Microalgae-Based Fluorimetric Bioassays for Studying Interferences on Photosynthesis Induced by Environmentally Relevant Concentrations of the Herbicide Diuron






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chlamydomonas reinhardtii Cell Suspensions
2.2. Herbicide DCMU Solutions
2.3. Microalgae-Based Bioassays
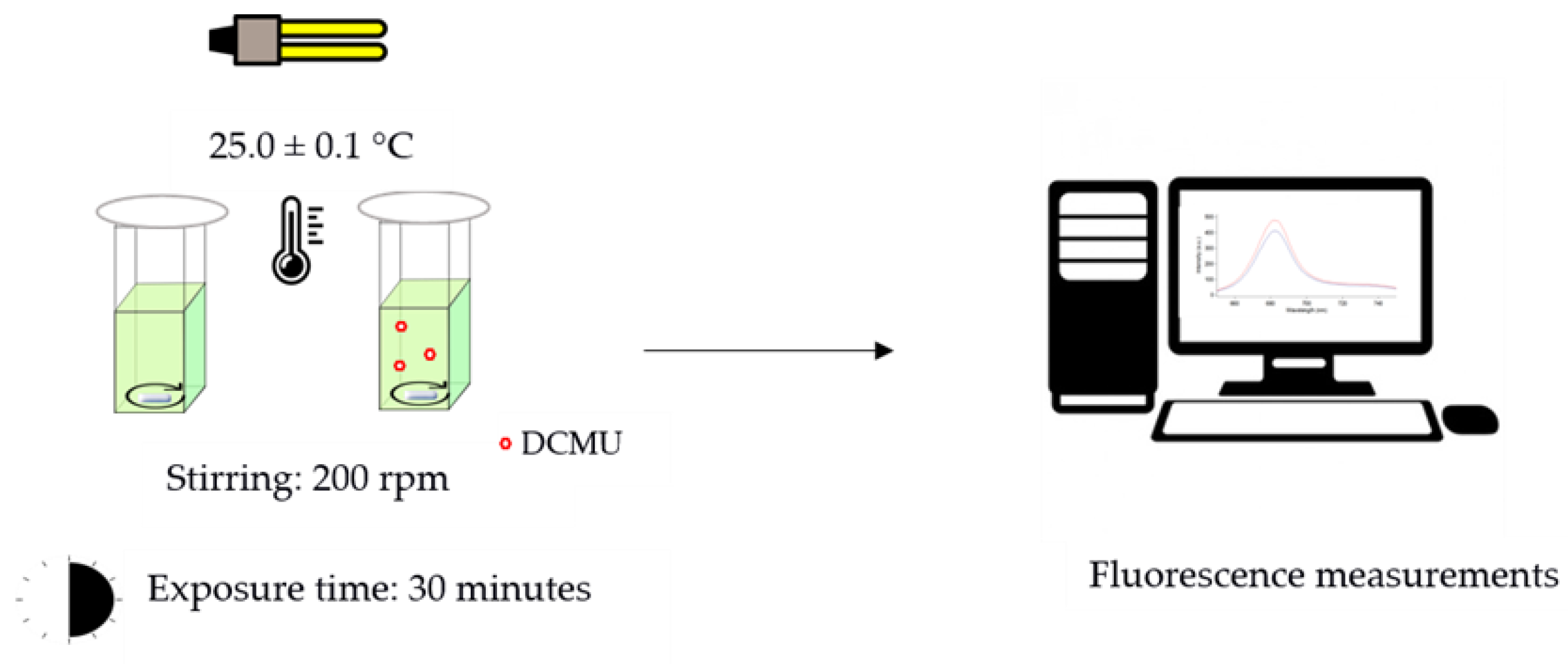
2.3.1. Measurements of In Vivo Steady-State Chl-a Fluorescence
- Control microalgae cell suspensions: 2.70 mL of microalgae cell suspension (OD750 nm of 0.20 ± 0.02) + 0.30 mL of high-purity deionized water.
