
Cyto–Genotoxic Effect Causing Potential of Polystyrene Micro-Plastics in Terrestrial Plants


Abstract
:1. Introduction
2. Materials and Methods
2.1. Model Plant
2.2. Ps-MPs Working Concentrations
2.3. Experimentation
2.4. Microscopic Analysis
2.5. Calculations
2.6. Statistical Analysis
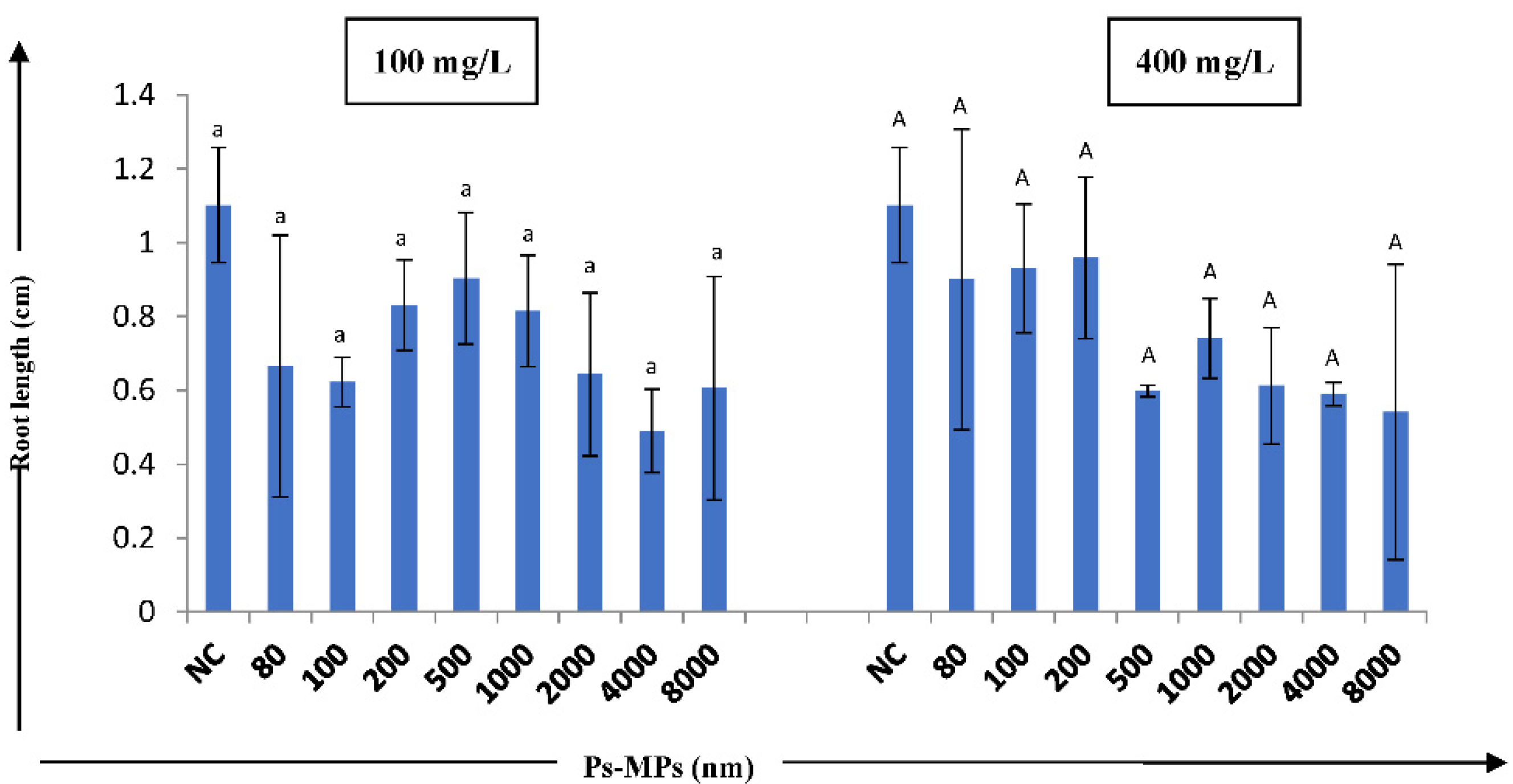
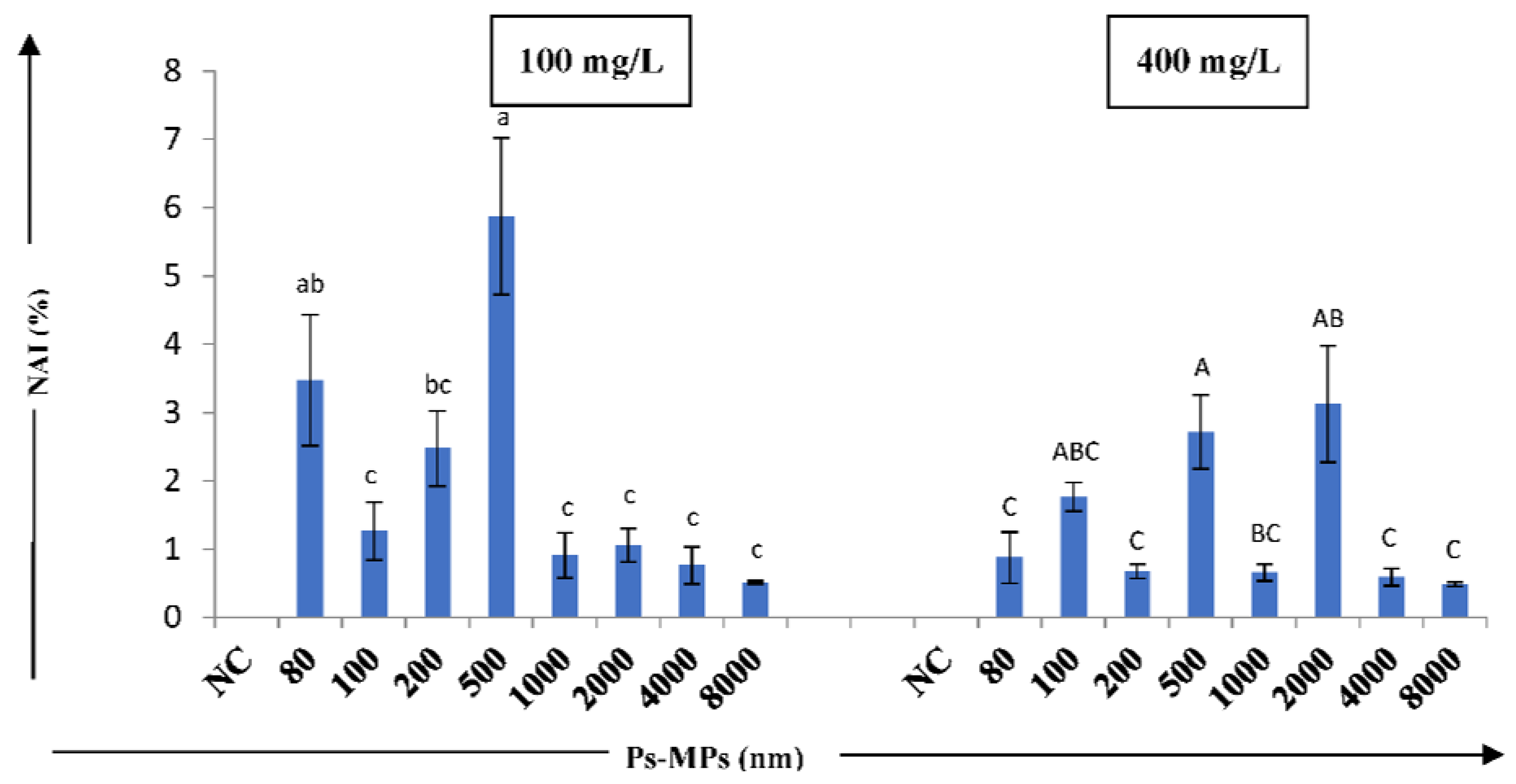
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gopinath, P.M.; Saranya, V.; Vijayakumar, S.; Meera, M.M.; Ruprekha, S.; Kunal, R.; Pranay, A.; Thomas, J.; Mukherjee, A.; Chandrasekaran, N. Assessment on interactive prospectives of nano-plastics with plasma proteins and the toxicological impacts of virgin, coronated and environmentally released-nanoplastics. Sci. Rep. 2019, 9, 8860. [Google Scholar] [CrossRef] [PubMed]
- Chamas, A.; Moon, H.; Zheng, J.; Qiu, Y.; Tabassum, T.; Jang, J.H.; Abu-Omar, M.; Scott, S.L.; Suh, S. Degradation rates of plastics in the environment. ACS Sustain. Chem. Eng. 2020, 8, 3494–3511. [Google Scholar] [CrossRef] [Green Version]
- World Economic Forum. The New Plastics Economy: Rethinking the Future of Plastics; Ellen MacArthur Foundation and McKinsey & Company: Geneva, Switzerland, 2016. [Google Scholar]
- Consumer Reports. You Are Literally Eating Micro-Plastics. How You Can Cut Down Exposure to Them. The Washington Post. 2019. Available online: https://www.google.com/amp/s/www.washingtonpost.com/health/youre-literally-eating microplastics-how-you-can-cut-down-exposure-to-them/2019/10/04/ 22ebd fb6 -e17a 11e9-8dc8-498eabc129a0_story.html%3foutputType=amp. (accessed on 20 September 2021).
