Low Dose Carbon Black Nanoparticle Exposure Does Not Aggravate Allergic Airway Inflammation in Mice Irrespective of the Presence of Surface Polycyclic Aromatic Hydrocarbons


Abstract
1. Introduction
2. Results
2.1. Characterization of CBNP and Their Suspensions
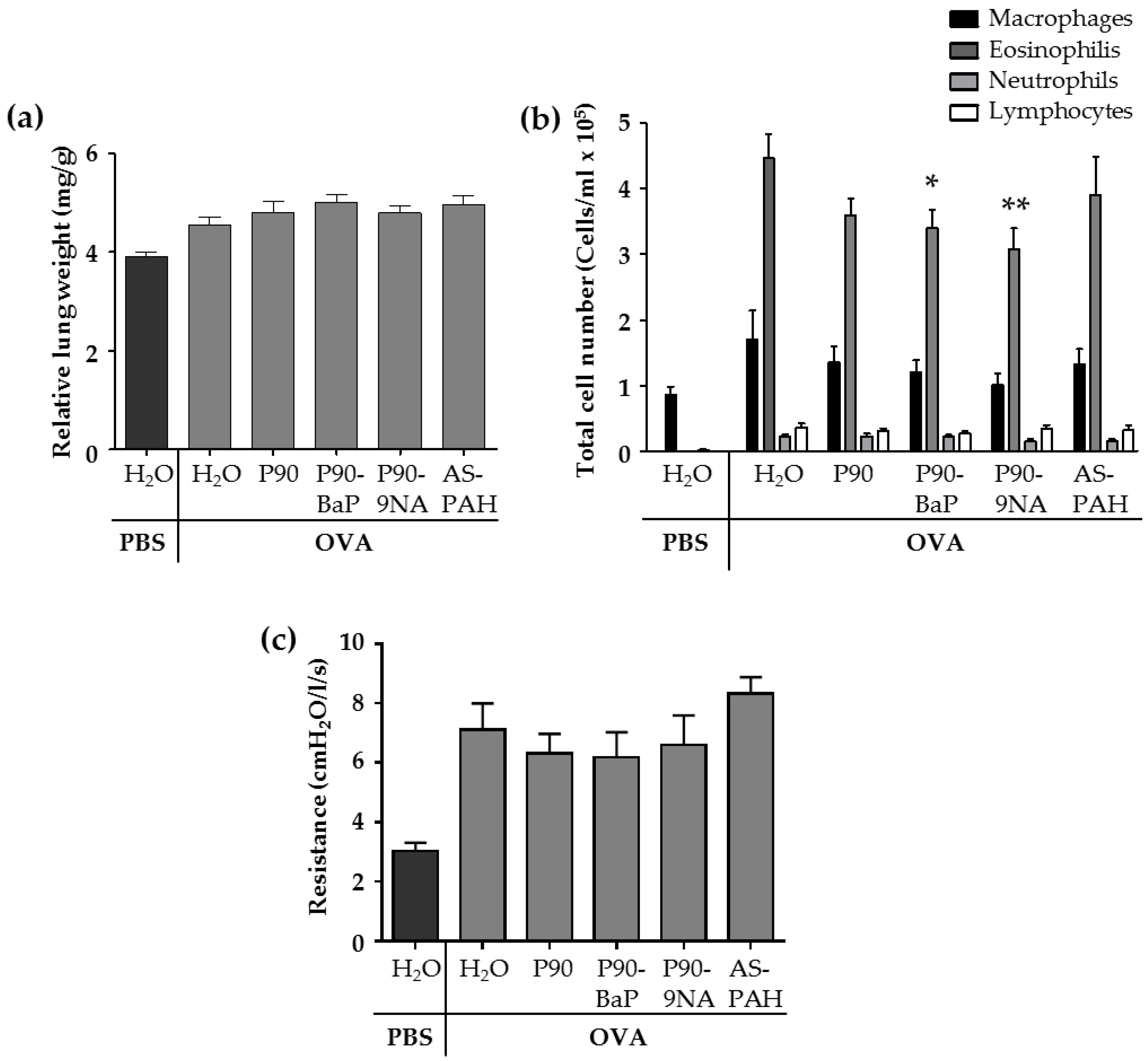
2.2. CBNP Did Not Increase Inflammatory Parameters in the Lungs of OVA-Treated Mice
2.3. CBNP Did Not Increase the mRNA Expression of Inflammatory-Relevant Markers in the Tracheal Epithelial Cells and Intrapulmonary Airways of OVA-Treated Mice
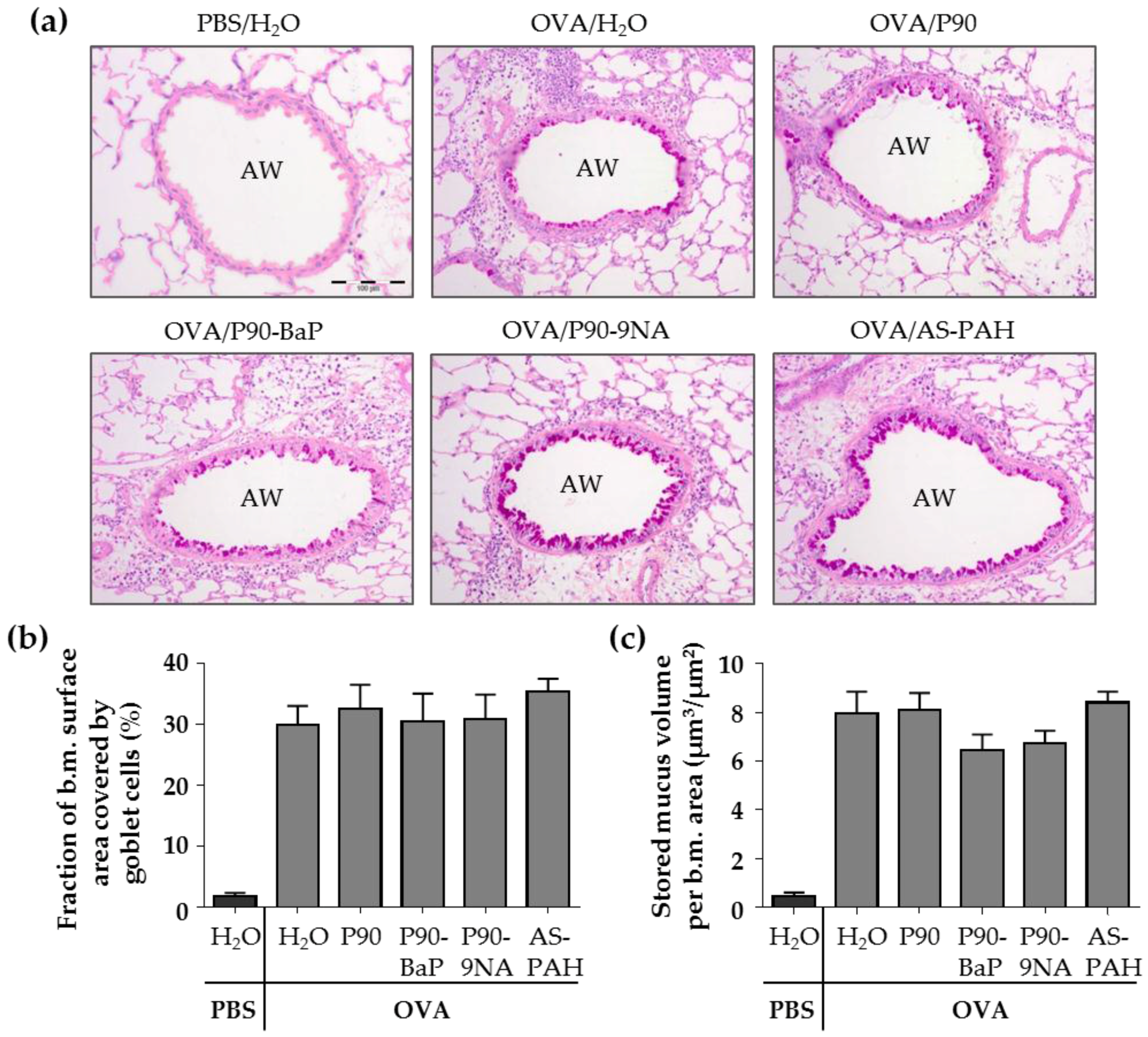
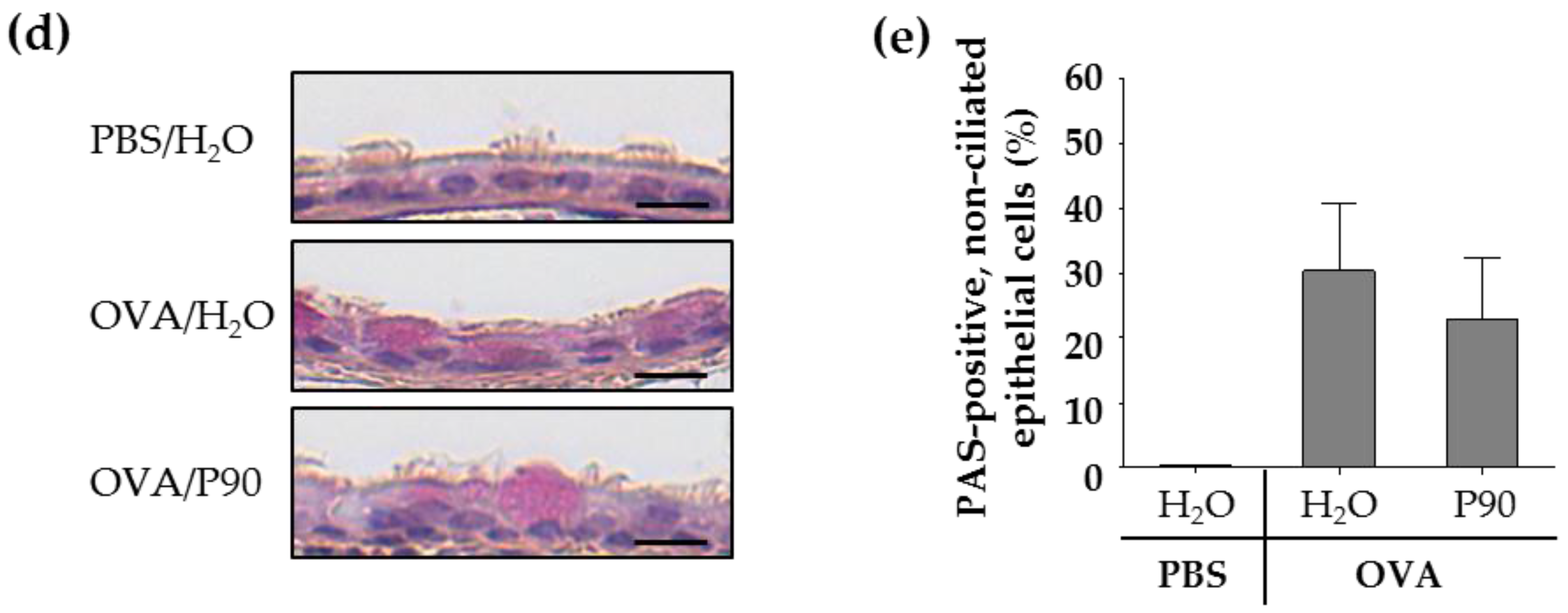
2.4. CBNP Did Not Increase the Number of Airway Epithelial Goblet Cells in OVA-Treated Mice
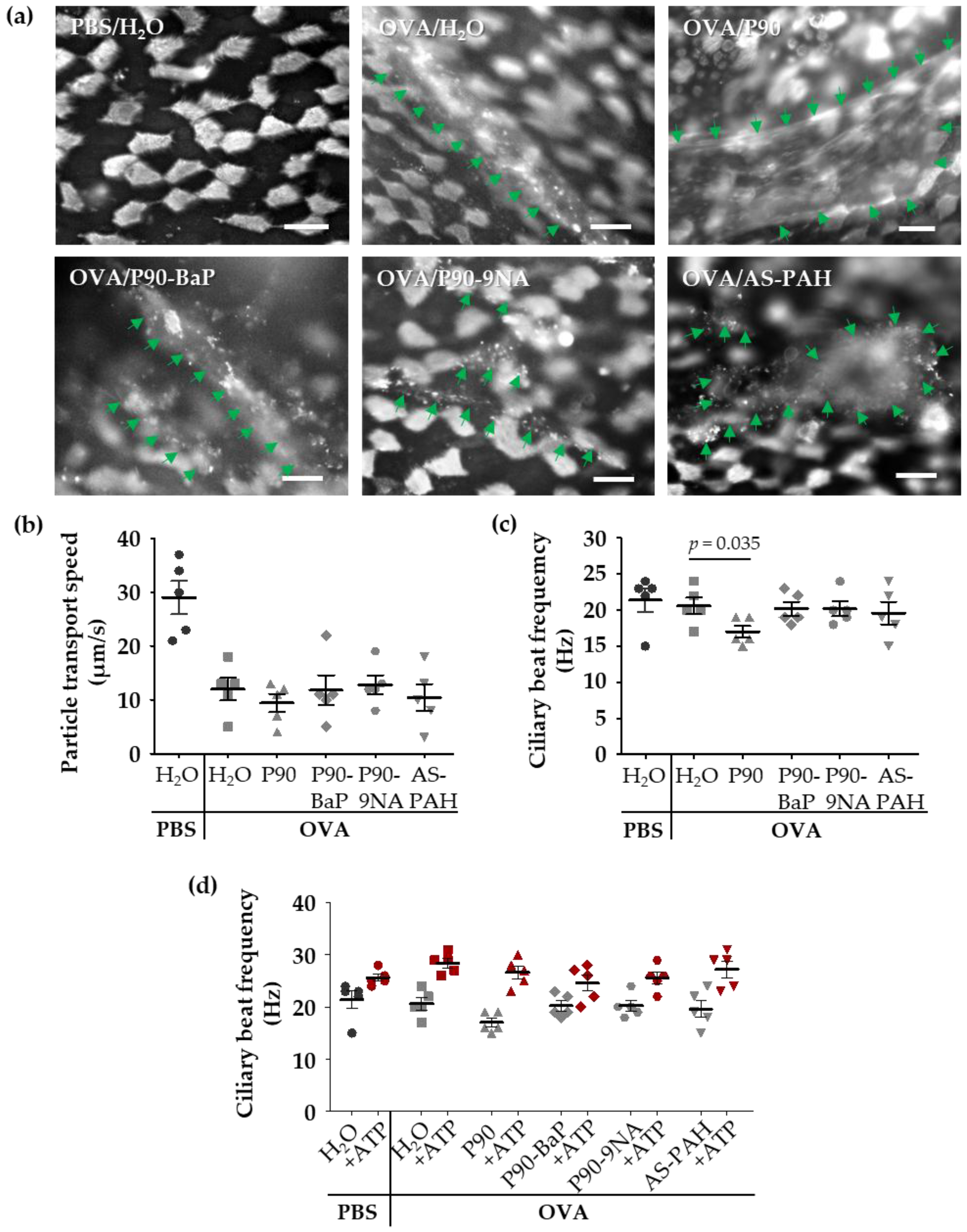
2.5. P90-Induced Mucus Release Affects the Ciliary Beat Frequency in Tracheal Epithelial Cells of OVA-Treated Mice
3. Discussion
4. Materials and Methods
4.1. Particle Preparation and Characterization
4.2. Preparation and Characterization of CBNP Suspension
4.3. Animals
4.4. Animal Treatment Protocol
4.5. Differential Cell Count in Bronchoalveolar Lavage (BAL) Fluid
4.6. Preparation of Airways
4.7. RNA Isolation and Quantitative RT-PCR
4.8. Measurement of the Protein Levels in the BAL Fluid
4.9. PAS Staining
4.10. Mucus Staining on the Vital Tracheal Epithelium
4.11. Whole Mount Analysis of Dead Cells
4.12. Particle Transport Speed and Ciliary Beat Frequency
4.13. Assessment of Airway Responsiveness to Methacholine
4.14. Preparation of Trachea Samples for Scanning Electron Microscopy
4.15. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Global Burden of Disease Study 2013 Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 301 acute and chronic diseases and injuries in 188 countries, 1990–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2015, 386, 743–800. [Google Scholar]
- Fanta, C.H. Asthma. N. Engl. J. Med. 2009, 360, 1002–1014. [Google Scholar] [CrossRef] [PubMed]
- Fahy, J.V.; Dickey, B.F. Airway mucus function and dysfunction. N. Engl. J. Med. 2010, 363, 2233–2247. [Google Scholar] [CrossRef] [PubMed]