- DCMU-exposed microalgae cell suspensions: 2.70 mL of microalgae cell suspension (OD750 nm of 0.20 ± 0.02) + 0.30 mL of DCMU working solutions (10 times more concentrated than the final concentrations tested, i.e., 0.2, 2, and 20 μg/L).
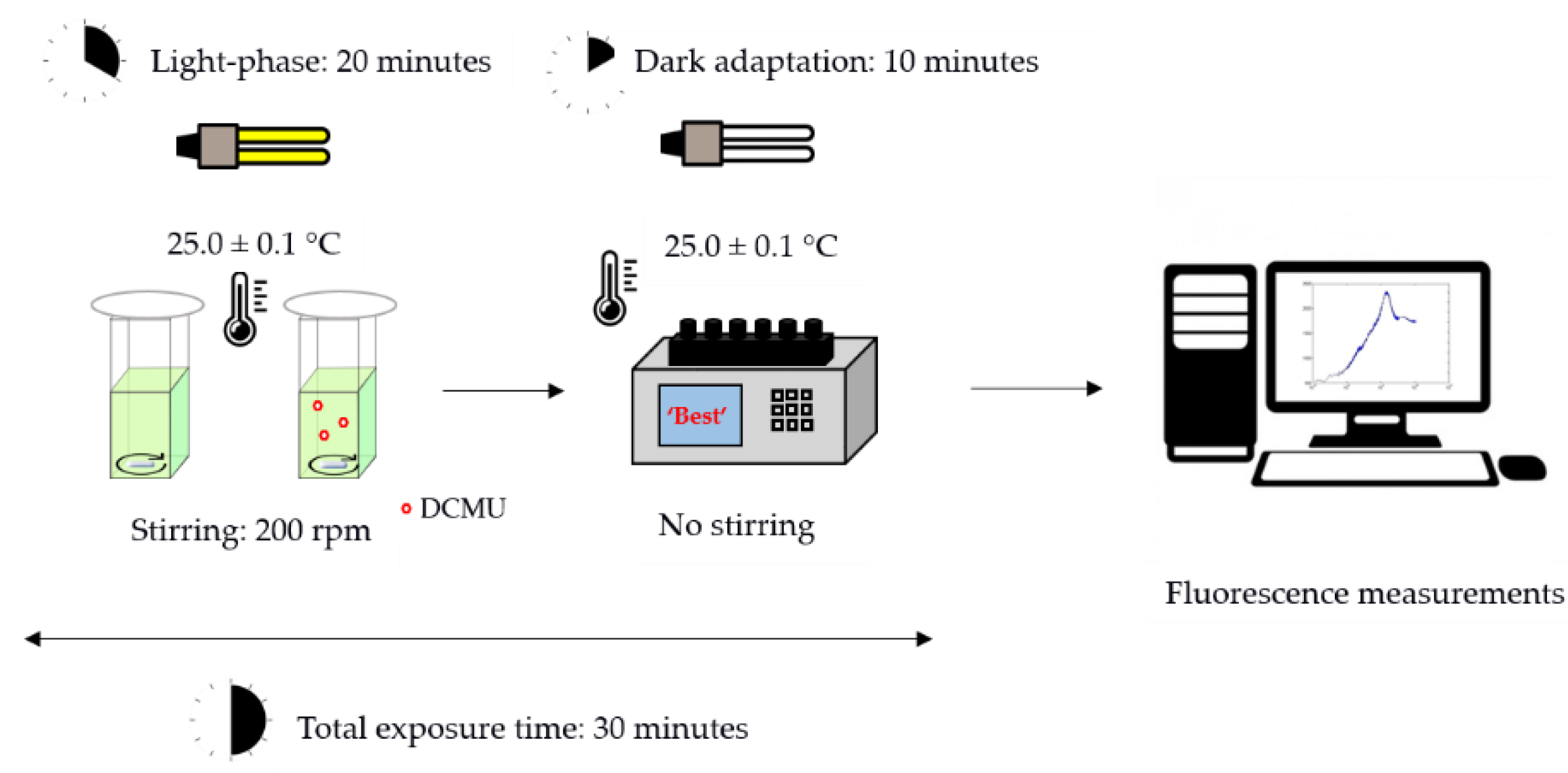
2.3.2. Kinetic Measurements of a Chl-a Fluorescence Transient
- Fo (minimum of fluorescence intensity yield in the absence of photosynthetic light): calculated with data between 0.1 and 0.14 ms as the intercept of linear regression;
- FM (maximum of fluorescence intensity);
- F8 ms (fluorescence intensity at 8 ms): average fluorescence values acquired at 8.0 ± 0.1 ms;
- V8 ms (the relative variable fluorescence at 8 ms) calculated as follows:V8 ms = (F8 ms − Fo)/(FM − Fo)
2.4. Data Analysis
3. Results
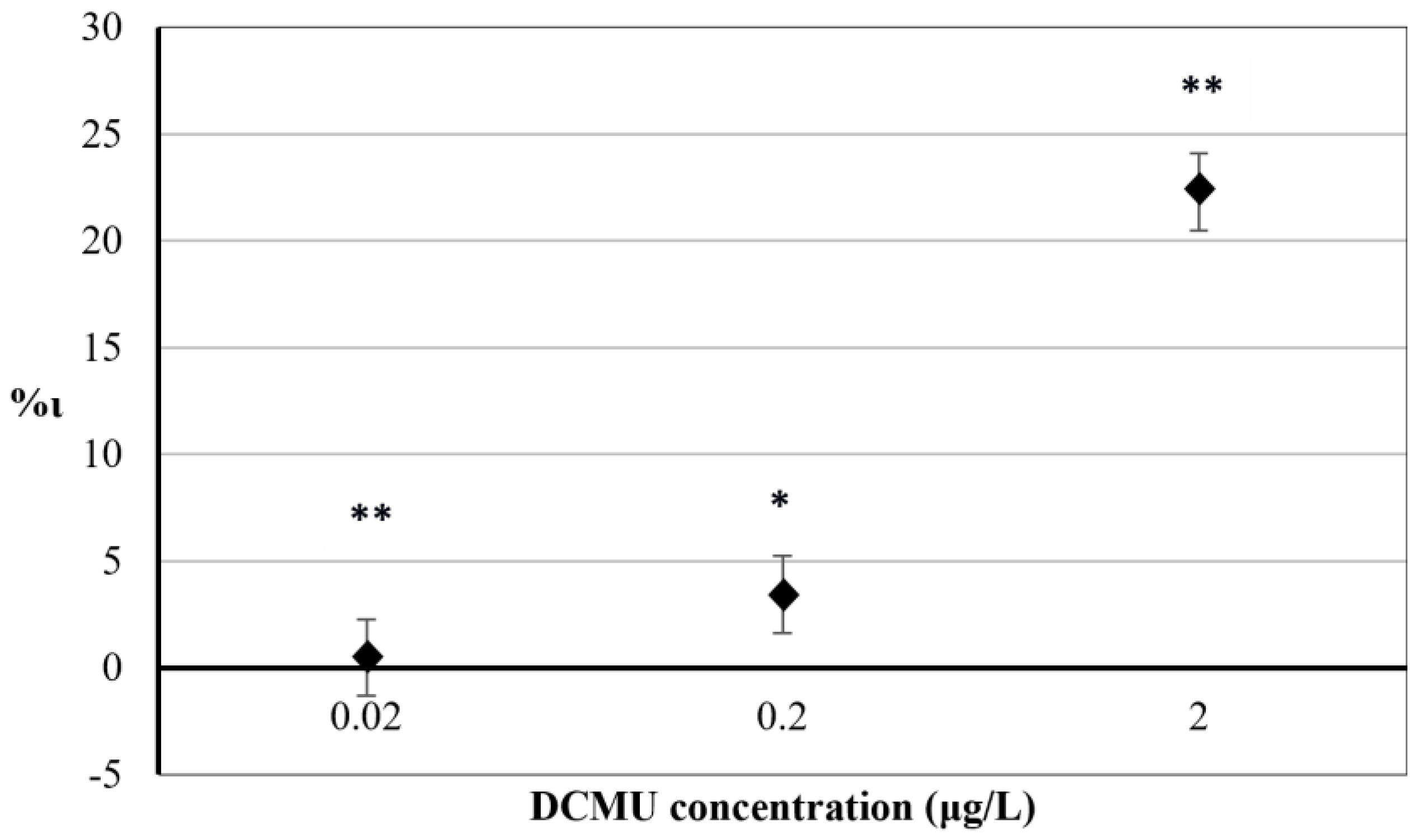
3.1. In Vivo Steady-State Fluorescence Emission of Chl-a
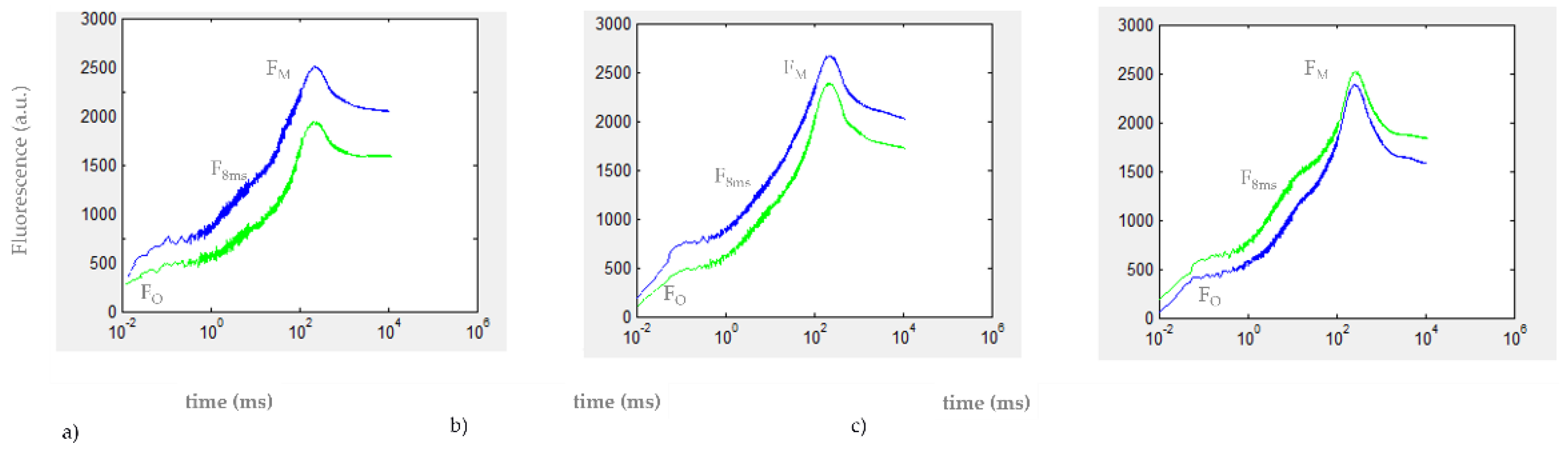
3.2. Chlorophyll a Fluorescence Transient
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Green, P.G.; Young, T.M. Loading of the herbicide diuron into the California water system. Environ. Eng. Sci. 2006, 23, 545–551. [Google Scholar]
