Yeast-Based Biosensors: Current Applications and New Developments


Abstract
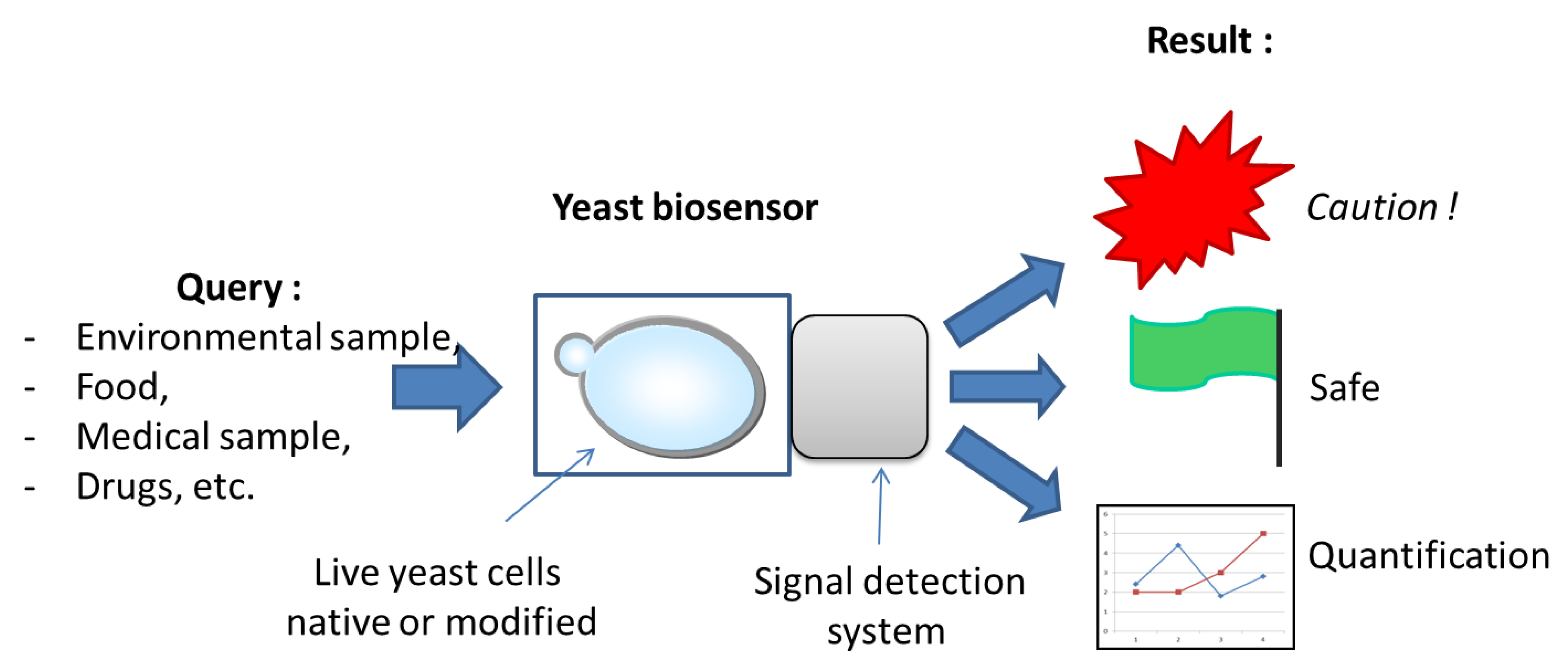
1. Introduction
2. Current Applications
2.1. Environment
2.2. Medical Domain/Health
2.2.1. Detection of Pathogens
2.2.2. Drug Discovery
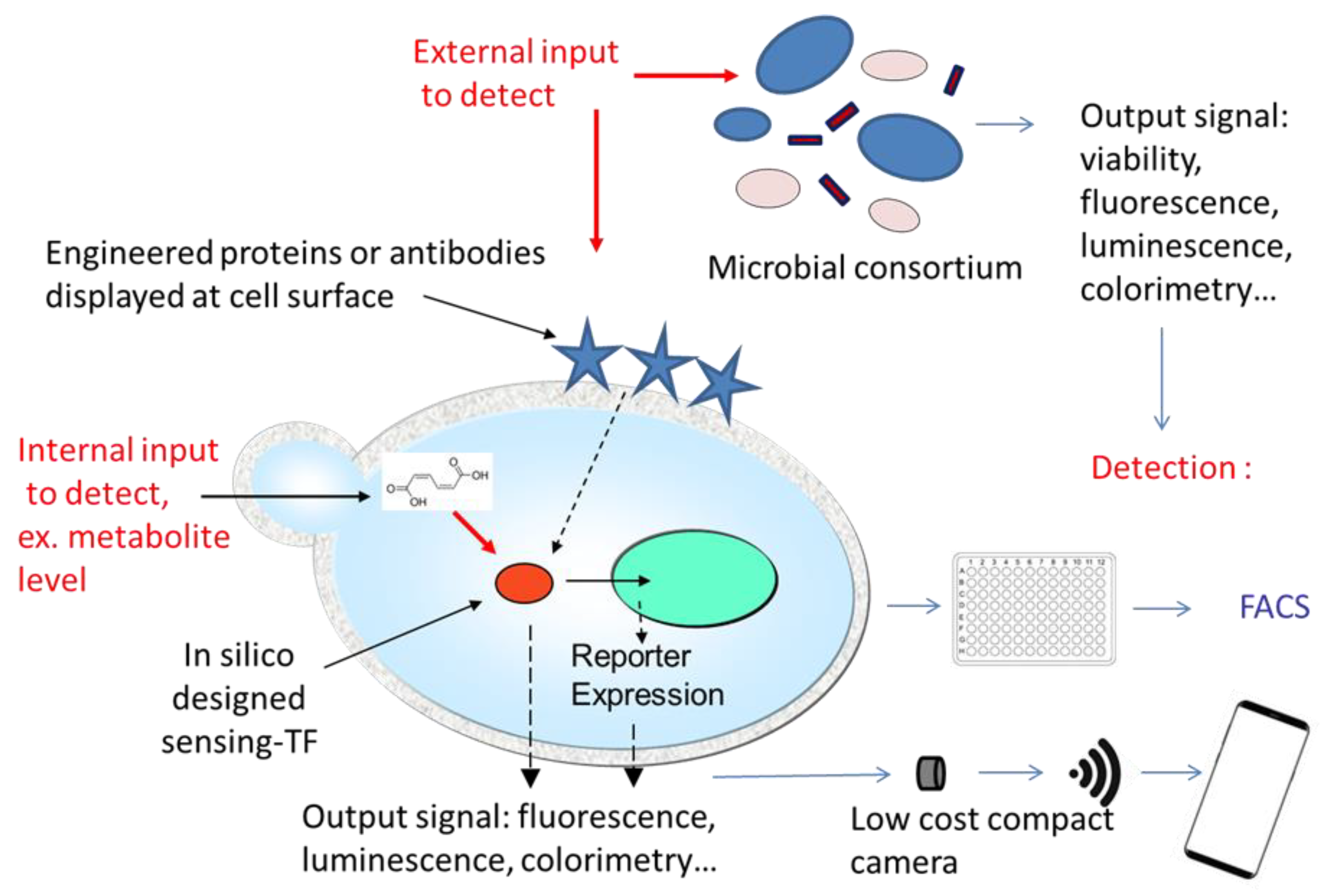
3. Current and Future Developments
3.1. Metabolic Biosensors
3.2. Multi-Strain Biosensors
3.3. Technological Developments
3.4. In Silico Design
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Gutiérrez, J.C.; Amaro, F.; Martín-González, A. Heavy metal whole-cell biosensors using eukaryotic microorganisms: An updated critical review. Front. Microbiol. 2015, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Belkin, S. Microbial whole-cell sensing systems of environmental pollutants. Curr. Opin. Microbiol. 2003, 6, 206–212. [Google Scholar] [CrossRef]
- Han, L.; Zhao, Y.; Cui, S.; Liang, B. Redesigning of Microbial Cell Surface and Its Application to Whole-Cell Biocatalysis and Biosensors. Appl. Biochem. Biotechnol. 2018, 185, 396–418. [Google Scholar] [CrossRef] [PubMed]
- King, J.M.; Digrazia, P.M.; Applegate, B.; Burlage, R.; Sanseverino, J.; Dunbar, P.; Larimer, F.; Sayler, G.S. Rapid, sensitive bioluminescent reporter technology for naphthalene exposure and biodegradation. Science 1990, 249, 778–781. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, R.M.; Keenan, P. The eukaryote alternative: Advantages of using yeasts in place of bacteria in microbial biosensor development. Biotechnol. Bioprocess Eng. 2000, 5, 387–394. [Google Scholar] [CrossRef]
- Henry, S.A.; Patton-Vogt, J.L. Genetic regulation of phospholipid metabolism: Yeast as a model eukaryote. Prog. Nucleic Acid Res. Mol. Biol. 1998, 61, 133–179. [Google Scholar]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 genes. Science 1996, 274, 563–567. [Google Scholar] [CrossRef]
- Välimaa, A.-L.; Kivistö, A.; Virta, M.; Karp, M. Real-time Monitoring of Non-specific Toxicity Using a Saccharomyces cerevisiae Reporter System. Sensors 2008, 8, 6433–6447. [Google Scholar] [CrossRef]
- Ernst, R.; Klemm, R.; Schmitt, L.; Kuchler, K. Yeast ATP-Binding Cassette Transporters: Cellular Cleaning Pumps. In Methods in Enzymology; Elsevier: Amsterdam, Netherlands, 2005; Volume 400, pp. 460–484. ISBN 978-0-12-182805-9. [Google Scholar]
- Hollis, R.P.; Killham, K.; Glover, L.A. Design and Application of a Biosensor for Monitoring Toxicity of Compounds to Eukaryotes. Appl. Environ. Microbiol. 2000, 66, 1676–1679. [Google Scholar] [CrossRef]
- Routledge, E.J.; Sumpter, J.P. Estrogenic activity of surfactants and some of their degradation products assessed using a recombinant yeast screen. Environ. Toxicol. Chem. 1996, 15, 241–248. [Google Scholar] [CrossRef]
- García-Reyero, N.; Grau, E.; Castillo, M.; De Alda, M.J.L.; Barceló, D.; Piña, B. Monitoring of endocrine disruptors in surface waters by the yeast recombinant assay. Environ. Toxicol. Chem. 2001, 20, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Bovee, T.F.; Helsdingen, R.J.; Koks, P.D.; Kuiper, H.A.; Hoogenboom, R.L.A.; Keijer, J. Development of a rapid yeast estrogen bioassay, based on the expression of green fluorescent protein. Gene 2004, 325, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Pham, H.T.M.; Kunath, K.; Gehrmann, L.; Giersberg, M.; Tuerk, J.; Uhlig, S.; Hanke, G.; Simon, K.; Baronian, K.; Kunze, G. Application of modified Arxula adeninivorans yeast cells in an online biosensor for the detection of estrogenic compounds in wastewater samples. Sens. Actuators B Chem. 2013, 185, 628–637. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, Y.S.; Kwon, H.B.; Lee, K. Novel yeast bioassay system for detection of androgenic and antiandrogenic compounds. Toxicol. In Vitro 2003, 17, 237–244. [Google Scholar] [CrossRef]
