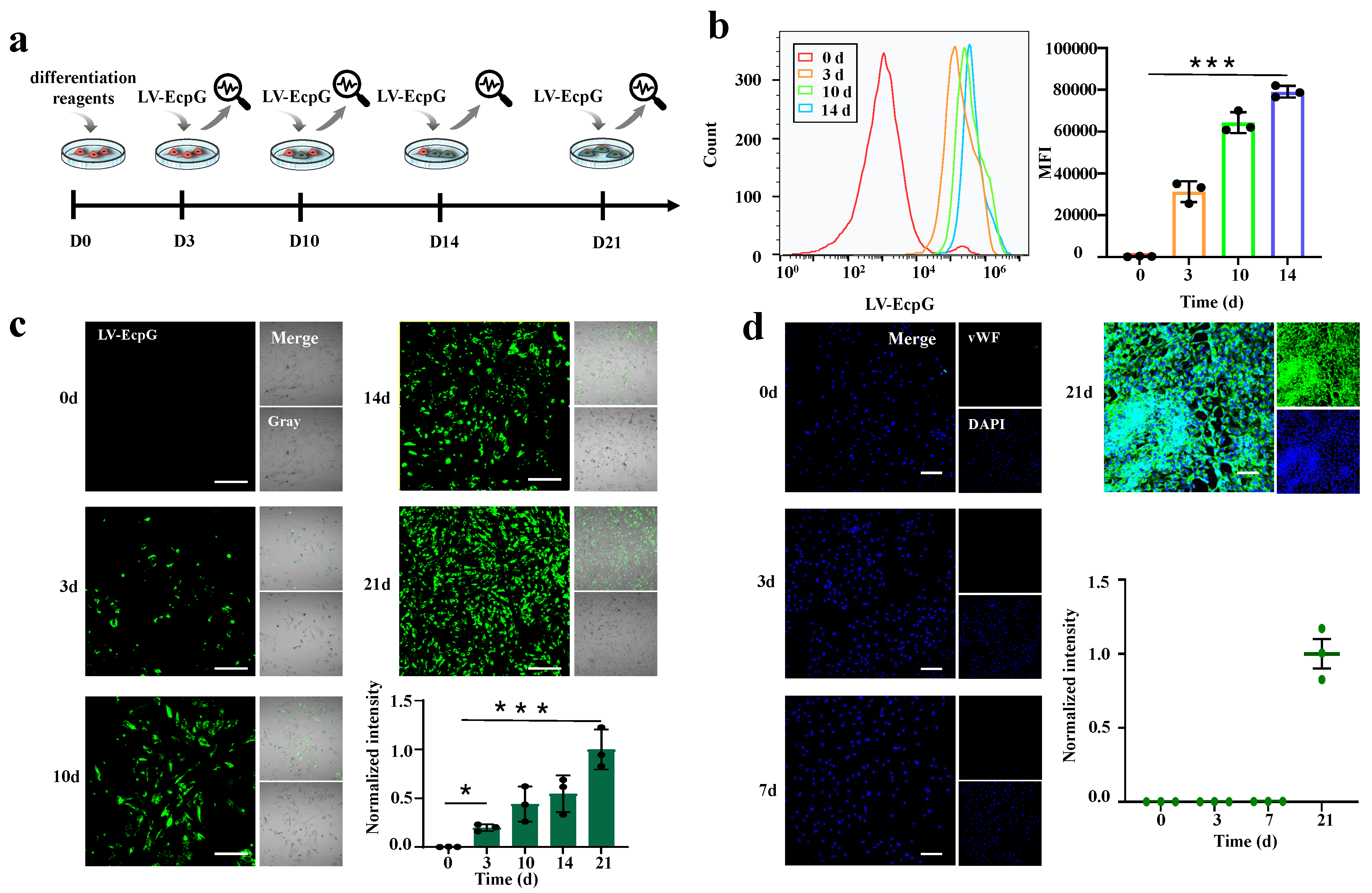
3.3. Visualization of the Directed MSC Differentiation Process into ECs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Xing, L.; Huang, G.; Chen, R.; Huang, L.; Liu, J.; Ren, X.; Wang, S.; Kuang, H.; Kumar, A.; Kim, J.K.; et al. Critical role of mitogen-inducible gene 6 in restraining endothelial cell permeability to maintain vascular homeostasis. J. Cell Commun. Signal. 2023, 17, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Amersfoort, J.; Eelen, G.; Carmeliet, P. Immunomodulation by endothelial cells—Partnering up with the immune system? Nat. Rev. Immunol. 2022, 22, 576–588. [Google Scholar] [CrossRef]
- Xu, S.; Ilyas, I.; Little, P.J.; Li, H.; Kamato, D.; Zheng, X.; Luo, S.; Li, Z.; Liu, P.; Han, J.; et al. Endothelial dysfunction in atherosclerotic cardiovascular diseases and beyond: From mechanism to pharmacotherapies. Pharmacol. Rev. 2021, 73, 924–967. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, X.; Kong, M.; Chen, S.; Bao, J.; Ji, Y. Three-dimensional microtumor formation of infantile hemangioma-derived endothelial cells for mechanistic exploration and drug screening. Pharmaceuticals 2022, 15, 1393. [Google Scholar] [CrossRef]
- Zakharova, I.; Saaya, S.; Shevchenko, A.; Stupnikova, A.; Zhiven’, M.; Laktionov, P.; Stepanova, A.; Romashchenko, A.; Yanshole, L.; Chernonosov, A.; et al. Mitomycin-treated endothelial and smooth muscle cells suitable for safe tissue engineering approaches. Front. Bioeng. Biotechnol. 2022, 10, 772981. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Xu, X.X.; Yu, S.X.; Wang, Y.R.; Liu, Y.; Liu, F.; Liu, W.; Li, X.L.; Luo, H.; Jing, G.; et al. Dynamics of Endothelial Cells Migration in Nature-Mimicking Blood Vessels. Talanta 2024, 277, 126415. [Google Scholar] [CrossRef] [PubMed]
- Bhogal, M.; Ang, H.P.; Lin, S.J.; Lwin, C.N.; Adnan, K.; Peh, G.; Mehta, J.S. Near infra-red labelling and tracking of corneal endothelial cells in-vivo. Sci. Rep. 2022, 12, 6338. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.B.; Ayachit, N.H.; Aminabhavi, T.M. Biosensors and microfluidic biosensors: From fabrication to application. Biosensors 2022, 12, 543. [Google Scholar] [CrossRef]
- Basak, M.; Kulkarni, M.; Narisepalli, S.; Chitkara, D.; Mittal, A. Exosomal fragment enclosed polyamine-salt nano-complex for co-delivery of docetaxel and mir-34a exhibits higher cytotoxicity and apoptosis in breast cancer cells. Sci. Rep. 2024, 14, 21669. [Google Scholar] [CrossRef]
- Yu, Y.; Wu, S.; Zhang, C.; Chen, F. Separation and enrichment of Sudan III using surface modified hollow glass microspheres and colorimetric detection. J. AOAC Int. 2021, 104, 165–171. [Google Scholar] [CrossRef]
- Greenwald, E.C.; Mehta, S.; Zhang, J. Genetically encoded fluorescent biosensors illuminate the spatiotemporal regulation of signaling networks. Chem. Rev. 2018, 118, 11707–11794. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.; Zhang, J. Biochemical activity architectures visualized–using genetically encoded fluorescent biosensors to map the spatial boundaries of signaling compartments. Acc. Chem. Res. 2021, 54, 2409–2420. [Google Scholar] [CrossRef] [PubMed]
