Engineering and Characterization of 3-Aminotyrosine-Derived Red Fluorescent Variants of Circularly Permutated Green Fluorescent Protein



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials, Reagents, and General Methods
2.2. Plasmid Construction and Screening
2.3. Cell Culture and Imaging
2.4. Protein Expression and Characterization
3. Results
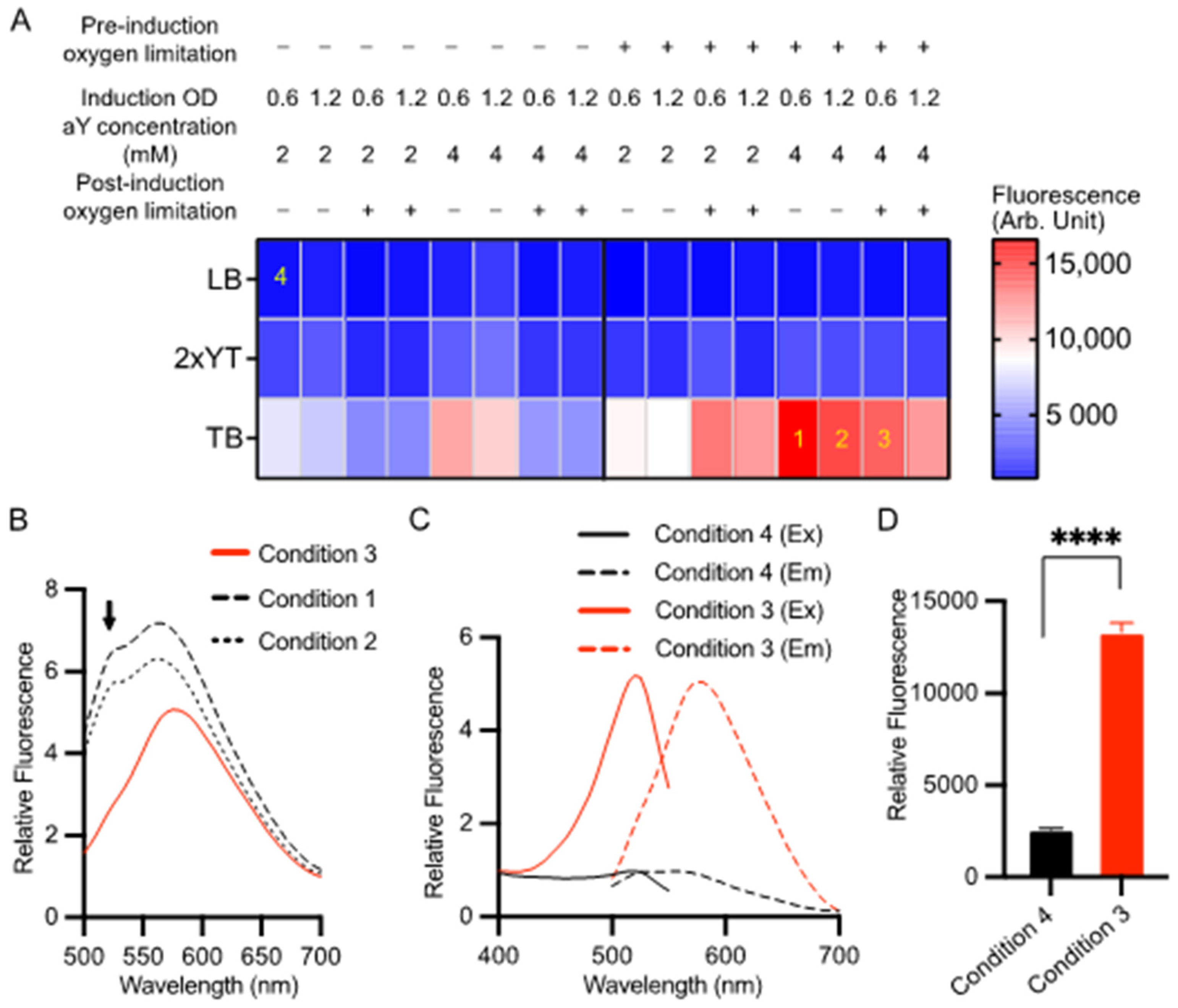
3.1. Optimization of the Expression of aY-cpGFP in Bacteria
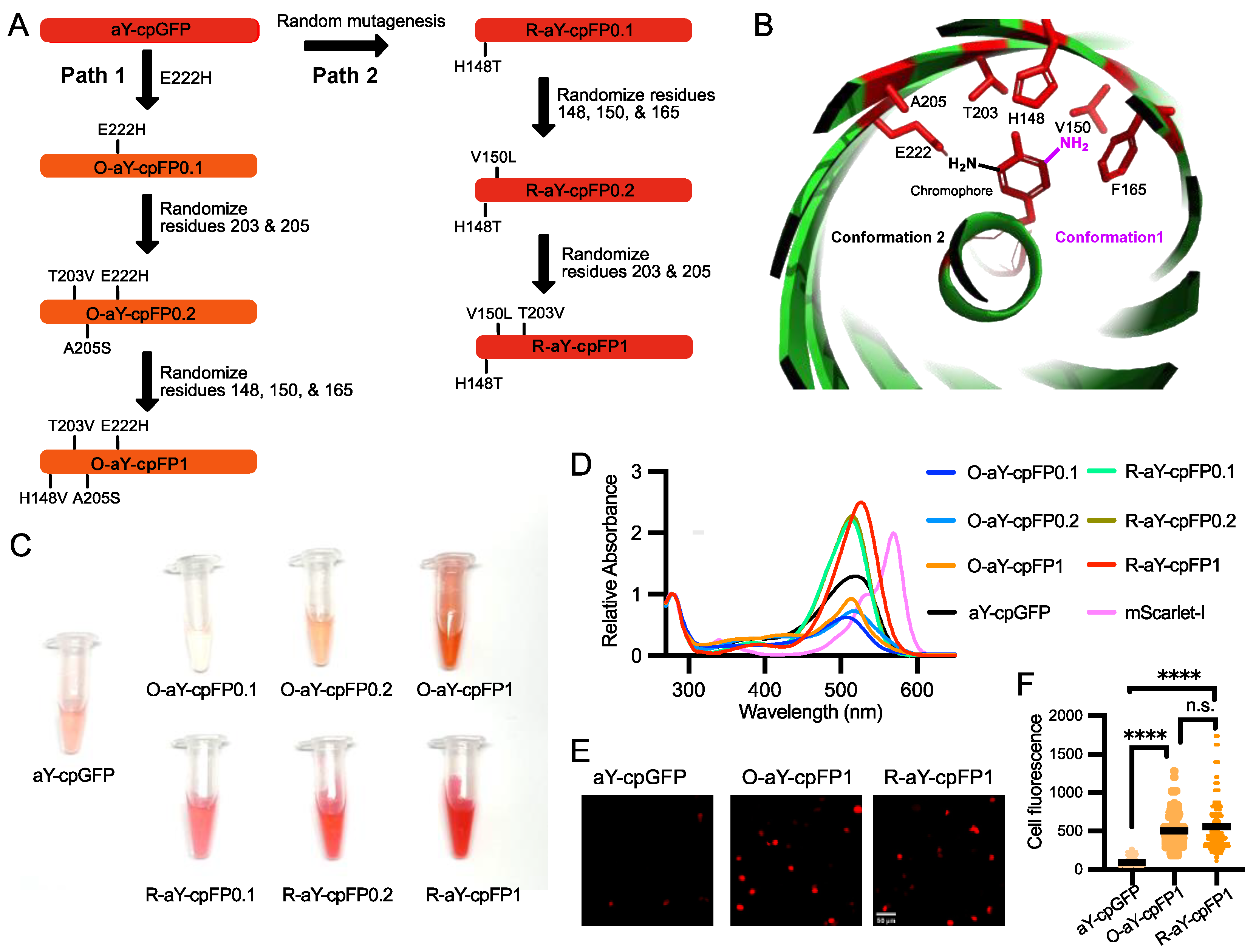
3.2. Brightness Improvement through Protein Engineering
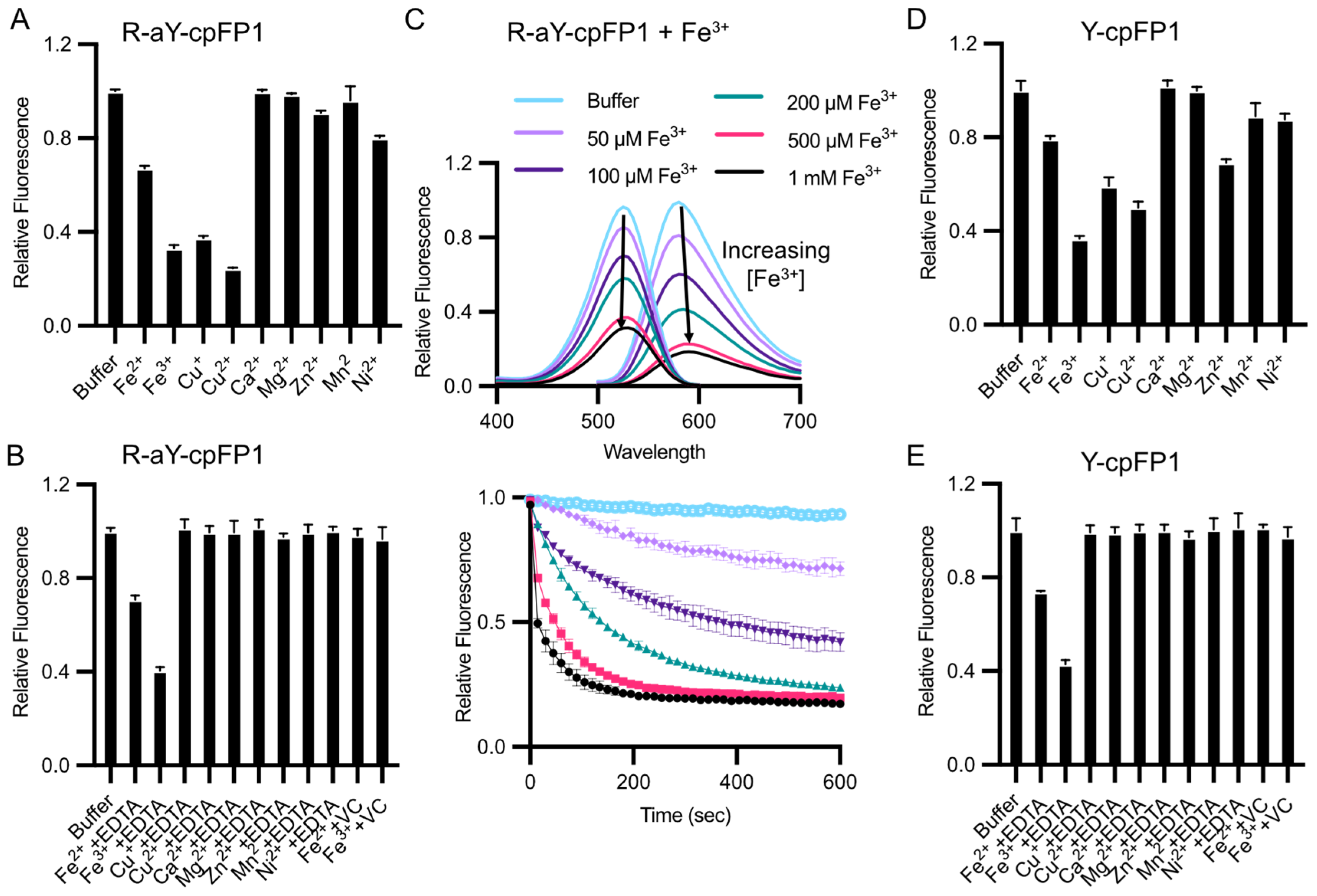
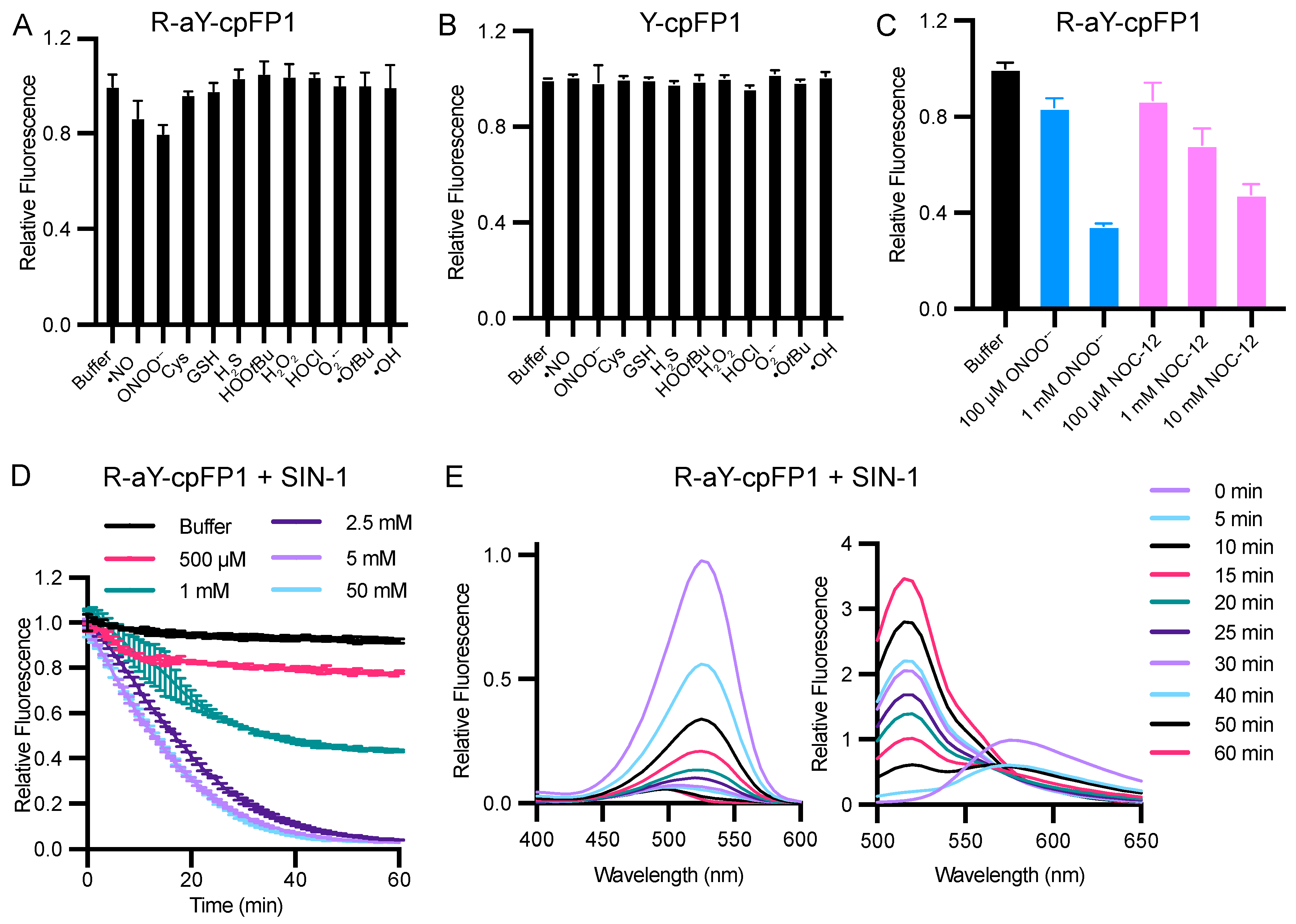
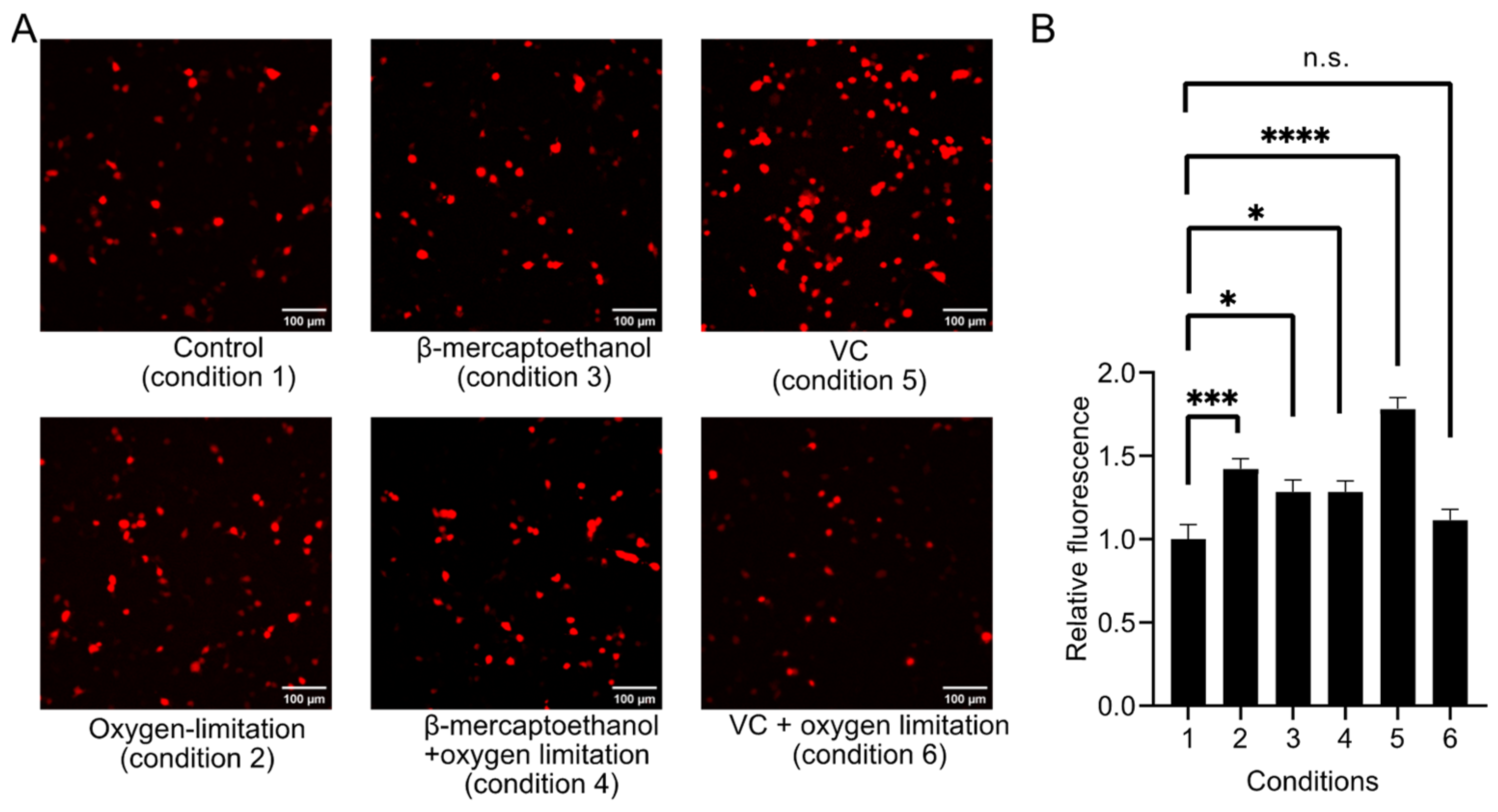
3.3. Metal Ion and Redox Responses of R-aY-cpFP1
3.4. Expression of R-aY-cpFP1 in Mammalian Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsien, R.Y. Constructing and exploiting the fluorescent protein paintbox (Nobel Lecture). Angew. Chem. Int. Ed. Engl. 2009, 48, 5612–5626. [Google Scholar] [CrossRef] [PubMed]
- Lambert, T.J. FPbase: A community-editable fluorescent protein database. Nat. Methods 2019, 16, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.E.; Tour, O.; Palmer, A.E.; Steinbach, P.A.; Baird, G.S.; Zacharias, D.A.; Tsien, R.Y. A monomeric red fluorescent protein. Proc. Natl. Acad. Sci. USA 2002, 99, 7877–7882. [Google Scholar] [CrossRef] [PubMed]
- Bindels, D.S.; Haarbosch, L.; van Weeren, L.; Postma, M.; Wiese, K.E.; Mastop, M.; Aumonier, S.; Gotthard, G.; Royant, A.; Hink, M.A.; et al. mScarlet: A bright monomeric red fluorescent protein for cellular imaging. Nat. Methods 2017, 14, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Mo, G.C.H.; Posner, C.; Rodriguez, E.A.; Sun, T.; Zhang, J. A rationally enhanced red fluorescent protein expands the utility of FRET biosensors. Nat. Commun. 2020, 11, 1848. [Google Scholar] [CrossRef] [PubMed]