- Rhodes, C.J. Plastic pollution and potential solutions. Sci. Prog. 2018, 101, 207–260. [Google Scholar] [CrossRef] [PubMed]
- Verla, A.W.; Enyoh, C.E.; Verla, E.N. Microplastics, an emerging concern: A review of analytical techniques for detecting and quantifying microplatic. Anal. Methods Environ. Chem. J. 2019, 2, 15–32. [Google Scholar] [CrossRef]
- Verla, A.W.; Enyoh, C.E.; Verla, E.N.; Nwarnorh, K.O. Micro-plastic toxic chemical interaction: A review study on quantified levels, mechanism and implications. SNAppl. Sci. 2019, 1, 400. [Google Scholar] [CrossRef] [Green Version]
- Bouwmeester, H.; Hollman, P.C.; Peters, R.J. Potential health impact of environmentally released micro- and nanoplastics in the human food production chain. Environ. Sci. Technol. 2015, 49, 8932–8947. [Google Scholar] [CrossRef]
- Plastic Europe. Plastics–the Facts 2018. An Analysis of European Plastics Production, Demand and Waste Data; PlasticsEurope AISBL: Brussels, Belgium, 2018. [Google Scholar]
- Zhu, B.K.; Fang, Y.M.; Zhu, D.; Christie, P.; Ke, X.; Zhu, Y.G. Exposure of nanoplastics disturbs the gut microbiome in the soil oligochaete Enchytraeus crypticus. Environ. Pollut. 2018, 239, 408–415. [Google Scholar] [CrossRef]
- Loos, C.; Syrovets, T.; Musyanovych, A.; Mailänder, V.; Landfester, K.; Nienhaus, G.U.; Simmet, T. Functionalized polystyrene nanoparticles as a platform for studying bio-nano interactions. Beilstein. J. Nano Technol. 2014, 5, 2403–2412. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Seijo, A.; Lourenço, J.; Rocha-Santos, T.; Da Costa, J.; Duarte, A.; Vala, H.; Pereira, R. Histo-pathological and molecular effects of micro-plastics in Eisenia Andrei Bouché. Environ Pollut. 2017, 220, 495–503. [Google Scholar] [CrossRef]
- Zhang, G.S.; Liu, Y.F. The distribution of microplastics in soil aggregate fractions in southwestern China. Sci. Total Environ. 2018, 642, 12–20. [Google Scholar] [CrossRef]
- Huang, F.Y.; Yang, K.; Zhang, Z.X.; Su, J.Q.; Zhu, Y.G.; Zhang, X. Effects of microplastics on antibiotic resistance genes in estuarine sediments. Acta Sci. Circum. 2019, 8, 2234–2239. [Google Scholar] [CrossRef]
- Forte, M.; Iachetta, G.; Tussellino, M.; Carotenuto, R.; Prisco, M.; Falco, M.D.; Laforgia, V.; Valiante, S. Polystyrene nanoparticles internalization in human gastric adenocarcinoma cells. Toxicol. Vitr. 2016, 31, 126–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, H.K.A.; Chan, K.Y.K. Negative effects of microplastic exposure on growth and development of Crepidula onyx. Environ Pollut. 2018, 233, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.; Lindeque, P.; Fileman, E.; Halsband, C.; Galloway, T.S. The impact of polystyrene microparticles on feeding, function and fecundity in the marine copepod Calanus helgolandicus. Environ. Sci. Technol. 2015, 49, 1130–1137. [Google Scholar] [CrossRef]
- Botterell, Z.L.R.; Beaumont, N.; Dorrington, T.; Steinke, M.; Thompson, R.C.; Lindeque, P.K. Bioavailability and effects of microplastics on marine zooplankton: A review. Environ. Pollut. 2019, 245, 98–110. [Google Scholar] [CrossRef]
- Yong, M.M.H.; Leistenschneider, C.; Miranda, J.A.; Paler, M.K.; Legaspi, C.; Germanov, E.; Araujo, G.; Burkhardt-Holm, P.; Erni-Cassola, G. Microplastics in fecal samples of whale sharks (Rhincodon typus) and from surface water in the Philippines. Micro. Nano. 2021, 1, 17. [Google Scholar] [CrossRef]