- Sekhon, H.; Ha, J.H.; Presti, M.F.; Procopio, S.B.; Jarvis, A.R.; Mirsky, P.O.; John, A.M.; Loh, S.N. Adaptable, turn-on maturation (ATOM) fluorescent biosensors for multiplexed detection in cells. Nat. Methods 2023, 20, 1920–1929. [Google Scholar] [CrossRef]
- Kostyuk, A.I.; Demidovich, A.D.; Kotova, D.A.; Belousov, V.V.; Bilan, D.S. Circularly permuted fluorescent protein-based indicators: History, principles, and classification. Int. J. Mol. Sci. 2019, 20, 4200. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shang, Z.; Chen, J.H.; Gu, W.; Yao, L.; Yang, X.; Sun, X.; Wang, L.; Wang, T.; Liu, S.; et al. Engineering of NEMO as calcium indicators with large dynamics and high sensitivity. Nat. Methods 2023, 20, 918–924. [Google Scholar] [CrossRef] [PubMed]
- Baird, G.S.; Zacharias, D.A.; Tsien, R.Y. Circular permutation and receptor insertion within green fluorescent proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 11241–11246. [Google Scholar] [CrossRef]
- Takizawa, M.; Osuga, Y.; Ishida, R.; Mita, M.; Harada, K.; Ueda, H.; Kitaguchi, T.; Tsuboi, T. Development of a red fluorescent protein-based cGMP indicator applicable for live-cell imaging. Commun. Biol. 2022, 5, 833. [Google Scholar] [CrossRef]
- Dmitrieva, M.D.; Voitova, A.A.; Dymova, M.A.; Richter, V.A.; Kuligina, E.V. Tumor-targeting peptides search strategy for the delivery of therapeutic and diagnostic molecules to tumor cells. Int. J. Mol. Sci. 2020, 22, 314. [Google Scholar] [CrossRef]
- Ramaraju, H.; Kohn, D.H. Cell and material-specific phage display peptides increase iPS-MSC mediated bone and vasculature formation in vivo. Adv. Healthc. Mater. 2019, 8, 1801356. [Google Scholar] [CrossRef]
- Matsuda, S.; Harada, K.; Ito, M.; Takizawa, M.; Wongso, D.; Tsuboi, T.; Kitaguchi, T. Generation of a cGMP indicator with an expanded dynamic range by optimization of amino acid linkers between a fluorescent protein and PDE5α. ACS Sens. 2017, 2, 46–51. [Google Scholar] [CrossRef]
- Jing, M.; Zhang, P.; Wang, G.; Feng, J.; Mesik, L.; Zeng, J.; Jiang, H.; Wang, S.; Looby, J.C.; Guagliardo, N.A.; et al. A genetically encoded fluorescent acetylcholine indicator for in vitro and in vivo studies. Nat. Biotechnol. 2018, 36, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Wan, J.; Li, G.; Dong, H.; Xia, X.; Wang, Y.; Li, X.; Zhuang, C.; Zheng, Y.; Liu, L.; et al. Improved green and red GRAB sensors for monitoring spatiotemporal serotonin release in vivo. Nat. Methods 2024, 21, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Balleza, E.; Kim, J.M.; Cluzel, P. Systematic characterization of maturation time of fluorescent proteins in living cells. Nat. Methods 2018, 15, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Nisticò, N.; Aloisio, A.; Lupia, A.; Zimbo, A.M.; Mimmi, S.; Maisano, D.; Russo, R.; Marino, F.; Scalise, M.; Chiarella, E.; et al. Development of cyclic peptides targeting the epidermal growth factor receptor in mesenchymal triple-negative breast cancer subtype. Cells 2023, 12, 1078. [Google Scholar] [CrossRef] [PubMed]