- Akerboom, J.; Carreras Calderon, N.; Tian, L.; Wabnig, S.; Prigge, M.; Tolo, J.; Gordus, A.; Orger, M.B.; Severi, K.E.; Macklin, J.J.; et al. Genetically encoded calcium indicators for multi-color neural activity imaging and combination with optogenetics. Front. Mol. Neurosci. 2013, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Kumar, M.; Zhang, J.; Zhao, S.; Drobizhev, M.; McCollum, M.; Anderson, C.T.; Wang, Y.; Pokorny, A.; Tian, X.; et al. A genetically encoded far-red fluorescent indicator for imaging synaptically released Zn2+. Sci. Adv. 2023, 9, eadd2058. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Araki, S.; Wu, J.; Teramoto, T.; Chang, Y.F.; Nakano, M.; Abdelfattah, A.S.; Fujiwara, M.; Ishihara, T.; Nagai, T.; et al. An expanded palette of genetically encoded Ca2+ indicators. Science 2011, 333, 1888–1891. [Google Scholar] [CrossRef]
- Matz, M.V.; Fradkov, A.F.; Labas, Y.A.; Savitsky, A.P.; Zaraisky, A.G.; Markelov, M.L.; Lukyanov, S.A. Fluorescent proteins from nonbioluminescent Anthozoa species. Nat. Biotechnol. 1999, 17, 969–973. [Google Scholar] [CrossRef]
- Shagin, D.A.; Barsova, E.V.; Yanushevich, Y.G.; Fradkov, A.F.; Lukyanov, K.A.; Labas, Y.A.; Semenova, T.N.; Ugalde, J.A.; Meyers, A.; Nunez, J.M. GFP-like proteins as ubiquitous metazoan superfamily: Evolution of functional features and structural complexity. Mol. Biol. Evol. 2004, 21, 841–850. [Google Scholar] [CrossRef]
- Shaner, N.C.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.N.G.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 2004, 22, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Gadella, T.W.J., Jr.; van Weeren, L.; Stouthamer, J.; Hink, M.A.; Wolters, A.H.G.; Giepmans, B.N.G.; Aumonier, S.; Dupuy, J.; Royant, A. mScarlet3: A brilliant and fast-maturing red fluorescent protein. Nat. Methods 2023, 20, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, E.C.; Mehta, S.; Zhang, J. Genetically Encoded Fluorescent Biosensors Illuminate the Spatiotemporal Regulation of Signaling Networks. Chem. Rev. 2018, 118, 11707–11794. [Google Scholar] [CrossRef] [PubMed]
- Baird, G.S.; Zacharias, D.A.; Tsien, R.Y. Circular permutation and receptor insertion within green fluorescent proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 11241–11246. [Google Scholar] [CrossRef] [PubMed]