- Peña, F.; Cárdenas, S.; Gallego, M.; Valcárcel, M. Analysis of phenylurea herbicides from plants by GC/MS. Talanta 2002, 56, 727–734. [Google Scholar]
- Giacomazzi, S.; Cochet, N. Environmental impact of diuron transformation: A review. Chemosphere 2004, 56, 1021–1032. [Google Scholar]
- Da Rocha, M.S.; Arnold, L.L.; Pennington, K.L.; Muirhead, D.; Dodmane, P.R.; Anwar, M.M.; Cohen, S.M. Diuron-induced rat bladder epithelial cytotoxicity. Toxicol. Sci. 2012, 130, 281–288. [Google Scholar]
- European Food Safety Authority. Reasoned opinion of EFSA: Review of the Existing Maximum Residue Levels (MRLs) for diuron according to article 12 of regulation (EC) No. 396/2005. EFSA J. 2011, 9, 2344. [Google Scholar] [CrossRef] [Green Version]
- Agbaogun, B.K.; Fischer, K. Adsorption of phenylurea herbicides by tropical soils. Environ. Monit. Assess. 2020, 192, 212. [Google Scholar]
- Liu, Y.; Xu, Z.; Wu, X.; Gui, W.; Zhu, G. Adsorption and desorption behavior of herbicide diuron on various Chinese cultivated soils. J. Hazard. Mater. 2010, 178, 462–468. [Google Scholar]
- Marlatt, V.L.; Martyniuk, C.J. Biological responses to phenylurea herbicides in fish and amphibians: New directions for characterizing mechanisms of toxicity. Comp. Biochem. Physiol. C Toxicol. Pharm. 2017, 194, 9–21. [Google Scholar]