- Munkholm, M.; Mortensen, J. Mucociliary clearance: Pathophysiological aspects. Clin. Physiol. Funct. Imaging 2014, 34, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Wardlaw, A.J.; Brightling, C.E.; Green, R.; Woltmann, G.; Bradding, P.; Pavord, I.D. New insights into the relationship between airway inflammation and asthma. Clin. Sci. 2002, 103, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Brightling, C.E.; Symon, F.A.; Birring, S.S.; Bradding, P.; Pavord, I.D.; Wardlaw, A.J. TH2 cytokine expression in bronchoalveolar lavage fluid T lymphocytes and bronchial submucosa is a feature of asthma and eosinophilic bronchitis. J. Allergy Clin. Immunol. 2002, 110, 899–905. [Google Scholar] [CrossRef] [PubMed]
- James, A.L.; Wenzel, S. Clinical relevance of airway remodelling in airway diseases. Eur. Respir. J. 2007, 30, 134–155. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, C.; Tulic, M.K.; Hamid, Q. Airway remodelling in asthma: From benchside to clinical practice. Can. Respir J. 2010, 17, e85–e93. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.J.; Sykes, A.; Mallia, P.; Johnston, S.L. Asthma exacerbations: Origin, effect, and prevention. J. Allergy Clin. Immunol. 2011, 128, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Guarnieri, M.; Balmes, J.R. Outdoor air pollution and asthma. Lancet 2014, 383, 1581–1592. [Google Scholar] [CrossRef]
- Weinmayr, G.; Romeo, E.; De Sario, M.; Weiland, S.K.; Forastiere, F. Short-term effects of PM10 and NO2 on respiratory health among children with asthma or asthma-like symptoms: A systematic review and meta-analysis. Environ. Health Perspect. 2010, 118, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Spira-Cohen, A.; Chen, L.C.; Kendall, M.; Lall, R.; Thurston, G.D. Personal exposures to traffic-related air pollution and acute respiratory health among Bronx schoolchildren with asthma. Environ. Health Perspect. 2011, 119, 559–565. [Google Scholar] [CrossRef] [PubMed]
- McCreanor, J.; Cullinan, P.; Nieuwenhuijsen, M.J.; Stewart-Evans, J.; Malliarou, E.; Jarup, L.; Harrington, R.; Svartengren, M.; Han, I.; Ohman-Strickland, P.; et al. Respiratory effects of exposure to diesel traffic in persons with asthma. N. Engl. J. Med. 2007, 357, 2348–2358. [Google Scholar] [CrossRef] [PubMed]