- Topell, S.; Hennecke, J.; Glockshuber, R. Circularly permuted variants of the green fluorescent protein. FEBS Lett. 1999, 457, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Pandey, N.; Kuypers, B.E.; Nassif, B.; Thomas, E.E.; Alnahhas, R.N.; Segatori, L.; Silberg, J.J. Tolerance of a knotted near-Infrared fluorescent protein to random circular permutation. Biochemistry 2016, 55, 3763–3773. [Google Scholar] [CrossRef] [PubMed]
- Shui, B.; Wang, Q.; Lee, F.; Byrnes, L.J.; Chudakov, D.M.; Lukyanov, S.A.; Sondermann, H.; Kotlikoff, M.I. Circular permutation of red fluorescent proteins. PLoS ONE 2011, 6, e20505. [Google Scholar] [CrossRef]
- Huang, Y.M.; Bystroff, C. Complementation and reconstitution of fluorescence from circularly permuted and truncated green fluorescent protein. Biochemistry 2009, 48, 929–940. [Google Scholar] [CrossRef]
- Nasu, Y.; Shen, Y.; Kramer, L.; Campbell, R.E. Structure- and mechanism-guided design of single fluorescent protein-based biosensors. Nat. Chem. Biol. 2021, 17, 509–518. [Google Scholar] [CrossRef]
- Wiedenmann, J.; Ivanchenko, S.; Oswald, F.; Schmitt, F.; Röcker, C.; Salih, A.; Spindler, K.-D.; Nienhaus, G.U. EosFP, a fluorescent marker protein with UV-inducible green-to-red fluorescence conversion. Proc. Natl. Acad. Sci. USA 2004, 101, 15905–15910. [Google Scholar] [CrossRef]
- Gurskaya, N.G.; Verkhusha, V.V.; Shcheglov, A.S.; Staroverov, D.B.; Chepurnykh, T.V.; Fradkov, A.F.; Lukyanov, S.; Lukyanov, K.A. Engineering of a monomeric green-to-red photoactivatable fluorescent protein induced by blue light. Nat. Biotechnol. 2006, 24, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Moeyaert, B.; Nguyen Bich, N.; De Zitter, E.; Rocha, S.; Clays, K.; Mizuno, H.; Van Meervelt, L.; Hofkens, J.; Dedecker, P. Green-to-red photoconvertible Dronpa mutant for multimodal super-resolution fluorescence microscopy. ACS Nano 2014, 8, 1664–1673. [Google Scholar] [CrossRef]