- Kamarudin, N.A.; Zulkifli, S.Z.; Azmai, M.N.A.; Aziz, F.Z.A.; Ismail, A. Herbicide diuron as endocrine disrupting chemicals (EDCs) through histopathalogical analysis in gonads of Javanese medaka (Oryzias javanicus, Bleeker 1854). Animals 2020, 10, 525. [Google Scholar]
- Moreira, L.B.; Diamante, G.; Giroux, M.; Coffin, S.; Xu, E.G.; Moledo de Souza Abessa, D.; Schlenk, D. Impacts of salinity and temperature on the thyroidogenic effects of the biocide diuron in Menidia beryllina. Environ. Sci. Technol. 2018, 52, 3146–3155. [Google Scholar]
- Moon, Y.S.; Kim, M.; Hong, C.P.; Kang, J.H.; Jung, J.H. Overlapping and unique toxic effects of three alternative antifouling biocides (Diuron, Irgarol 1051®, Sea-Nine 211®) on non-target marine fish. Ecotoxicol. Environ. Saf. 2019, 180, 23–32. [Google Scholar]
- Nestler, H.; Groh, K.J.; Schönenberger, R.; Behra, R.; Schirmer, K.; Eggen, R.I.; Suter, M.J.F. Multiple-endpoint assay provides a detailed mechanistic view of responses to herbicide exposure in Chlamydomonas reinhardtii. Aquat. Toxicol. 2012, 214–224. [Google Scholar]
- Morin, S.; Chaumet, B.; Mazzella, N. A time-dose response model to assess diuron-induced photosynthesis inhibition in freshwater biofilms. Front. Environ. Sci. 2018, 6, 131. [Google Scholar]