- Ziajahromi, S.; Kumar, A.; Neale, P.A.; Leusch, F.D.L. Impact of microplastic beads and fibers on water flea (Ceriodaphnia dubia) survival, growth, and reproduction: Implications of single and mixture exposures. Environ. Sci. Technol. 2017, 51, 13397–13406. [Google Scholar] [CrossRef] [Green Version]
- Horn, D.A.; Granek, E.F.; Steele, C.L. Effects of environmentally relevant concentrations of microplastic fibers on Pacific mole crab (Emerita analoga) mortality and reproduction. Limnol. Ocean. Lett. 2019, 5, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Park, E.-J.; Han, S.; Park, E.-J.; Seong, E.; Lee, G.-H.; Kim, D.-W.; Son, H.-Y.; Han, H.-Y.; Lee, B.-S. Repeated-oral dose toxicity of polyethylene microplastics and the possible implications on reproduction and development of the next generation. Toxicol. Lett. 2020, 324, 75–85. [Google Scholar] [CrossRef]
- Jin, H.; Ma, T.; Sha, X.; Liu, Z.; Zhou, Y.; Meng, X.; Chen, Y.; Han, X.; Ding, J. Polystyrene microplastics induced male reproductive toxicity in mice. J. Hazard Mater. 2021, 401, 123430. [Google Scholar] [CrossRef]
- Ragusa, A.; Svelato, A.; Santacroce, C.; Catalano, P.; Notarstefano, V.; Carnevali, O.; Papa, F.; Rongioletti, M.C.A.; Baiocco, F.; Draghi, S.; et al. Plasticenta: First evidence of microplastics in human placenta. Environ. Int. 2021, 146, 106274. [Google Scholar] [CrossRef] [PubMed]
- Cobanoglu, H.; Belivermis, M.; Sikdokur, E.; Kilic, O.; Cayir, A. Genotoxic and cytotoxic effects of polyethylene microplastics on human peripheral blood lymphocytes. Chemosphere 2021, 272, 129805. [Google Scholar] [CrossRef] [PubMed]
- Pathan, S.I.; Arfaioli, P.; Bardelli, T.; Ceccherini, M.T.; Nannipieri, P.; Pietramellara, G. Soil pollution from micro- and nanoplastic debris: A hidden and unknown biohazard. Sustainability 2020, 12, 7255. [Google Scholar] [CrossRef]
- Horton, A.A.; Svendsen, C.; Williams, R.J.; Spurgeon, D.J.; Lahive, E. Large microplastic particles in sediments of tributaries of the River Thames, UK-Abundance, sources and methods for effective quantification. Mar. Pollut. Bull. 2017, 114, 218–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wu, C.; Xue, Q.; Hui, X. Effects of plastic contamination on water evaporation and desiccation cracking in soil. Sci. Total Environ. 2019, 654, 576–582. [Google Scholar] [CrossRef]
- Li, Q.; Feng, Z.; Zhang, T.; Ma, C.; Shi, H. Microplastics in the commercial seaweed nori. J. Hazard Mater. 2020, 388, 122060. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, Z.; Wollmann, C.; Schaefer, M.; Buchmann, C.; David, J.; Tröger, J.; Muñoz, K.; Frör, O.; Schaumann, G.E. Plastic mulching in agriculture. Trading short-term agronomic benefits for long-term soil degradation. Sci. Total Environ. 2016, 550, 690–705. [Google Scholar] [CrossRef]
- Bläsing, M.; Amelung, W. Plastics in soil: Analytical methods and possible sources. Sci. Total Environ. 2018, 612, 422–435. [Google Scholar] [CrossRef]
- Brahney, J.; Mahowald, N.; Prank, M.; Cornwell, G.; Klimont, Z.; Matsul, H.; Prather, K.A. Constraining the atmospheric limb of the plastic cycle. Proc. Nat. Acad. Sci. USA 2021, 118, e2020719118. [Google Scholar] [CrossRef]