- Ando, R.; Hama, H.; Yamamoto-Hino, M.; Mizuno, H.; Miyawaki, A. An optical marker based on the UV-induced green-to-red photoconversion of a fluorescent protein. Proc. Natl. Acad. Sci. USA 2002, 99, 12651–12656. [Google Scholar] [CrossRef] [PubMed]
- Sawin, K.E.; Nurse, P. Photoactivation of green fluorescent protein. Curr. Biol. 1997, 7, R606–R607. [Google Scholar] [CrossRef] [PubMed]
- Elowitz, M.B.; Surette, M.G.; Wolf, P.-E.; Stock, J.; Leibler, S. Photoactivation turns green fluorescent protein red. Curr. Biol. 1997, 7, 809–812. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, A.M.; Mishin, A.S.; Yampolsky, I.V.; Belousov, V.V.; Chudakov, D.M.; Subach, F.V.; Verkhusha, V.V.; Lukyanov, S.; Lukyanov, K.A. Green fluorescent proteins are light-induced electron donors. Nat. Chem. Biol. 2009, 5, 459–461. [Google Scholar] [CrossRef]
- Mishin, A.S.; Subach, F.V.; Yampolsky, I.V.; King, W.; Lukyanov, K.A.; Verkhusha, V.V. The first mutant of the Aequorea victoria green fluorescent protein that forms a red chromophore. Biochemistry 2008, 47, 4666–4673. [Google Scholar] [CrossRef]
- Saha, R.; Verma, P.K.; Rakshit, S.; Saha, S.; Mayor, S.; Pal, S.K. Light driven ultrafast electron transfer in oxidative redding of Green Fluorescent Proteins. Sci. Rep. 2013, 3, 1580. [Google Scholar] [CrossRef]
- Imamura, H.; Otsubo, S.; Nishida, M.; Takekawa, N.; Imada, K. Red fluorescent proteins engineered from green fluorescent proteins. Proc. Natl. Acad. Sci. USA 2023, 120, e2307687120. [Google Scholar] [CrossRef]
- Liu, C.C.; Schultz, P.G. Adding new chemistries to the genetic code. Annu. Rev. Biochem. 2010, 79, 413–444. [Google Scholar] [CrossRef]
- Wang, L.; Xie, J.; Deniz, A.A.; Schultz, P.G. Unnatural amino acid mutagenesis of green fluorescent protein. J. Org. Chem. 2003, 68, 174–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Niu, W.; Guo, J.T.; Schultz, P.G. Unnatural Amino Acid Mutagenesis of Fluorescent Proteins. Angew. Chem. Int. Ed. 2012, 51, 10132–10135. [Google Scholar] [CrossRef] [PubMed]