- Chaumet, B.; Morin, S.; Hourtané, O.; Artigas, J.; Delest, B.; Eon, M.; Mazzella, N. Flow conditions influence diuron toxicokinetics and toxicodynamics in freshwater biofilms. Sci. Total Environ. 2019, 652, 1242–1251. [Google Scholar]
- Sasso, S.; Stibor, H.; Mittag, M.; Grossman, A.R. The Natural History of Model Organisms: From molecular manipulation of domesticated Chlamydomonas reinhardtii to survival in nature. Elife 2018, 7, e39233. [Google Scholar]
- Cid, A.; Prado, R.; Rioboo, C.; Suarez-Bregua, P.; Herrero, C. Use of Microalgae as Biological Indicators of Pollution: Looking for New Relevant Cytotoxicity Endpoints; Johnsen, M.N., Ed.; Nova Science Publishers: New York, NY, USA, 2012; pp. 311–323. [Google Scholar]
- Directive 2013/39/EU of the European Parliament and of the Council of 12 August 2013 amending Directives 2000/60/EC and 2008/105/EC as Regards Priority Substances in the Field of Water Policy. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwidpb7-6cH1AhXOCd4KHdFZAjYQFnoECAUQAQ&url=https%3A%2F%2Feur-lex.europa.eu%2FLexUriServ%2FLexUriServ.do%3Furi%3DOJ%3AL%3A2013%3A226%3A0001%3A0017%3Aen%3APDF&usg=AOvVaw1YjeR01_YUxTs_BAKtm311 (accessed on 8 August 2021).