- Ogawa, M.; Yamaji, R.; Mitani, T.; Murata, Y.; Nakao, M.; Harada, N.; Nakano, Y.; Inui, H. A yeast bioassay for androgenic and anti-androgenic compounds based on the NH2- and COOH-terminal interaction of androgen receptor. Biosci. Biotechnol. Biochem. 2010, 74, 1965–1968. [Google Scholar] [CrossRef]
- Pham, H.T.M.; Giersberg, M.; Gehrmann, L.; Hettwer, K.; Tuerk, J.; Uhlig, S.; Hanke, G.; Weisswange, P.; Simon, K.; Baronian, K.; et al. The determination of pharmaceuticals in wastewater using a recombinant Arxula adeninivorans whole cell biosensor. Sens. Actuators B Chem. 2015, 211, 439–448. [Google Scholar] [CrossRef]
- Pham, H.T.M.; Chamas, A.; Nieter, A.; Giersberg, M.; Rutten, T.; Gehrmann, L.; Hettwer, K.; Tuerk, J.; Uhlig, S.; Simon, K.; et al. Determination of glucocorticoids using photometric (A-YGS) and spectrofluorometric (A-YGFS) bioassays based on modified Arxula adeninivorans cells: Applications in environmental analysis. Sens. Actuators B Chem. 2016, 223, 540–549. [Google Scholar] [CrossRef]
- Schappert, K.T.; Khachatourians, G.G. A yeast bioassay for T-2 toxin. J. Microbiol. Methods 1984, 3, 43–46. [Google Scholar] [CrossRef]
- Engler, K.H.; Coker, R.; Evans, I.H. A novel colorimetric yeast bioassay for detecting trichothecene mycotoxins. J. Microbiol. Methods 1999, 35, 207–218. [Google Scholar] [CrossRef]
- Mitterbauer, R.; Weindorfer, H.; Safaie, N.; Krska, R.; Lemmens, M.; Ruckenbauer, P.; Kuchler, K.; Adam, G. A Sensitive and Inexpensive Yeast Bioassay for the Mycotoxin Zearalenone and Other Compounds with Estrogenic Activity. Appl. Environ. Microbiol. 2003, 69, 805–811. [Google Scholar] [CrossRef]
- Lehmann, M.; Riedel, K.; Adler, K.; Kunze, G. Amperometric measurement of copper ions with a deputy substrate using a novel Saccharomyces cerevisiae sensor. Biosens. Bioelectron. 2000, 15, 211–219. [Google Scholar] [CrossRef]
- Shetty, R.S.; Deo, S.K.; Liu, Y.; Daunert, S. Fluorescence-based sensing system for copper using genetically engineered living yeast cells. Biotechnol. Bioeng. 2004, 88, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Roda, A.; Roda, B.; Cevenini, L.; Michelini, E.; Mezzanotte, L.; Reschiglian, P.; Hakkila, K.; Virta, M. Analytical strategies for improving the robustness and reproducibility of bioluminescent microbial bioreporters. Anal. Bioanal. Chem. 2011, 401, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Vopálenská, I.; Váchová, L.; Palková, Z. New biosensor for detection of copper ions in water based on immobilized genetically modified yeast cells. Biosens. Bioelectron. 2015, 72, 160–167. [Google Scholar] [CrossRef]
- Park, J.-N.; Sohn, M.J.; Oh, D.-B.; Kwon, O.; Rhee, S.K.; Hur, C.-G.; Lee, S.Y.; Gellissen, G.; Kang, H.A. Identification of the Cadmium-Inducible Hansenula polymorpha SEO1 Gene Promoter by Transcriptome Analysis and Its Application to Whole-Cell Heavy-Metal Detection Systems. Appl. Environ. Microbiol. 2007, 73, 5990–6000. [Google Scholar] [CrossRef]
- Richter, I.; Fidler, A.E. Detection of marine microalgal biotoxins using bioassays based on functional expression of tunicate xenobiotic receptors in yeast. Toxicon 2015, 95, 13–22. [Google Scholar] [CrossRef]
- Martin-Yken, H.; Gironde, C.; Derick, S.; Darius, H.T.; Furger, C.; Laurent, D.; Chinain, M. Ciguatoxins activate the Calcineurin signalling pathway in Yeasts: Potential for development of an alternative detection tool? Environ. Res. 2018, 162, 144–151. [Google Scholar] [CrossRef]