- von Klot, S.; Wolke, G.; Tuch, T.; Heinrich, J.; Dockery, D.W.; Schwartz, J.; Kreyling, W.G.; Wichmann, H.E.; Peters, A. Increased asthma medication use in association with ambient fine and ultrafine particles. Eur. Respir. J. 2002, 20, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Nordenhall, C.; Pourazar, J.; Ledin, M.C.; Levin, J.O.; Sandstrom, T.; Adelroth, E. Diesel exhaust enhances airway responsiveness in asthmatic subjects. Eur. Respir. J. 2001, 17, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Szewczynska, M.; Posniak, M.; Dobrzynska, E. Study on individual PAHs content in ultrafine particles from solid fractions of diesel and biodiesel exhaust fumes. J. Chem. 2013, 2013, 528471. [Google Scholar] [CrossRef]
- Mabilia, R.; Cecinato, A.; Tomasi Sciano, M.C.; Di Palo, V.; Possanzini, M. Characterization of polycyclic aromatic hydrocarbons and carbonyl compounds in diesel exhaust emissions. Ann. Chim. 2004, 94, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Tobiszewski, M.; Namiesnik, J. PAH diagnostic ratios for the identification of pollution emission sources. Environ. Pollut. 2012, 162, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Ravindra, K.; Sokhi, R.; Van Grieken, R. Atmospheric polycyclic aromatic hydrocarbons: Source attribution, emission factors and regulation. Atmos. Environ. 2008, 42, 2895–2921. [Google Scholar] [CrossRef]
- Moorthy, B.; Chu, C.; Carlin, D.J. Polycyclic aromatic hydrocarbons: From metabolism to lung cancer. Toxicol. Sci. 2015, 145, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Lindner, K.; Strobele, M.; Schlick, S.; Webering, S.; Jenckel, A.; Kopf, J.; Danov, O.; Sewald, K.; Buj, C.; Creutzenberg, O.; et al. Biological effects of carbon black nanoparticles are changed by surface coating with polycyclic aromatic hydrocarbons. Part. Fibre Toxicol. 2017, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.Y.; Yim, D.H.; Moon, S.I.; Youn, J.W.; Kwon, H.J.; Oh, H.C.; Yang, J.J.; Park, S.K.; Yoo, K.Y.; Kim, H.S.; et al. Polycyclic aromatic hydrocarbon-induced oxidative stress, antioxidant capacity, and the risk of lung cancer: A pilot nested case-control study. Anticancer Res. 2013, 33, 3089–3097. [Google Scholar] [PubMed]