- Wals, K.; Ovaa, H. Unnatural amino acid incorporation in E. coli: Current and future applications in the design of therapeutic proteins. Front. Chem. 2014, 2, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, X.; Guo, X.; Li, J.; Liu, Q.; Kang, F.; Wang, X.; Hu, C.; Liu, H.; Gong, W.; et al. Significant expansion and red-shifting of fluorescent protein chromophore determined through computational design and genetic code expansion. Biophys. Rep. 2018, 4, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.H.; Rubini, M.; Jung, G.; Wiegand, G.; Seifert, M.H.; Azim, M.K.; Kim, J.S.; Zumbusch, A.; Holak, T.A.; Moroder, L.; et al. Expansion of the genetic code enables design of a novel “gold” class of green fluorescent proteins. J. Mol. Biol. 2003, 328, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ai, H.-w. A general strategy to red-shift green fluorescent protein-based biosensors. Nat. Chem. Biol. 2020, 16, 1434–1439. [Google Scholar] [CrossRef] [PubMed]
- Augustine, G.; Raghavan, S.; NumbiRamudu, K.; Easwaramoorthi, S.; Shanmugam, G.; Seetharani Murugaiyan, J.; Gunasekaran, K.; Govind, C.; Karunakaran, V.; Ayyadurai, N. Excited State Electronic Interconversion and Structural Transformation of Engineered Red-Emitting Green Fluorescent Protein Mutant. J. Phys. Chem. B 2019, 123, 2316–2324. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, H.; Zhang, J.; Ai, H.-w.; Fang, C. Structural origin and rational development of bright red noncanonical variants of green fluorescent protein. Phys. Chem. Chem. Phys. 2023, 25, 15624–15634. [Google Scholar] [CrossRef]
- Pedelacq, J.D.; Cabantous, S.; Tran, T.; Terwilliger, T.C.; Waldo, G.S. Engineering and characterization of a superfolder green fluorescent protein. Nat. Biotechnol. 2006, 24, 79–88. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, S.; Li, X.; Ai, H.-w. A high-performance genetically encoded fluorescent biosensor for imaging physiological peroxynitrite. Cell Chem. Biol. 2021, 28, 1542–1553.e5. [Google Scholar] [CrossRef]