- Yu, M.; Van Der Ploeg, M.; Lwanga, E.H.; Yang, X.; Zhang, S.; Ma, X.; Ritsema, C.J.; Geissen, V. Leaching of microplastics by preferential flow in earthworm (Lumbricus terrestris) burrows. Environ. Chem. 2019, 16, 31–41. [Google Scholar] [CrossRef]
- Bandmann, V.; Müller, J.D.; Köhler, T.; Homann, U. Uptake of fluorescent nanobeads into BY2-cells involves clathrin dependent and clathrin-independent endocytosis. FEBS Lett. 2012, 586, 3626–3632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Changyin, Z.; Chao, W.; Gu, C. Occurrence and ecological impacts of microplastics in soil systems: A Review. Bull.Environ. Contam. Toxicol. 2019, 102, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Campanale, C.; Massarelli, C.; Savino, I.; Locaputo, V.; Uricchio, V.F. A detailed review study on potential effects of microplastics and additives of concern on human health. Int. J. Environ. Res. Public Health 2020, 17, 1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barboza, L.G.A.; Vieira, L.R.; Branco, V.; Figueiredo, N.; Carvalho, F.; Carvalho, C.; Guilhermino, L. Microplastics cause neurotoxicity, oxidative damage and energy-related changes and interact with the bioaccumulation of mercury in the European seabass, Dicentrarchuslabrax (Linnaeus, 1758). Aquat. Toxicol. 2018, 195, 49–57. [Google Scholar] [CrossRef]
- Banaee, M.; Soltanian, S.; Sureda, A.; Gholamhosseini, A.; Haghi, B.N.; Akhlaghi, M.; Derikvandy, A. Evaluation of single and combined effects of cadmium and micro-plastic particles on biochemical and immunological parameters of common carp (Cyprinus carpio). Chemosphere 2019, 236, 124335. [Google Scholar] [CrossRef]
- Bradney, L.; Wijesekara, H.; Palansooriya, K.N.; Obadamudalige, N.; Bolan, N.S.; Ok, Y.S.; Rinklebe, J.; Kim, K.H.; Kirkham, M.B. Particulate plastics as a vector for toxic trace-element uptake by aquatic and terrestrial organisms and human health risk. Environ. Int. 2019, 131, 104937. [Google Scholar] [CrossRef]
- Singh, R.; Gautam, N.; Mishra, A.; Gupta, R. Heavy metals and living systems: An overview. Indian J. Pharmacol. 2011, 43, 246. [Google Scholar] [CrossRef] [Green Version]
- Boyle, K.; Ormeci, B. Microplastics and nanoplastics in the freshwater and terrestrial environment: A review. Water 2020, 12, 2633. [Google Scholar] [CrossRef]
- Rillig, M.C.; Lehmann, A.; Machado de Souza, A.A.; Yang, G. Microplastic effects on plants. New Phyto. 2019, 223, 1066–1070. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Chen, H.; Liao, Y.; Ye, Z.; Li, M.; Klobucar, G. Ecotoxicity and genotoxicity of polystyrene microplastics on higher plant Vicia faba. Environ. Pollut. 2019, 250, 831–838. [Google Scholar] [CrossRef]
- Taylor, S.; Pearce, C.; Sanguinet, K.; Hu, D.; Chrisler, W.; Kim, Y.; Wang, Z.; Flury, M. Polystyrene nano- and microplastic accumulation at Arabidopsis and wheat root cap cells, but no evidence for uptake into roots. Environ. Sci. Nano. 2020, 7, 1942–1953. [Google Scholar] [CrossRef]
- Qi, Y.; Yang, X.; Pelaez, A.M.; Lwanga, H.E.; Beriot, N.; Gertsen, H.; Garbeva, P.; Geissen, V. Macro- and micro- plastics in soil-plant system: Effects of plastic mulch film residues on wheat (Triticum aestivum) growth. Sci. Total Environ. 2018, 645, 1048–1056. [Google Scholar] [CrossRef] [PubMed]