- Lobas, M.A.; Tao, R.; Nagai, J.; Kronschläger, M.T.; Borden, P.M.; Marvin, J.S.; Looger, L.L.; Khakh, B.S. A genetically encoded single-wavelength sensor for imaging cytosolic and cell surface ATP. Nat. Commun. 2019, 10, 711. [Google Scholar] [CrossRef] [PubMed]
- Apostolou, E.; Blau, H.; Chien, K.; Lancaster, M.A.; Tata, P.R.; Trompouki, E.; Watt, F.M.; Zeng, Y.A.; Zernicka-Goetz, M. Progress and challenges in stem cell biology. Nat. Cell Biol. 2023, 25, 203–206. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, I.; Shin, K. A new era of stem cell and developmental biology: From blastoids to synthetic embryos and beyond. Exp. Mol. Med. 2023, 55, 2127–2137. [Google Scholar] [CrossRef]
- De Plano, L.M.; Oddo, S.; Guglielmino, S.P.; Caccamo, A.; Conoci, S. Generation of a helper phage for the fluorescent detection of peptide-target interactions by dual-display phages. Sci. Rep. 2023, 13, 18927. [Google Scholar] [CrossRef]
- Martins, I.M.; Canadas, R.F.; Pereira, H.; Azeredo, J.; Reis, R.L.; Oliveira, J.M.; Azevedo, H.S. Phage Display Identified Peptide with Selectivity for Human Osteoarthritic Chondrocytes. Adv. Ther. 2023, 6, 2300263. [Google Scholar] [CrossRef]
- De Plano, L.M.; Franco, D.; Bonsignore, M.; Fazio, E.; Trusso, S.; Allegra, A.; Musolino, C.; Cavaliere, R.; Ferlazzo, G.; Neri, F.; et al. Phage-phenotype imaging of myeloma plasma cells by phage display. Appl. Sci. 2021, 11, 7910. [Google Scholar] [CrossRef]
- Wang, F.; Zhong, Y.; Bruns, O.; Liang, Y.; Dai, H. In vivo NIR-II fluorescence imaging for biology and medicine. Nat. Photonics 2024, 18, 535–547. [Google Scholar] [CrossRef]
- Balyan, P.; Gupta, S.; Mavileti, S.K.; Pandey, S.S.; Kato, T. NIR-Sensitive Squaraine Dye—Peptide Conjugate for Trypsin Fluorogenic Detection. Biosensors 2024, 14, 458. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Q.; Shao, S.; Li, N.; Zhang, Z.; Liu, B. Non-Invasive On–Off Fluorescent Biosensor for Endothelial Cell Detection. Biosensors 2024, 14, 489. https://doi.org/10.3390/bios14100489
Jiang Q, Shao S, Li N, Zhang Z, Liu B. Non-Invasive On–Off Fluorescent Biosensor for Endothelial Cell Detection. Biosensors. 2024; 14(10):489. https://doi.org/10.3390/bios14100489
Chicago/Turabian StyleJiang, Qingyun, Shuai Shao, Na Li, Zhengyao Zhang, and Bo Liu. 2024. "Non-Invasive On–Off Fluorescent Biosensor for Endothelial Cell Detection" Biosensors 14, no. 10: 489. https://doi.org/10.3390/bios14100489
APA StyleJiang, Q., Shao, S., Li, N., Zhang, Z., & Liu, B. (2024). Non-Invasive On–Off Fluorescent Biosensor for Endothelial Cell Detection. Biosensors, 14(10), 489. https://doi.org/10.3390/bios14100489