- Osgood, R.S.; Upham, B.L.; Hill, T., 3rd; Helms, K.L.; Velmurugan, K.; Babica, P.; Bauer, A.K. Polycyclic aromatic hydrocarbon-induced signaling events relevant to inflammation and tumorigenesis in lung cells are dependent on molecular structure. PLoS ONE 2014, 8, e65150. [Google Scholar] [CrossRef] [PubMed]
- Alessandrini, F.; Beck-Speier, I.; Krappmann, D.; Weichenmeier, I.; Takenaka, S.; Karg, E.; Kloo, B.; Schulz, H.; Jakob, T.; Mempel, M.; et al. Role of oxidative stress in ultrafine particle-induced exacerbation of allergic lung inflammation. Am. J. Respir. Crit. Care Med. 2009, 179, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Alessandrini, F.; Schulz, H.; Takenaka, S.; Lentner, B.; Karg, E.; Behrendt, H.; Jakob, T. Effects of ultrafine carbon particle inhalation on allergic inflammation of the lung. J. Allergy Clin. Immunol. 2006, 117, 824–830. [Google Scholar] [CrossRef] [PubMed]
- de Haar, C.; Hassing, I.; Bol, M.; Bleumink, R.; Pieters, R. Ultrafine carbon black particles cause early airway inflammation and have adjuvant activity in a mouse allergic airway disease model. Toxicol. Sci. 2005, 87, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Takano, H.; Yanagisawa, R.; Sakurai, M.; Ichinose, T.; Sadakane, K.; Yoshikawa, T. Effects of nano particles on antigen-related airway inflammation in mice. Respir. Res. 2005, 6, 106. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Takano, H.; Yanagisawa, R.; Sakurai, M.; Abe, S.; Yoshino, S.; Yamaki, K.; Yoshikawa, T. Effects of nanoparticles on lung physiology in the presence or absence of antigen. Int. J. Immunopathol. Pharmacol. 2007, 20, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Morawska, L.; Birmili, W.; Paasonen, P.; Hu, M.; Kulmala, M.; Harrison, R.M.; Norford, L.; Britter, R. Ultrafine particles in cities. Environ. Int. 2014, 66, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Aranda, A.; Diaz-de-Mera, Y.; Notario, A.; Rodriguez, D.; Rodriguez, A. Fine and ultrafine particles in small cities. A case study in the south of Europe. Environ. Sci. Pollut. Res. Int. 2015, 22, 18477–18486. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.H.; Lai, W.F.; Chiang, H.L. Characteristics of particulate constituents and gas precursors during the episode and non-episode periods. J. Air Waste Manag. Assoc. 2013, 63, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Kyjovska, Z.O.; Jacobsen, N.R.; Saber, A.T.; Bengtson, S.; Jackson, P.; Wallin, H.; Vogel, U. DNA damage following pulmonary exposure by instillation to low doses of carbon black (Printex 90) nanoparticles in mice. Environ. Mol. Mutagen. 2015, 56, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Lunding, L.P.; Webering, S.; Vock, C.; Behrends, J.; Wagner, C.; Holscher, C.; Fehrenbach, H.; Wegmann, M. Poly(inosinic-cytidylic) acid-triggered exacerbation of experimental asthma depends on IL-17A produced by NK cells. J. Immunol. 2015, 194, 5615–5625. [Google Scholar] [CrossRef] [PubMed]
- Bonser, L.R.; Erle, D.J. Airway Mucus and Asthma: The Role of MUC5AC and MUC5B. J. Clin. Med. 2017, 6, 112. [Google Scholar] [CrossRef] [PubMed]
- Bakand, S.; Hayes, A.; Dechsakulthorn, F. Nanoparticles: A review of particle toxicology following inhalation exposure. Inhal. Toxicol. 2012, 24, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Keohavong, P.; Melacrinos, A.; Shukla, R. In vitro mutational spectrum of cyclopenta[cd]pyrene in the human HPRT gene. Carcinogenesis 1995, 16, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Nesnow, S.; Mass, M.J.; Ross, J.A.; Galati, A.J.; Lambert, G.R.; Gennings, C.; Carter, W.H., Jr.; Stoner, G.D. Lung tumorigenic interactions in strain A/J mice of five environmental polycyclic aromatic hydrocarbons. Environ. Health Perspect. 1998, 106 (Suppl. 6), 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- The Danish Environmental Protection Agency. Exposure to Nanomaterials from the Danish Environment; The Danish Environmental Protection Agency: København, Denmark, 2015; ISBN 978-87-93283-54-1. [Google Scholar]
- Department for Environment, Food and Rural A. Air Pollution in the UK 2013. 2014. Available online: https://uk-air.defra.gov.uk/assets/documents/annualreport/air_pollution_uk_2013_issue_1.pdf (accessed on 1 November 2014).