- Cadwell, R.C.; Joyce, G.F. Randomization of genes by PCR mutagenesis. Genome Res. 1992, 2, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Zhang, H.; Ai, H.-w. Improved red fluorescent redox indicators for monitoring cytosolic and mitochondrial thioredoxin redox dynamics. Biochemistry 2022, 61, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Romei, M.G.; Lin, C.-Y.; Mathews, I.I.; Boxer, S.G. Electrostatic control of photoisomerization pathways in proteins. Biophys. J. 2020, 118, 609a. [Google Scholar] [CrossRef]
- Carballal, S.; Bartesaghi, S.; Radi, R. Kinetic and mechanistic considerations to assess the biological fate of peroxynitrite. Biochim. Biophys. Acta 2014, 1840, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Martin-Romero, F.J.; Gutierrez-Martin, Y.; Henao, F.; Gutierrez-Merino, C. Fluorescence measurements of steady state peroxynitrite production upon SIN-1 decomposition: NADH versus dihydrodichlorofluorescein and dihydrorhodamine 123. J. Fluoresc. 2004, 14, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, L.; Matsuda, T.; Zhao, Y.; Rebane, A.; Drobizhev, M.; Chang, Y.F.; Araki, S.; Arai, Y.; March, K.; et al. Improved orange and red Ca2+ indicators and photophysical considerations for optogenetic applications. ACS Chem. Neurosci. 2013, 4, 963–972. [Google Scholar] [CrossRef]
- Ermakova, Y.G.; Bilan, D.S.; Matlashov, M.E.; Mishina, N.M.; Markvicheva, K.N.; Subach, O.M.; Subach, F.V.; Bogeski, I.; Hoth, M.; Enikolopov, G.; et al. Red fluorescent genetically encoded indicator for intracellular hydrogen peroxide. Nat. Commun. 2014, 5, 5222. [Google Scholar] [CrossRef]
- Hassanein, M.; Abdo, M.; Gerges, S.; El-Khalafy, S. Study of the oxidation of 2-aminophenol by molecular oxygen catalyzed by cobalt (II) phthalocyaninetetrasodiumsulfonate in water. J. Mol. Catal. A Chem. 2008, 287, 53–56. [Google Scholar] [CrossRef]
- Mohammed, T.P.; George, A.; Sivaramakrishnan, M.P.; Vadivelu, P.; Balasubramanian, S.; Sankaralingam, M. Deciphering the effect of amine versus imine ligands of copper (II) complexes in 2-aminophenol oxidation. J. Inorg. Biochem. 2023, 247, 112309. [Google Scholar] [CrossRef]