- Verla, A.W.; Enyoh, C.E.; Obinna, I.B.; Verla, E.N.; Qing, W.; Chowdhury Md, A.H.; Enyoh, E.C.; Chowdhury, T. Effect of macro-and micro-plastics in soil on growth of Juvenile Lime Tree (Citrus aurantium). AIMS Environ. Sci. 2020, 7, 526–541. [Google Scholar] [CrossRef]
- Smith, M.; Love, D.C.; Rochman, C.M.; Neff, R.A. Microplastics in seafood and the implications for human health. Curr. Environ. Health Rep. 2018, 5, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.H.; Monteiro, R.T.R. Toxicity assessment of silica nanoparticles on Allium cepa. Ecotoxicol. Environ. Contam. 2017, 12, 25–31. [Google Scholar] [CrossRef]
- Kaur, M.; Bhatti, S.S.; Soodan, R.K.; Katnoria, J.K.; Bhardwaj, R.; Nagpal, A.K.; Xu, M. Physico-chemical characterization of agricultural soil samples and their modulatory effects on cytogenetic and biochemical parameters of Allium cepa. J. Soil Sci. Plant Nutr. 2021, 21, 1890–1903. [Google Scholar] [CrossRef]
- Datta, S.; Singh, J.; Singh, J.; Singh, S.; Singh, S. Assessment of genotoxic effects of pesticide and vermicompost treated soil with Allium cepa test. Sustain. Environ. Res. 2018, 28, 171–178. [Google Scholar] [CrossRef]
- Verma, S.; Srivastava, A. Morphotoxicity and cytogenotoxicity of pendimethalin in the test plant Allium cepa L.—A biomarker based study. Chemosphere 2018, 206, 248–254. [Google Scholar] [CrossRef]
- Maity, S.; Chatterjee, A.; Guchhait, R.; Deb, S.; Pramanick, K. Cytogenotoxic potential of a hazardous material, polystyrene microparticles on Allium cepa L. J. Hazard Mater. 2020, 385, 121560. [Google Scholar] [CrossRef]
- Giorgetti, L.; Span_o, C.; Muccifora, S.; Bottega, S.; Barbieri, F.; Bellani, L.; Ruffini Castiglione, M. Exploring the interaction between polystyrene nanoplastics and Allium cepa during germination: Internalization in root cells, induction of toxicity and oxidative stress. Plant Physiol. Biochem. 2020, 149, 170–177. [Google Scholar] [CrossRef]
- Levan, A. The effect of colchicine on root mitosis in Allium. Hereditas 1938, 24, 471–486. [Google Scholar] [CrossRef]
- Leme, D.M.; Marin-Morales, M.A. Allium cepa test in environmental monitoring: A review on its application. Mutat. Res. Rev. Mutat. Res. 2009, 682, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Sabeen, M.; Mahmood, Q.; Bhatti, Z.A.; Faridullah; Irshad, M.; Bilal, M.; Hayat, T.; Irshad, U.; Akbar, T.A.; Arslan, M.; et al. Allium cepa assay based comparative study of selected vegetables and the chromosomal aberrations due to heavy metal accumulation. Saudi J. Biol. Sci. 2020, 27, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.; Sharma, A. Chromosome Techniques: Theory and Practice, 3rd ed.; Butterworths and Co. Ltd.: London, UK, 1980. [Google Scholar]
- Hemachandra, C.K.; Pathiratne, A. Assessing toxicity of copper, cadmium and chromium levels relevant to discharge limits of industrial effluents into inland surface waters using common onion, Allium cepa bioassay. Bull. Environ. Contam. Toxicol. 2015, 94, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Hemachandra, C.K.; Pathiratne, A. Cytogenotoxicity screening of source water, wastewater and treated water of drinking water treatment plants using two in vivo test systems: Allium cepa root based and Nile tilapia erythrocyte based tests. Water Res. 2017, 108, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Atuanya, E.I.; Udochukwu, U.; Dave-Omoregie, A.O. Bioavailability and Toxicity of Plastic Contaminants to Soil and Soil Bacteria. Br. Microbiol. Res. J. 2016, 13, 1–8. [Google Scholar] [CrossRef]