- Netherlands Environmental Assessment Agency. PM2.5 in the Netherlands-Consequences of the New European Air Quality Standards; MNP Report 50009901/2007; Netherlands Environmental Assessment Agency: The Hague, The Netherlands, 2007; ISBN 978-90-6960-176-2. [Google Scholar]
- European Environment Agency. Air quality in Europe—2016 Report; EEA Report No 28/2016; European Environment Agency: Copenhagen, Denmark, 2016; ISBN 978-92-9213-847-9. [Google Scholar]
- Guerreiro, C.B.B.; Foltescu, V.; de Leeuw, F. Air quality status and trends in Europe. Atmos. Environ. 2014, 98, 376–384. [Google Scholar] [CrossRef]
- Lowsen, D.H.; Conway, G.A. Air Pollution in Major Chinese Cities: Some Progress, But Much More to Do. J. Environ. Prot. 2017, 7, 2081–2094. [Google Scholar] [CrossRef] [PubMed][Green Version]
- San Martini, F.M.; Hasenkopf, C.A.; Roberts, D.C. Statistical analysis of PM2.5 observations from diplomatic facilities in China. Atmos. Environ. 2015, 110, 174–185. [Google Scholar] [CrossRef]
- Zhao, P.; Lin, C.-C. Air Quality at Night Markets in Taiwan. J. Air Waste Manag. Assoc. 2010, 60, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Naeher, L.P. A review of traffic-related air pollution exposure assessment studies in the developing world. Environ. Int. 2006, 32, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Mouse Phenome Database at The Jackson Laboratory. 2014. Available online: https://phenome.jax.org/measureset/35101 (accessed on 1 October 2014).
- Oldham, M.J.; Phalen, R.F.; Budiman, T. Comparison of predicted and experimentally measured aerosol deposition efficiency in BALB/c mice in a new nose-only exposure system. Aerosol. Sci. Technol. 2009, 43, 970–977. [Google Scholar] [CrossRef]
- Möller, W.; Felten, K.; Sommerer, K.; Scheuch, G.; Meyer, G.; Meyer, P.; Haussinger, K.; Kreyling, W.G. Deposition, retention, and translocation of ultrafine particles from the central airways and lung periphery. Am. J. Respir. Crit. Care Med. 2008, 177, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Schaumann, F.; Fromke, C.; Dijkstra, D.; Alessandrini, F.; Windt, H.; Karg, E.; Muller, M.; Winkler, C.; Braun, A.; Koch, A.; et al. Effects of ultrafine particles on the allergic inflammation in the lung of asthmatics: Results of a double-blinded randomized cross-over clinical pilot study. Part. Fibre Toxicol. 2014, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Gour, N.; Wills-Karp, M. IL-4 and IL-13 signaling in allergic airway disease. Cytokine 2015, 75, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1beta secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Alam, R.; York, J.; Boyars, M.; Stafford, S.; Grant, J.A.; Lee, J.; Forsythe, P.; Sim, T.; Ida, N. Increased MCP-1, RANTES, and MIP-1alpha in bronchoalveolar lavage fluid of allergic asthmatic patients. Am. J. Respir. Crit. Care Med. 1996, 153, 1398–1404. [Google Scholar] [CrossRef] [PubMed]
- Sahiner, U.M.; Birben, E.; Erzurum, S.; Sackesen, C.; Kalayci, O. Oxidative stress in asthma. World Allergy Organ. J. 2011, 4, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Dunican, E.M.; Elicker, B.M.; Gierada, D.S.; Nagle, S.K.; Schiebler, M.L.; Newell, J.D.; Raymond, W.W.; Lachowicz-Scroggins, M.E.; Di Maio, S.; Hoffman, E.A.; et al. Mucus plugs in patients with asthma linked to eosinophilia and airflow obstruction. J. Clin. Investig. 2018, 128, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K. Promoting effects of nanoparticles/materials on sensitive lung inflammatory diseases. Environ. Health Prev. Med. 2011, 16, 139–143. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Roy, M.G.; Livraghi-Butrico, A.; Fletcher, A.A.; McElwee, M.M.; Evans, S.E.; Boerner, R.M.; Alexander, S.N.; Bellinghausen, L.K.; Song, A.S.; Petrova, Y.M.; et al. Muc5b is required for airway defence. Nature 2014, 505, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.M.; Raclawska, D.S.; Ttofali, F.; Liptzin, D.R.; Fletcher, A.A.; Harper, D.N.; McGing, M.A.; McElwee, M.M.; Williams, O.W.; Sanchez, E.; et al. The polymeric mucin Muc5ac is required for allergic airway hyperreactivity. Nat. Commun. 2015, 6, 6281. [Google Scholar] [CrossRef] [PubMed]