- Hikuma, M.; Suzuki, H.; Yasuda, T.; Karube, I.; Suzuki, S. Amperometric estimation of BOD using living immobilised yeasts. Eur. J. Appl. Microbiol. Biotechnol. 1979, 8, 289–297. [Google Scholar] [CrossRef]
- Yudina, N.Y.; Arlyapov, V.A.; Chepurnova, M.A.; Alferov, S.V.; Reshetilov, A.N. A yeast co-culture-based biosensor for determination of waste water contamination levels. Enzyme Microb. Technol. 2015, 78, 46–53. [Google Scholar] [CrossRef]
- Nakamura, H.; Kobayashi, S.; Hirata, Y.; Suzuki, K.; Mogi, Y.; Karube, I. A spectrophotometric biochemical oxygen demand determination method using 2,6-dichlorophenolindophenol as the redox color indicator and the eukaryote Saccharomyces cerevisiae. Anal. Biochem. 2007, 369, 168–174. [Google Scholar] [CrossRef]
- Venkatesh, A.G.; Sun, A.; Brickner, H.; Looney, D.; Hall, D.A.; Aronoff-Spencer, E. Yeast dual-affinity biobricks: Progress towards renewable whole-cell biosensors. Biosens. Bioelectron. 2015, 70, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Ostrov, N.; Jimenez, M.; Billerbeck, S.; Brisbois, J.; Matragrano, J.; Ager, A.; Cornish, V.W. A modular yeast biosensor for low-cost point-of-care pathogen detection. Sci. Adv. 2017, 3, e1603221. [Google Scholar] [CrossRef] [PubMed]
- Fleming, L.; Depledge, M.; McDonough, N.; White, M.; Pahl, S.; Austen, M.; Goksoyr, A.; Solo-Gabriele, H.; Stegeman, J. The Oceans and Human Health; Oxford University Press: Oxford, UK, 2015; Volume 1. [Google Scholar]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.G.; Glasser, N.R.; Palecek, S.P. The utilization of a Saccharomyces cerevisiae HUG1P-GFP promoter–reporter construct for the selective detection of DNA damage. Mutat. Res. Toxicol. Environ. Mutagen. 2007, 633, 21–34. [Google Scholar] [CrossRef]
- Bui, V.N.; Nguyen, T.T.H.; Mai, C.T.; Bettarel, Y.; Hoang, T.Y.; Trinh, T.T.L.; Truong, N.H.; Chu, H.H.; Nguyen, V.T.T.; Nguyen, H.D.; et al. Procarcinogens—Determination and Evaluation by Yeast-Based Biosensor Transformed with Plasmids Incorporating RAD54 Reporter Construct and Cytochrome P450 Genes. PLoS ONE 2016, 11, e0168721. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Acero, T.; Rodríguez-Escudero, I.; Vicente, F.; Monteiro, M.C.; Tormo, J.R.; Cantizani, J.; Molina, M.; Cid, V.J. A Yeast-Based In Vivo Bioassay to Screen for Class I Phosphatidylinositol 3-Kinase Specific Inhibitors. J. Biomol. Screen. 2012, 17, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Diehl, B.; Hoffmann, T.M.; Mueller, N.C.; Burkhart, J.L.; Kazmaier, U.; Schmitt, M.J. Novel Yeast Bioassay for High-Throughput Screening of Matrix Metalloproteinase Inhibitors. Appl. Environ. Microbiol. 2011, 77, 8573–8577. [Google Scholar] [CrossRef][Green Version]
- Mohamad, U.H.; Hamid, U.M.A.; Abdullah, M.F.F. Development of a yeast bioassay for the screening of anti-malarial compounds with artemisinin-like activities. In Proceedings of the 2012 IEEE Colloquium on Humanities, Science and Engineering (CHUSER), Kota Kinabalu, Malaysia, 3–4 December 2012; pp. 122–125. [Google Scholar]
- Cottier, V.; Barberis, A.; Lüthi, U. Novel Yeast Cell-Based Assay To Screen for Inhibitors of Human Cytomegalovirus Protease in a High-Throughput Format. Antimicrob. Agents Chemother. 2006, 50, 565–571. [Google Scholar] [CrossRef]
- Jarque, S.; Bittner, M.; Blaha, L.; Hilscherova, K. Yeast Biosensors for Detection of Environmental Pollutants: Current State and Limitations. Trends Biotechnol. 2016, 34, 408–419. [Google Scholar] [CrossRef]
- Middelhoven, W.J.; de Jong, I.M.; de Winter, M. Arxula adeninivorans, a yeast assimilating many nitrogenous and aromatic compounds. Antonie Van Leeuwenhoek 1991, 59, 129–137. [Google Scholar]