- Podder, N.; Mandal, S. Aerobic oxidation of 2-aminophenol catalysed by a series of mononuclear copper (ii) complexes: Phenoxazinone synthase-like activity and mechanistic study. New J. Chem. 2020, 44, 12793–12805. [Google Scholar] [CrossRef]
- Szávuly, M.; Csonka, R.; Speier, G.; Barabás, R.; Giorgi, M.; Kaizer, J. Oxidation of 2-aminophenol by iron (III) isoindoline complexes. J. Mol. Catal. A Chem. 2014, 392, 120–126. [Google Scholar] [CrossRef]
- Çeşme, M. 2-Aminophenol-based ligands and Cu(II) complexes: Synthesis, characterization, X-ray structure, thermal and electrochemical properties, and in vitro biological evaluation, ADMET study and molecular docking simulation. J. Mol. Struct. 2023, 1271, 134073. [Google Scholar] [CrossRef]
- Beltrán, A.; Burguete, M.I.; Abánades, D.R.; Pérez-Sala, D.; Luis, S.V.; Galindo, F. Turn-on fluorescent probes for nitric oxide sensing based on the ortho-hydroxyamino structure showing no interference with dehydroascorbic acid. Chem. Commun. 2014, 50, 3579–3581. [Google Scholar] [CrossRef] [PubMed]
- Kwun, M.S.; Lee, D.G. Investigation of distinct contribution of nitric oxide and each reactive oxygen species in indole-3-propionic-acid-induced apoptosis-like death in Escherichia coli. Life Sci. 2021, 285, 120003. [Google Scholar] [CrossRef] [PubMed]
- Corker, H.; Poole, R.K. Nitric oxide formation by Escherichia coli. Dependence on nitrite reductase, the NO-sensing regulator Fnr, and flavohemoglobin Hmp. J. Biol. Chem. 2003, 278, 31584–31592. [Google Scholar] [CrossRef]
- Deeb, R.S.; Nuriel, T.; Cheung, C.; Summers, B.; Lamon, B.D.; Gross, S.S.; Hajjar, D.P. Characterization of a cellular denitrase activity that reverses nitration of cyclooxygenase. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H687–H698. [Google Scholar] [CrossRef]
- Smallwood, H.S.; Lourette, N.M.; Boschek, C.B.; Bigelow, D.J.; Smith, R.D.; Pasa-Tolic, L.; Squier, T.C. Identification of a denitrase activity against calmodulin in activated macrophages using high-field liquid chromatography--FTICR mass spectrometry. Biochemistry 2007, 46, 10498–10505. [Google Scholar] [CrossRef]