- Lachowicz-Scroggins, M.E.; Yuan, S.; Kerr, S.C.; Dunican, E.M.; Yu, M.; Carrington, S.D.; Fahy, J.V. Abnormalities in MUC5AC and MUC5B Protein in Airway Mucus in Asthma. Am. J. Respir. Crit. Care Med. 2016, 194, 1296–1299. [Google Scholar] [CrossRef] [PubMed]
- Bonser, L.R.; Zlock, L.; Finkbeiner, W.; Erle, D.J. Epithelial tethering of MUC5AC-rich mucus impairs mucociliary transport in asthma. J. Clin. Investig. 2016, 126, 2367–2371. [Google Scholar] [CrossRef] [PubMed]
- Erle, D.J.; Sheppard, D. The cell biology of asthma. J. Cell Biol. 2014, 205, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, P.G.; Modrek, B.; Choy, D.F.; Jia, G.; Abbas, A.R.; Ellwanger, A.; Koth, L.L.; Arron, J.R.; Fahy, J.V. T-helper type 2-driven inflammation defines major subphenotypes of asthma. Am. J. Respir. Crit. Care Med. 2009, 180, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Sigurdsson, H.H.; Kirch, J.; Lehr, C.M. Mucus as a barrier to lipophilic drugs. Int. J. Pharm. 2013, 453, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, M.; Fehrenbach, H.; Fehrenbach, A.; Held, T.; Schramm, C.; Garn, H.; Renz, H. Involvement of distal airways in a chronic model of experimental asthma. Clin. Exp. Allergy 2005, 35, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Baker, G.L.; Shultz, M.A.; Fanucchi, M.V.; Morin, D.M.; Buckpitt, A.R.; Plopper, C.G. Assessing gene expression in lung subcompartments utilizing in situ RNA preservation. Toxicol. Sci. 2004, 77, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Mattfeldt, T.; Mall, G.; Gharehbaghi, H.; Moller, P. Estimation of surface area and length with the orientator. J. Microsc. 1990, 159, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Weibel, E.R.; Hsia, C.C.W.; Ochs, M. How much is there really? Why stereology is essential in lung morphometry. J. Appl. Physiol. 2007, 102, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Crouzier, T.; Beckwitt, C.H.; Ribbeck, K. Mucin Multilayers Assembled through Sugar-Lectin Interactions. Biomacromolecules 2012, 13, 3401–3408. [Google Scholar] [CrossRef] [PubMed]
- Kawano, K.; Uehara, F.; Sameshima, M.; Ohba, N. Application of lectins for detection of goblet cell carbohydrates of the human conjunctiva. Exp. Eye Res. 1984, 38, 439–447. [Google Scholar] [CrossRef]
- Chazotte, B. Labeling membrane glycoproteins or glycolipids with fluorescent wheat germ agglutinin. Cold Spring Harb. Protoc. 2011, 2011, pdb.prot5623. [Google Scholar] [CrossRef] [PubMed]
- Graziadei, G.A.; Graziadei, P.P. Neurogenesis and neuron regeneration in the olfactory system of mammals. II. Degeneration and reconstitution of the olfactory sensory neurons after axotomy. J. Neurocytol. 1979, 8, 197–213. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Particle Characteristics | Printex®90 (P90) | P90 Coated with benzo[a]pyrene (P90-BaP) | P90 Coated with 9-Nitroanthracene (P90-9NA) | Acetylene Soot with PAH-Mixture (AS-PAH) |
|---|---|---|---|---|
| Mean primary particle size (nm) | 16.5 ± 0.4 | 16.5 ± 0.4 * | 16.5 ± 0.4 * | 14.4 ± 0.3 * |
| Specific surface area (m2/g) | 302 ± 16 | 120 ± 2 | 91 # | 124 ± 2 |
| Mass loss up to 1000 °C (%) | 0.5 ± 0.4 | 10.0 ± 0.2 | 14.6 ± 0.1 | 15.3 ± 0.2 |
| Hydrodynamic diameter in water/BSA (nm) | 166 ± 9 | 163 ± 6 | 161 ± 3 | 163 ± 6 |
| ζ-potential in water/BSA (mV) | −33 ± 1 | −32 ± 1 | −33 ± 1 | −32 ± 1 |
| Protein | PBS/H2O | OVA/H2O | OVA/P90 | OVA/P90-BaP | OVA/P90-9NA | OVA/AS-PAH |
|---|---|---|---|---|---|---|
| Mcp-1 | 0.01 ± 0.01 | 27.0 ± 5.7 | 12.7 ± 3.9 * | 7.4 ± 5.5 * | 11.7 ± 5.7 * | 0.6 ± 0.3 * |
| KC | 9.5 ± 2.3 | 21.6 ± 5.7 | 16.7 ± 4.3 | 17.7 ± 4.2 | 21.3 ± 7.9 | 9.9 ± 2.5 |
| IL-1β | 0.06 ± 0.02 | 1.3 ± 0.2 | 0.8 ± 0.2 | 0.8 ± 0.2 | 0.7 ± 0.2 * | 0.5 ± 0.1 * |
| IL-6 | 0.4 ± 0.1 | 1.1 ± 0.3 | 0.8 ± 0.2 | 0.7 ± 0.1 | 0.7 ± 0.2 | 0.7 ± 0.2 |
| IL-4 | 0.8 ± 0.6 | 1.2 ± 0.2 | 0.7 ± 0.1 | 0.7 ± 0.1 * | 0.9 ± 0.2 | 0.6 ± 0.2 * |
| IL-5 | 0.8 ± 0.3 | 12.7 ± 2.1 | 10.4 ± 1.8 | 13.4 ± 1.8 | 11.1 ± 1.8 | 11.7 ± 2.2 |