- Baronian, K.H.R. The use of yeast and moulds as sensing elements in biosensors. Biosens. Bioelectron. 2004, 19, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Fang, D.; Yu, Y.; Wu, L.; Wang, Y.; Zhi, J. A double-mediator based whole cell electrochemical biosensor for acute biotoxicity assessment of wastewater. Talanta 2017, 167, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Jouanneau, S.; Recoules, L.; Durand, M.J.; Boukabache, A.; Picot, V.; Primault, Y.; Lakel, A.; Sengelin, M.; Barillon, B.; Thouand, G. Methods for assessing biochemical oxygen demand (BOD): A review. Water Res. 2014, 49, 62–82. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.P.F.; Cunha, E.; Azevedo, A.M.O.; Pereira, S.A.P.; Neves, A.F.D.C.; Vilaranda, A.G.; Araujo, A.R.T.S.; Passos, M.L.C.; Pinto, P.C.A.G.; Saraiva, M.L.M.F.S. Microfluidic Chemiluminescence System with Yeast Saccharomyces cerevisiae for Rapid Biochemical Oxygen Demand Measurement. ACS Sustain. Chem. Eng. 2018, 6, 6094–6101. [Google Scholar] [CrossRef]
- Rose, M.; Casadaban, M.J.; Botstein, D. Yeast genes fused to beta-galactosidase in Escherichia coli can be expressed normally in yeast. Proc. Natl. Acad. Sci. USA 1981, 78, 2460–2464. [Google Scholar] [CrossRef]
- Guarente, L.; Ptashne, M. Fusion of Escherichia coli lacZ to the cytochrome c gene of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1981, 78, 2199–2203. [Google Scholar] [CrossRef]
- Brix, R.; Noguerol, T.-N.; Piña, B.; Balaam, J.; Nilsen, A.J.; Tollefsen, K.E.; Levy, W.; Schramm, K.-W.; Barceló, D. Evaluation of the suitability of recombinant yeast-based estrogenicity assays as a pre-screening tool in environmental samples. Environ. Int. 2010, 36, 361–367. [Google Scholar] [CrossRef]
- Balsiger, H.A.; de la Torre, R.; Lee, W.-Y.; Cox, M.B. A Four-Hour Yeast Bioassay for the Direct Measure of Estrogenic Activity in Wastewater without Sample Extraction, Concentration, or Sterilization. Sci. Total Environ. 2010, 408, 1422–1429. [Google Scholar] [CrossRef]
- Sanseverino, J.; Gupta, R.K.; Layton, A.C.; Patterson, S.S.; Ripp, S.A.; Saidak, L.; Simpson, M.L.; Schultz, T.W.; Sayler, G.S. Use of Saccharomyces cerevisiae BLYES Expressing Bacterial Bioluminescence for Rapid, Sensitive Detection of Estrogenic Compounds. Appl. Environ. Microbiol. 2005, 71, 4455–4460. [Google Scholar] [CrossRef]
- Sanseverino, J.; Eldridge, M.L.; Layton, A.C.; Easter, J.P.; Yarbrough, J.; Schultz, T.W.; Sayler, G.S. Screening of Potentially Hormonally Active Chemicals Using Bioluminescent Yeast Bioreporters. Toxicol. Sci. 2009, 107, 122–134. [Google Scholar] [CrossRef]
- Rajasärkkä, J.; Hakkila, K.; Virta, M. Developing a compound-specific receptor for bisphenol a by directed evolution of human estrogen receptor α. Biotechnol. Bioeng. 2011, 108, 2526–2534. [Google Scholar] [CrossRef] [PubMed]
- Di Dea Bergamasco, A.M.; Eldridge, M.; Sanseverino, J.; Sodré, F.F.; Montagner, C.C.; Pescara, I.C.; Jardim, W.F.; de Aragao Umbuzeiro, G. Bioluminescent yeast estrogen assay (BLYES) as a sensitive tool to monitor surface and drinking water for estrogenicity. J. Environ. Monit. 2011, 13, 3288–3293. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Eldridge, M.; Menn, F.; Dykes, T.; Sayler, G. Standardized application of yeast bioluminescent reporters as endocrine disruptor screen for comparative analysis of wastewater effluents from membrane bioreactor and traditional activated sludge. Ecotoxicology 2015, 24, 2088–2099. [Google Scholar] [CrossRef]