- Council Directive 98/83/EC of 3 November 1998 on the Quality of Water Intended for Human Consumption. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A31998L0083 (accessed on 8 August 2021).
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dabrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar]
- Griffiths, M.J.; Garcin, C.; van Hille, R.P.; Harrison, S.T. Interference by pigment in the estimation of microalgal biomass concentration by optical density. J. Microbiol. Methods 2011, 85, 119–123. [Google Scholar]
- Goswami, A.; Jiang, J.Q. Simultaneous quantification of gabapentin, sulfamethoxazole, terbutryn, terbuthylazine and diuron by uv-vis spectrophotometer. Biointerface Res. Appl. Chem. 2018, 8, 3111–3117. [Google Scholar]
- Franck, F.; Juneau, P.; Popovic, R. Resolution of the photosystem I and photosystem II contributions to chlorophyll fluorescence of intact leaves at room temperature. Biochim. Biophys. Acta. (BBA)-Bioenerg 2002, 1556, 239–246. [Google Scholar]
- Jee, G. Sixty-three years since Kautsky: Chlorophyll a fluorescence. Aust. J. Plant Physiol. 1995, 22, 131–160. [Google Scholar]
- Eullaffroy, P.; Vernet, G. The F684/F735 chlorophyll fluorescence ratio: A potential tool for rapid detection and determination of herbicide phytotoxicity in algae. Water Res. 2003, 37, 1983–1990. [Google Scholar]
- Fai, P.B.; Grant, A.; Reid, B. Chlorophyll a fluorescence as a biomarker for rapid toxicity assessment. Environ. Toxicol. Chem. 2007, 26, 1520–1531. [Google Scholar]
- Stirbet, A. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B 2011, 104, 236–257. [Google Scholar]
- Stirbet, A.; Riznichenko, G.Y.; Rubin, A.B. Modeling chlorophyll a fluorescence transient: Relation to photosynthesis. Biochemistry 2014, 79, 291–323. [Google Scholar]
- Lombardo, A.; Boselli, C.; Amatiste, S.; Ninci, S.; Frazzoli, C.; Dragone, R.; De Rossi, A.; Grasso, G.; Mantovani, A.; Brajon, G. From invention to innovation: Risk analysis to integrate one health technology in the dairy farm. Front. Public Health 2017, 5, 302. [Google Scholar]
- System (Best-Integrated Toxicity (Bio) Sensor’s System for Hazard Analysis and Management in the Food Chain and the Environment) for Environment Diagnostic and Monitoring and Self-Control of Food Chain, Including Primary Production, and Relevant Method for Quick Management of Hazard. Available online: https://patents.google.com/patent/EP2304428B1/en?oq=EP2304428B1 (accessed on 8 August 2021).
- Dragone, R.; Grasso, G.; Frazzoli, C. Amperometric Cytosensor for Studying Mitochondrial Interferences Induced by Plasticizers Bisphenol B and Bisphenol A. Molecules 2020, 25, 5185. [Google Scholar]
- ISO 8692:1989; Water Quality—Fresh Water Algal Growth Inhibition Test with Scenedesmus Subspicatus and Selenastrum Capricornutum. International Organisation for Standardisation: Geneva, Switzerland, 1989; pp. 1–6.