- Irie, Y.; Saeki, M.; Kamisaki, Y.; Martin, E.; Murad, F. Histone H1.2 is a substrate for denitrase, an activity that reduces nitrotyrosine immunoreactivity in proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 5634–5639. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | λex (nm) | λem (nm) | Bacterial Lysate Brightness | ε (mM−1 cm−1) | ϕ | Molecular Brightness 1 | τ (ns) | pKa 2 |
|---|---|---|---|---|---|---|---|---|
| aY-cpGFP | 521 | 590 | 100% | 48.7 | 0.14 | 6.7 | 1.0 | 6.8 |
| O-aY-cpFP0.1 | 512 | 571 | 45% | 39.1 | 0.26 | 10.1 | 2.7 | 7.2 |
| O-aY-cpFP0.2 | 515 | 577 | 80% | 46.6 | 0.21 | 9.9 | 2.6 | 7.4 |
| O-aY-cpFP1 | 513 | 577 | 139% | 39.2 | 0.29 | 11.5 | 2.6 | 7.0 |
| R-aY-cpFP0.1 | 515 | 590 | 561% | 114.7 | 0.16 | 18.6 | 1.2 | 6.3 |
| R-aY-cpFP0.2 | 516 | 589 | 748% | 118.8 | 0.16 | 19.0 | 1.2 | 6.3 |
| R-aY-cpFP1 | 528 | 580 | 1312% | 124.4 | 0.15 | 18.7 | 1.1 | 6.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Tian, X.; Zhang, J.; Ai, H.-w. Engineering and Characterization of 3-Aminotyrosine-Derived Red Fluorescent Variants of Circularly Permutated Green Fluorescent Protein. Biosensors 2024, 14, 54. https://doi.org/10.3390/bios14010054
Zhang H, Tian X, Zhang J, Ai H-w. Engineering and Characterization of 3-Aminotyrosine-Derived Red Fluorescent Variants of Circularly Permutated Green Fluorescent Protein. Biosensors. 2024; 14(1):54. https://doi.org/10.3390/bios14010054
Chicago/Turabian StyleZhang, Hao, Xiaodong Tian, Jing Zhang, and Hui-wang Ai. 2024. "Engineering and Characterization of 3-Aminotyrosine-Derived Red Fluorescent Variants of Circularly Permutated Green Fluorescent Protein" Biosensors 14, no. 1: 54. https://doi.org/10.3390/bios14010054
APA StyleZhang, H., Tian, X., Zhang, J., & Ai, H.-w. (2024). Engineering and Characterization of 3-Aminotyrosine-Derived Red Fluorescent Variants of Circularly Permutated Green Fluorescent Protein. Biosensors, 14(1), 54. https://doi.org/10.3390/bios14010054