- Westlund, P.; Yargeau, V. Investigation of the presence and endocrine activities of pesticides found in wastewater effluent using yeast-based bioassays. Sci. Total Environ. 2017, 607–608, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Roszko, M.Ł.; Kamińska, M.; Szymczyk, K.; Piasecka-Jóźwiak, K.; Chabłowska, B. Endocrine disrupting potency of organic pollutant mixtures isolated from commercial fish oil evaluated in yeast-based bioassays. PLoS ONE 2018, 13, e0197907. [Google Scholar] [CrossRef] [PubMed]
- Rajasärkkä, J.; Koponen, J.; Airaksinen, R.; Kiviranta, H.; Virta, M. Monitoring bisphenol A and estrogenic chemicals in thermal paper with yeast-based bioreporter assay. Anal. Bioanal. Chem. 2014, 406, 5695–5702. [Google Scholar] [CrossRef]
- Gao, G.; Qian, J.; Fang, D.; Yu, Y.; Zhi, J. Development of a mediated whole cell-based electrochemical biosensor for joint toxicity assessment of multi-pollutants using a mixed microbial consortium. Anal. Chim. Acta 2016, 924, 21–28. [Google Scholar] [CrossRef]
- Furger, C. Live Cell Assays: Live Cell Assays: From Research to Regulatory Applications; John Wiley and Sons Inc.: 111 River Street, Hoboken, NJ, USA, 2016; ISBN 978-1-84821-858-1. [Google Scholar]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [Google Scholar] [CrossRef]
- da Silva, E.O.; Bracarense, A.P.F.L.; Oswald, I.P. Mycotoxins and oxidative stress: Where are we? World Mycotoxin J. 2018, 11, 113–134. [Google Scholar] [CrossRef]
- Nguyen, N.-T.; Sekhon, S.S.; Yoon, J.; Kim, Y.-H.; Min, J. Effect of heavy metals, pesticides and pharmaceuticals on yeast’s vacuoles as a biomarker for toxic detection. Mol. Cell. Toxicol. 2017, 13, 287–294. [Google Scholar] [CrossRef]
- Mager, W.H.; Winderickx, J. Yeast as a model for medical and medicinal research. Trends Pharmacol. Sci. 2005, 26, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Parry, J.M. Use of tests in yeasts and fungi in the detection and evaluation of carcinogens. IARC Sci. Publ. 1999, 146, 471–485. [Google Scholar]
- Brennan, R.J.; Schiestl, R.H. Positive responses to carcinogens in the yeast DEL recombination assay are not due to selection of preexisting spontaneous revertants. Mutat. Res. Mol. Mech. Mutagen. 1998, 421, 117–120. [Google Scholar] [CrossRef]
- Brennan, R.J.; Schiestl, R.H. Detecting Carcinogens with the Yeast DEL Assay. In Genetic Recombination; Humana Press: Totowa, NJ, USA, 2004; pp. 111–124. ISBN 978-1-59259-761-1. [Google Scholar]
- Hontzeas, N.; Hafer, K.; Schiestl, R.H. Development of a microtiter plate version of the yeast DEL assay amenable to high-throughput toxicity screening of chemical libraries. Mutat. Res. Toxicol. Environ. Mutagen. 2007, 634, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Lasserre, J.-P.; Dautant, A.; Aiyar, R.S.; Kucharczyk, R.; Glatigny, A.; Tribouillard-Tanvier, D.; Rytka, J.; Blondel, M.; Skoczen, N.; Reynier, P.; et al. Yeast as a system for modeling mitochondrial disease mechanisms and discovering therapies. Dis. Model. Mech. 2015, 8, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Becker, B.; Schnöder, T.; Schmitt, M.J. Yeast Reporter Assay to Identify Cellular Components of Ricin Toxin a Chain Trafficking. Toxins 2016, 8, 366. [Google Scholar] [CrossRef]
- Chernoff, Y.O. Stress and prions: Lessons from the yeast model. FEBS Lett. 2007, 581, 3695–3701. [Google Scholar] [CrossRef]
- Ter-Avanesyan, M.D.; Dagkesamanskaya, A.R.; Kushnirov, V.V.; Smirnov, V.N. The SUP35 omnipotent suppressor gene is involved in the maintenance of the non-Mendelian determinant [psi+] in the yeast Saccharomyces cerevisiae. Genetics 1994, 137, 671–676. [Google Scholar]
- Bach, S.; Talarek, N.; Andrieu, T.; Vierfond, J.-M.; Mettey, Y.; Galons, H.; Dormont, D.; Meijer, L.; Cullin, C.; Blondel, M. Isolation of drugs active against mammalian prions using a yeast-based screening assay. Nat. Biotechnol. 2003, 21, 1075–1081. [Google Scholar] [CrossRef]