- Tóth, S.Z.; Schansker, G.; Strasser, R.J. A non-invasive assay of the plastoquinone pool redox state based on the OJIP-transient. Photosynth. Res. 2007, 93, 193–203. [Google Scholar]
- Samson, G.; Prášil, O.; Yaakoubd, B. Photochemical and thermal phases of chlorophyll a fluorescence. Photosynthetica 1999, 37, 163–182. [Google Scholar]
- Alric, J.; Lavergne, J.; Rappaport, F. Redox and ATP control of photosynthetic cyclic electron flow in Chlamydomonas reinhardtii (I) aerobic conditions. Biochim. Biophys Acta. (BBA)-Bioenerg. 2010, 1797, 44–51. [Google Scholar]
- Finazzi, G.; Furia, A.; Barbagallo, R.P.; Forti, G. State transitions, cyclic and linear electron transport and photophosphorylation in Chlamydomonas reinhardtii. Biochim. Biophys Acta. (BBA)-Bioenerg. 1999, 1413, 117–129. [Google Scholar]
- Alric, J. The plastoquinone pool, poised for cyclic electron flow? Front. Plant Sci. 2015, 6, 540. [Google Scholar]
- Zhu, X.G.; Baker, N.R.; Desturler, E.; Ort, D.R.; Long, S.P. Chlorophyll a fluorescence induction kinetics in leaves predicted from a model describing each discrete step of excitation energy and electron transfer associated with photosystem II. Planta 2015, 223, 114–133. [Google Scholar]
- Nawrocki, W.J.; Bailleul, B.; Cardol, P.; Rappaport, F.; Wollman, F.A.; Joliot, P. Maximal cyclic electron flow rate is independent of PGRL1 in Chlamydomonas. Biochim. Biophys Acta (BBA)-Bioenerg 2019, 1860, 425–432. [Google Scholar]
- Foyer, C.H.; Neukermans, J.; Queval, G.; Noctor, G.; Harbinson, J. Photosynthetic control of electron transport and the regulation of gene expression. J. Exp. Bot. 2012, 63, 1637–1661. [Google Scholar]
- Finazzi, G.; Rappaport, F.; Furia, A.; Fleischmann, M.; Rochaix, J.D.; Zito, F.; Forti, G. Involvement of state transitions in the switch between linear and cyclic electron flow in Chlamydomonas reinhardtii. EMBO Rep. 2002, 3, 280–285. [Google Scholar]
- Alric, J.; Johnson, X. Alternative electron transport pathways in photosynthesis: A confluence of regulation. Curr. Opin. Plant Biol. 2017, 37, 78–86. [Google Scholar]
- Nestler, H.; Groh, K.J.; Schönenberger, R.; Eggen, R.I.; Suter, M.J.F. Linking proteome responses with physiological and biochemical effects in herbicide-exposed Chlamydomonas reinhardtii. J. Proteom. 2012, 75, 5370–5385. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grasso, G.; Cocco, G.; Zane, D.; Frazzoli, C.; Dragone, R. Microalgae-Based Fluorimetric Bioassays for Studying Interferences on Photosynthesis Induced by Environmentally Relevant Concentrations of the Herbicide Diuron. Biosensors 2022, 12, 67. https://doi.org/10.3390/bios12020067
Grasso G, Cocco G, Zane D, Frazzoli C, Dragone R. Microalgae-Based Fluorimetric Bioassays for Studying Interferences on Photosynthesis Induced by Environmentally Relevant Concentrations of the Herbicide Diuron. Biosensors. 2022; 12(2):67. https://doi.org/10.3390/bios12020067
Chicago/Turabian StyleGrasso, Gerardo, Giulia Cocco, Daniela Zane, Chiara Frazzoli, and Roberto Dragone. 2022. "Microalgae-Based Fluorimetric Bioassays for Studying Interferences on Photosynthesis Induced by Environmentally Relevant Concentrations of the Herbicide Diuron" Biosensors 12, no. 2: 67. https://doi.org/10.3390/bios12020067
APA StyleGrasso, G., Cocco, G., Zane, D., Frazzoli, C., & Dragone, R. (2022). Microalgae-Based Fluorimetric Bioassays for Studying Interferences on Photosynthesis Induced by Environmentally Relevant Concentrations of the Herbicide Diuron. Biosensors, 12(2), 67. https://doi.org/10.3390/bios12020067