- Scherer, C.; Brennholt, N.; Reifferscheid, G.; Wagner, M. Feeding type and development drive the ingestion of microplastics by freshwater invertebrates. Sci. Rep. 2017, 7, 1–9. [Google Scholar]
- D’Aquino, L.; Depinto, M.C.; Nardi, L.; Morgana, M.; Tommasi, F. Effect of some light rare earth elements on seed germination, seedling growth and antioxidant metabolism in Triticum durum. Chemosphere 2009, 75, 900–905. [Google Scholar] [CrossRef]
- Bosker, T.; Bouwman, L.; Brun, N.; Behrens, P.; Vijver, M. Microplastics accumulate on pores in seed capsule and delay germination and root growth of the terrestrial vascular plant Lepidium sativum. Chemosphere 2019, 226, 774–781. [Google Scholar] [CrossRef]
- Miralles, P.; Johnson, E.; Church, T.L.; Harris, A.T. Multiwalled carbon nanotubes in alfalfa and wheat: Toxicology and uptake. J. R. Soc. Interface 2012, 9, 3514–3527. [Google Scholar] [CrossRef]
- Lin, S.; Reppert, J.; Hu, Q.; Hudson, J.S.; Reid, M.L.; Ratnikova, T.A.; Rao, A.M.; Luo, H.; Ke, P.C. Uptake, translocation, and transmission of carbon nano-materials in rice plants. Small 2009, 5, 1128–1132. [Google Scholar] [CrossRef] [PubMed]
- Pérez-de-Luque, A. Interaction of nanomaterials with plants: What do we need for real applications in agriculture? Front. Environ. Sci. 2017, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Smaka-Kincl, V.; Stegnar, P.; Lovka, M.; Toman, M.J. The evaluation of waste, surface and ground water quality using the Allium test procedure. Mutat. Res. 1996, 368, 171–179. [Google Scholar] [CrossRef]
- Fernandes, T.C.C.; Mazzeo, D.E.C.; Marin-Morales, M.A. Mechanism of micronuclei formation in polyploidizated cells of Allium cepa exposed to trifluralin herbicide. Pestic. Biochem. Phys. 2007, 88, 252–259. [Google Scholar] [CrossRef]
- Singh, D.; Roy, B.K. Evaluation of malathion-induced cytogenetical effects and oxidative stress in plants using Allium test. Acta Physiol. Plant 2017, 39, 92–102. [Google Scholar] [CrossRef]
- Kumari, M.; Khan, S.S.; Pakrashi, S.; Mukherjee, A.; Chandrasekaran, N. Cytogenetic and genotoxic effects of zinc oxide nanoparticles on root cells of Allium cepa. J. Hazard Mater. 2011, 190, 613–621. [Google Scholar] [CrossRef]
- Sharma, S.; Vig, A.P. Genotoxicity of atrazine, avenoxan, diuron and quizalofop-pethyl herbicides using the Allium cepa root chromosomal aberration assay. Terr. Aquat. Environ. Toxicol. 2012, 6, 90–95. [Google Scholar]
- Oh, N.; Park, J.H. Endocytosis and exocytosis of nanoparticles in mammalian cells. Int. J. Nanomed. 2014, 9, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Boots, B.; Russell, C.W.; Green, D.S. Effects of micro-plastics in soil ecosystems: Above and below ground. Sci. Technol. Environ. 2019, 53, 11496–11506. [Google Scholar] [CrossRef]
- Machado, A.A.S.; Kloas, W.; Zarfl, C.; Hempel, S.; Rillig, M.C. Microplastics as an emerging threat to terrestrial ecosystems. Glob. Chang. Biol. 2018, 24, 1405–1416. [Google Scholar] [CrossRef] [Green Version]
- Fatma, F.; Verma, S.; Kamal, A. Monitoring of morphotoxic, cytotoxic and genotoxic potential of mancozeb using Allium assay. Chemosphere 2018, 195, 864–870. [Google Scholar] [PubMed]