- Bach, S.; Tribouillard, D.; Talarek, N.; Desban, N.; Gug, F.; Galons, H.; Blondel, M. A yeast-based assay to isolate drugs active against mammalian prions. Methods 2006, 39, 72–77. [Google Scholar] [CrossRef]
- Heinisch, J.J. Baker’s yeast as a tool for the development of antifungal kinase inhibitors--targeting protein kinase C and the cell integrity pathway. Biochim. Biophys. Acta 2005, 1754, 171–182. [Google Scholar] [CrossRef]
- D’Ambrosio, V.; Jensen, M.K. Lighting up yeast cell factories by transcription factor-based biosensors. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef]
- Jensen, M.K.; Keasling, J.D. Recent applications of synthetic biology tools for yeast metabolic engineering. FEMS Yeast Res. 2015, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- De Paepe, B.; Peters, G.; Coussement, P.; Maertens, J.; De Mey, M. Tailor-made transcriptional biosensors for optimizing microbial cell factories. J. Ind. Microbiol. Biotechnol. 2017, 44, 623–645. [Google Scholar] [CrossRef] [PubMed]
- Skjoedt, M.L.; Snoek, T.; Kildegaard, K.R.; Arsovska, D.; Eichenberger, M.; Goedecke, T.J.; Rajkumar, A.S.; Zhang, J.; Kristensen, M.; Lehka, B.J.; et al. Engineering prokaryotic transcriptional activators as metabolite biosensors in yeast. Nat. Chem. Biol. 2016, 12, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Ambri, F.; Snoek, T.; Skjoedt, M.L.; Jensen, M.K.; Keasling, J.D. Design, Engineering, and Characterization of Prokaryotic Ligand-Binding Transcriptional Activators as Biosensors in Yeast. In Synthetic Metabolic Pathways; Jensen, M.K., Keasling, J.D., Eds.; Springer: New York, NY, USA, 2018; Volume 1671, pp. 269–290. ISBN 978-1-4939-7294-4. [Google Scholar]
- Snoek, T.; Romero-Suarez, D.; Zhang, J.; Ambri, F.; Skjoedt, M.L.; Sudarsan, S.; Jensen, M.K.; Keasling, J.D. An Orthogonal and pH-Tunable Sensor-Selector for Muconic Acid Biosynthesis in Yeast. ACS Synth. Biol. 2018, 7, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Si, T.; Wang, M.; Zhao, H. Development of a Synthetic Malonyl-CoA Sensor in Saccharomyces cerevisiae for Intracellular Metabolite Monitoring and Genetic Screening. ACS Synth. Biol. 2015, 4, 1308–1315. [Google Scholar] [CrossRef]
- Zhang, J.; Sonnenschein, N.; Pihl, T.P.B.; Pedersen, K.R.; Jensen, M.K.; Keasling, J.D. Engineering an NADPH/NADP + Redox Biosensor in Yeast. ACS Synth. Biol. 2016, 5, 1546–1556. [Google Scholar] [CrossRef]
- Liu, L.; Deng, L.; Yong, D.; Dong, S. Native biofilm cultured under controllable condition and used in mediated method for BOD measurement. Talanta 2011, 84, 895–899. [Google Scholar] [CrossRef]
- Rajpoot, K. Recent Advances and Applications of Biosensors in Novel Technology. Biosens. J. 2017, 6, 1–12. [Google Scholar] [CrossRef]
- Tian, Y.; Lu, Y.; Xu, X.; Wang, C.; Zhou, T.; Li, X. Construction and comparison of yeast whole-cell biosensors regulated by two RAD54 promoters capable of detecting genotoxic compounds. Toxicol. Mech. Methods 2017, 27, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Cevenini, L.; Calabretta, M.M.; Tarantino, G.; Michelini, E.; Roda, A. Smartphone-interfaced 3D printed toxicity biosensor integrating bioluminescent “sentinel cells”. Sens. Actuators B Chem. 2016, 225, 249–257. [Google Scholar] [CrossRef]
- Lopreside, A.; Calabretta, M.M.; Montali, L.; Ferri, M.; Tassoni, A.; Branchini, B.R.; Southworth, T.; D’Elia, M.; Roda, A.; Michelini, E. Prêt-à-porter nanoYESα and nanoYESβ bioluminescent cell biosensors for ultrarapid and sensitive screening of endocrine-disrupting chemicals. Anal. Bioanal. Chem. 2019, 411, 4937–4949. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Neuta, I.; Neumann, F.; Brightmeyer, J.; Ba Tis, T.; Madaboosi, N.; Wei, Q.; Ozcan, A.; Nilsson, M. Smartphone-based clinical diagnostics: Towards democratization of evidence-based health care. J. Intern. Med. 2019, 285, 19–39. [Google Scholar] [CrossRef] [PubMed]