| IL-13 | 0.03 ± 0.01 | 7.3 ± 1.1 | 4.4 ± 0.9 * | 5.7 ± 1.1 | 4.4 ± 0.6 * | 5.4 ± 1.2 |
| IL-17A | 0.2 ± 0.1 | 0.4 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.4 ± 0.1 | 0.3 ± 0.1 |
| Gene | Airway Section | PBS/H2O | OVA/H2O | OVA/P90 | OVA/P90-BaP | OVA/P90-9NA | OVA/AS-PAH |
|---|---|---|---|---|---|---|---|
| Cyp1a1 | intrapulmonary airways | 1.0 ± 0.1 | 1.0 ± 0.2 | 1.3 ± 0.4 | 1.3 ± 0.2 | 0.7 ± 0.1 | 1.5 ± 0.3 |
| tracheal epithelial cells | bld | bld | bld | bld | bld | bld | |
| Cyp1b1 | intrapulmonary airways | 1.0 ± 0.1 | 0.6 ± 0.1 | 0.6 ± 0.1 | 0.8 ± 0.1 | 0.5 ± 0.1 | 0.9 ± 0.1 |
| tracheal epithelial cells | 1.1 ± 0.2 | 1.2 ± 0.3 | 1.4 ± 0.3 | 1.3 ± 0.4 | 1.6 ± 0.7 | 2.8 ± 0.9 | |
| Gpx3 | intrapulmonary airways | 1.0 ± 0.1 | 0.9 ± 0.1 | 0.8 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 |
| tracheal epithelial cells | 1.0 ± 0.1 | 0.3 ± 0.1 | 0.6 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.4 ± 0.1 | |
| Gr | intrapulmonary airways | 1.0 ± 0.1 | 1.3 ± 0.1 | 1.1 ± 0.1 | 1.5 ± 0.1 | 1.0 ± 0.1 | 1.5 ± 0.1 |
| tracheal epithelial cells | 1.0 ± 0.2 | 1.1 ± 0.4 | 1.5 ± 0.3 | 0.9 ± 0.3 | 0.8 ± 0.2 | 0.9 ± 0.4 | |
| HO-1 | intrapulmonary airways | 1.0 ± 0.1 | 1.3 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.2 ± 0.1 |
| tracheal epithelial cells | 1.2 ± 0.3 | 1.0 ± 0.2 | 1.1 ± 0.2 | 1.1 ± 0.2 | 1.1 ± 0.4 | 1.1 ± 0.3 | |
| Mcp-1 | intrapulmonary airways | 1.0 ± 0.6 | 12 ± 5 | 11 ± 4 | 10 ± 4 | 8 ± 3 | 11 ± 4 |
| tracheal epithelial cells | 1.1 ± 0.3 | 1.2 ± 0.3 | 1.8 ± 0.6 | 0.4 ± 0.1 | 0.5 ± 0.2 | 1.1 ± 0.5 | |
| KC | intrapulmonary airways | 1.0 ± 0.3 | 2.1 ± 0.6 | 1.7 ± 0.3 | 1.9 ± 0.3 | 1.1 ± 0.2 | 1.5 ± 0.3 |
| tracheal epithelial cells | 1.7 ± 0.2 | bld | bld | bld | bld | bld | |
| IL-6 | intrapulmonary airways | 1.0 ± 1.3 | 7.9 ± 1.2 | 6.8 ± 1.4 | 8.5 ± 1.0 | 4.3 ± 0.5 * | 5.7 ± 0.8 |
| tracheal epithelial cells | 1.2 ± 0.3 | 1.5 ± 0.6 | 2.4 ± 1.3 | 3.1 ± 1.7 | 0.6 ± 0.3 | 3.5 ± 1.8 | |
| IL-13 | intrapulmonary airways | 1.0 ± 0.4 | 52 ± 8 | 40 ± 7 | 48 ± 7 | 34 ± 4 * | 48 ± 7 |
| tracheal epithelial cells | 1.1 ± 0.3 | 41 ± 14 | 27 ± 7 | 20 ± 9 | 26 ± 10 | 31 ± 10 | |
| IL-17a | intrapulmonary airways | 1.0 ± 2.9 | 13 ± 3 | 13 ± 3 | 11 ± 1 | 8 ± 2 | 19 ± 4 |
| tracheal epithelial cells | bld | bld | bld | bld | bld | bld | |
| Muc5ac | intrapulmonary airways | 1.0 ± 0.3 | 44 ± 11 | 29 ± 4 | 52 ± 9 | 26 ± 5 | 82 ± 25 |
| tracheal epithelial cells | 1.1 ± 0.3 | 35 ± 10 | 101 ± 15 ** | 38 ± 11 | 39 ± 23 | 39 ± 20 | |
| Muc5b | intrapulmonary airways | 1.0 ± 0.2 | 31 ± 5 | 18 ± 3 * | 32 ± 4 | 20 ± 2 | 39 ± 11 |
| tracheal epithelial cells | 1.1 ± 0.2 | 2.2 ± 0.6 | 2.2 ± 0.2 | 2.1 ± 0.2 | 2.2 ± 0.4 | 2.4 ± 0.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindner, K.; Webering, S.; Stroebele, M.; Bockhorn, H.; Hansen, T.; König, P.; Fehrenbach, H. Low Dose Carbon Black Nanoparticle Exposure Does Not Aggravate Allergic Airway Inflammation in Mice Irrespective of the Presence of Surface Polycyclic Aromatic Hydrocarbons. Nanomaterials 2018, 8, 213. https://doi.org/10.3390/nano8040213
Lindner K, Webering S, Stroebele M, Bockhorn H, Hansen T, König P, Fehrenbach H. Low Dose Carbon Black Nanoparticle Exposure Does Not Aggravate Allergic Airway Inflammation in Mice Irrespective of the Presence of Surface Polycyclic Aromatic Hydrocarbons. Nanomaterials. 2018; 8(4):213. https://doi.org/10.3390/nano8040213
Chicago/Turabian StyleLindner, Karina, Sina Webering, Michael Stroebele, Henning Bockhorn, Tanja Hansen, Peter König, and Heinz Fehrenbach. 2018. "Low Dose Carbon Black Nanoparticle Exposure Does Not Aggravate Allergic Airway Inflammation in Mice Irrespective of the Presence of Surface Polycyclic Aromatic Hydrocarbons" Nanomaterials 8, no. 4: 213. https://doi.org/10.3390/nano8040213
APA StyleLindner, K., Webering, S., Stroebele, M., Bockhorn, H., Hansen, T., König, P., & Fehrenbach, H. (2018). Low Dose Carbon Black Nanoparticle Exposure Does Not Aggravate Allergic Airway Inflammation in Mice Irrespective of the Presence of Surface Polycyclic Aromatic Hydrocarbons. Nanomaterials, 8(4), 213. https://doi.org/10.3390/nano8040213