- Aronoff-Spencer, E.; Venkatesh, A.G.; Sun, A.; Brickner, H.; Looney, D.; Hall, D.A. Detection of Hepatitis C core antibody by dual-affinity yeast chimera and smartphone-based electrochemical sensing. Biosens. Bioelectron. 2016, 86, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Shaw, W.M.; Yamauchi, H.; Mead, J.; Gowers, G.-O.F.; Bell, D.J.; Öling, D.; Larsson, N.; Wigglesworth, M.; Ladds, G.; Ellis, T. Engineering a Model Cell for Rational Tuning of GPCR Signaling. Cell 2019, 177, 782–796. [Google Scholar] [CrossRef]
- Vavitsas, K. Turning G protein-coupled receptors into tunable biosensors. Synth. Biol. 2019, 4, ysz011. [Google Scholar] [CrossRef]
- Arai, R. Hierarchical design of artificial proteins and complexes toward synthetic structural biology. Biophys. Rev. 2018, 10, 391–410. [Google Scholar] [CrossRef]
- Kobayashi, N.; Inano, K.; Sasahara, K.; Sato, T.; Miyazawa, K.; Fukuma, T.; Hecht, M.H.; Song, C.; Murata, K.; Arai, R. Self-Assembling Supramolecular Nanostructures Constructed from de Novo Extender Protein Nanobuilding Blocks. ACS Synth. Biol. 2018, 7, 1381–1394. [Google Scholar] [CrossRef]
- Castaño-Cerezo, S.; Fournié, M.; Urban, P.; Faulon, J.-L.; Truan, G. Development of a Biosensor for Detection of Benzoic Acid Derivatives in Saccharomyces cerevisiae. Front. Bioeng. Biotechnol. 2020, 7, 372. [Google Scholar] [CrossRef]
- Donnelly, A.E.; Murphy, G.S.; Digianantonio, K.M.; Hecht, M.H. A de novo enzyme catalyzes a life-sustaining reaction in Escherichia coli. Nat. Chem. Biol. 2018, 14, 253–255. [Google Scholar] [CrossRef] [PubMed]
- Foo, J.L.; Chang, M.W. Synthetic yeast genome reveals its versatility. Nature 2018, 557, 647–648. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Detected Coumponds | Yeast Species | Type of Response (LoD or EC50 if Available) | Detection (Reporter Gene) | References |
|---|---|---|---|---|
| Environment: | ||||
| Coumpounds "toxic to eukaryotic cells" (all types) | Saccharomyces cerevisiae | Yes/No | Luminescence (Luc), viability decrease. | (Hollis et al., 2000) [10] |
| Estrogenic coumponds (Endocrine Disruptors) | ||||
| Saccharomyces cerevisiae | Yes/No (2 ng/L) | Colorimetry (LacZ) | (Routledge and Sumpter, 1996) [11] | |
| Saccharomyces cerevisiae | Quantification (20 ng/L) | Fluorescence (LacZ) | (García-Reyero et al., 2001) [12] | |
| Saccharomyces cerevisiae | Quantification (0.4 nM) | Fluorescence (yEGFP) | (Bovee et al., 2004) [13] | |
| Arxula adeninivorans | Quantification (2 ng/L) | Amperometry or biochemistry (phyK) | (Pham et al., 2013) [14] | |
| Androgenic and Anti-androgenic compounds | Saccharomyces cerevisiae | Quantification (15 nM for Testosterone) | Two-hybrids System, (LacZ). | (Lee et al, 2003) [15] |
| Saccharomyces cerevisiae | Quantification (5 nM) | Two-hybrids System, (GFP). | (Ogawa et al., 2010) [16] | |
| Glucocorticoids (cortisol, corticosterone) | Arxula adeninivorans | Quantification (0.3 μM) | Amperometry or biochemistry (phyK) | (Pham et al., 2016) [17] |
| Pharmaceuticals (omeprazole, lansoprazole) | Arxula adeninivorans | Quantification (95 μg/L) | Amperometry or biochemistry (phyK) | (Pham et al., 2015) [18] |
| Mycotoxins: | ||||
| T-2 toxin and other trichothecenes such as verrucarin A | Kluyveromyces fragilis | Yes/No | Growth inhibition (disk halo) | (Schappert and Khachatourians, 1984) [19] |
| Trichothecene mycotoxins | Kluyveromyces marxianus | Quantification (1 pg/L) | Colorimetry (LacZ) | (Engler et al., 1999) [20] |
| Mycotoxin Zearalenone, and other compounds with estrogenic activity | Saccharomyces cerevisiae | Quantification (1 μg/L) | Metabolic construct | (Mitterbauer et al., 2003) [21] |
| Heavy metals: | ||||
| Saccharomyces cerevisiae | Quantification (0.5 mM Cu2+) | Amperometry (LacZ). | (Lehmann et al., 2000) [22] | |
| Saccharomyces cerevisiae | Quantification (5 × 10-7M Cu2+) | Fluorescence (GFP) | (Shetty et al., 2004) [23] | |
| Saccharomyces cerevisiae | Quantification (5 × 10-7M Cu2+) | Luminescnce (Luc) | (Roda et al., 2011) [24] | |
| Saccharomyces cerevisiae | Quantification (1 μM Cu2+) | Colorimetry (ADE2) | (Vopálenská et al., 2015) [25] | |
| Cadnium, Arsenic. | Hansenula polymorpha | Quantification (1 mM Cd) | Fluorescence (GFP) | (Park et al., 2007) [26] |
| Marine toxins: | ||||
| Okadaic acid, pectenotoxin-11, portimine | Saccharomyces cerevisiae | Quantification (19 nM OA) | Colorimetry (LacZ) | (Richter and Fidler, 2015) [27] |
| Ciguatoxins | Saccharomyces cerevisiae | Quantification (0.1 ng/L PCTX3C) | Colorimetry or fluorescence (LacZ) | (Martin-Yken et al., 2018) [28] |
| Biological Organics (BOD measure): | Trichosporon cutaneum | Quantification (3 mg/L) | Amperometry | (Hikuma et al., 1979) [29] |
| Arxula adeninivorans and other yeast species | Quantification (2.4 mg/L) | Cellular growth | (Yudina et al., 2015) [30] | |
| Saccharomyces cerevisiae | Quantification (2 mg/L) | Spectrophotometry | (Nakamura, 2007) [31] | |
| Medical Domain: | ||||
| Pathogens (of any type) | Saccharomyces cerevisiae | Quantification (nM range) | SPR (antigen cell surface display) | (Venkatesh et al., 2015) [32] |
| Fungal pathogens | Saccharomyces cerevisiae | Yes/No (nM range) | Colorimetry, Engineered GPCR | (Ostrov et al., 2017) [33] |
| Carcinogens, genotoxics: | Saccharomyces cerevisiae | Quantification (mg/mL range) | Reversion frequency (DEL assay) | (Brennan and Schiestl, 1998, 2004) [34,35] |
| Saccharomyces cerevisiae | Quantification (variable) | Fluorescence (GFP) | (Benton et al., 2007) [36] | |
| Pro-carcinogens | Saccharomyces cerevisiae | Quantification (μg/mL range) | CPR-CYP and RAD54-GFP expression | (Bui et al., 2016) [37] |
| PI3K inhibitors (oncogenesis related screen) | Saccharomyces cerevisiae | Quantification (μM range) | Reconstituted PI3K pathway | (Fernández-Acero et al., 2012) [38] |
| Screens: | ||||
| for Matrix Metalloproteinases (MMPs) inhibitors (anticancer) | Pichia pastoris | Quantification (nM range) | Cell surface expression | (Diehl et al., 2011) [39] |
| for Anti-Malarial Compounds with artemisinin-like activities | Saccharomyces cerevisiae | Yes/No (μM range) | Growth inhibition | (Mohamad et al., 2012) [40] |
| for Inhibitors of Human Cytomegalovirus Protease | Saccharomyces cerevisiae | Quantification (μM range) | Target-specific HTS system | (Cottier et al., 2006) [41] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Yken, H. Yeast-Based Biosensors: Current Applications and New Developments. Biosensors 2020, 10, 51. https://doi.org/10.3390/bios10050051
Martin-Yken H. Yeast-Based Biosensors: Current Applications and New Developments. Biosensors. 2020; 10(5):51. https://doi.org/10.3390/bios10050051
Chicago/Turabian StyleMartin-Yken, Helene. 2020. "Yeast-Based Biosensors: Current Applications and New Developments" Biosensors 10, no. 5: 51. https://doi.org/10.3390/bios10050051
APA StyleMartin-Yken, H. (2020). Yeast-Based Biosensors: Current Applications and New Developments. Biosensors, 10(5), 51. https://doi.org/10.3390/bios10050051