- Rajeshwari, A.; Roy, B.; Chandrasekaran, N.; Mukherjee, A. Cytogenetic evaluation of gold nanorods using Allium cepa test. Plant Physiol. Biochem. 2016, 109, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Gaulden, M.E. Hypothesis: Some mutagens directly alter specific chromosomal proteins (DNA topoisomerase II and peripheral proteins) to produce chromosome stickiness, which causes chromosome aberrations. Mutagenesis 1987, 2, 357–365. [Google Scholar] [CrossRef]
- Potapova, T.; Gorbsky, G.J. The consequences of chromosome segregation errors in mitosis and meiosis. Biology 2017, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Fiskesjö, G. Mercury and selenium in a modified Allium test. Hereditas 1979, 91, 169–178. [Google Scholar] [CrossRef]
- Evans, H.J. Molecular mechanism in the induction of chromosome aberrations. In Progress in Genetic Toxicology; Scott, D., Bridges, B.A., Sobels, F.H., Eds.; Elsevier/North-Holland and Biomedical Press: Amsterdam, The Netherland, 1977; p. 57. [Google Scholar]
- Panda, K.K.; Achary, V.M.M.; Krishnaveni, R.; Padhi, B.K.; Sarangi, S.N.; Sahu, S.N.; Panda, B.B. In vitro biosynthesis and genotoxicity bioassay of silver nanoparticles using plants. Toxicol. Vitro. 2011, 25, 1097–1105. [Google Scholar] [CrossRef]
- Gisselsson, D. Ring Chromosomes: Vicious Circles at the End and Beginning of Life; Department of Clinical Genetics, University Hospital: Lund, Sweden, 2001. [Google Scholar]
- Raghuvanshi, S.S.; Singh, A.K. Effect of gamma rays on growth and karyokinetic activity in Trigonella foenum-graecum L. Cytologia 1976, 41, 177–186. [Google Scholar]
- Vicars, H.; Karg, T.; Warecki, B.; Bast, I.; Sullivan, W. Kinetochore-independent mechanisms of sister chromosome separation. PLoS ONE Genet. 2021, 17, e1009304. [Google Scholar] [CrossRef]
- Kwon, M.; Leibowitz, M.L.; Lee, J.H. Small but mighty: The causes and consequences of micronucleus rupture. Exp. Mol. Med. 2020, 52, 1777–1786. [Google Scholar] [CrossRef]
- Fenech, M.; Chang, W.P.; Volders, M.K.; Holland, N.; Bonassi, S.; Zeiger, E. Human project: Detailed description of the scoring criteria for the cytokinesis block micronucleus assay using isolated human lymphocyte cultures. Mutat. Res. 2003, 534, 65–75. [Google Scholar] [CrossRef]
- Fernandes, T.C.C.; Mazzeo, D.E.C.; Marin-Morales, M.A. Origin of nuclear and chromosomal alterations derived from the action of an aneugenic agent e trifluralin herbicide. Ecotoxicol. Environ. Saf. 2009, 72, 1680–1686. [Google Scholar] [CrossRef] [PubMed]
- Nefic, H.; Musanovic, J.; Metovic, A.; Kurteshi, K. Chromosomal and nuclear alterations in root tip cells of Allium cepa L. Induced by alprazolam. Med. Arch. 2013, 67, 388–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ps-MPs Size (nm) | Conc. (mg/L) | TNC | IC | PRI (% ± S.E.) | MTI (% ± S.E.) | ANI (% ± S.E.) | T LI (% ± S.E.) | MI (% ± S.E.) * |
|---|---|---|---|---|---|---|---|---|
| NC | 0 | 12264 | 9199 | 39.38 ± 0.666 | 27.45 ± 0.755 | 23.49 ± 1.148 | 9.683 ± 0.935 | 25.05 ± 0.917 |
| 80 | 100 | 13036 | 11409 | 35.89 ± 1.649 | 24.26 ± 1.070 | 10.96 ± 1.848 | 4.730 ± 0.766 | 12.62 ± 0.656 |
| 400 | 13697 | 11881 | 33.04 ± 0.963 | 28.92 ± 0.967 | 10.98 ± 0.882 | 4.467 ± 0.395 | 13.32 ± 0.458 | |
| 100 | 100 | 14402 | 12671 | 35.89 ± 1.222 | 23.32 ± 0.592 | 10.41 ± 0.859 | 5.376 ± 0.344 | 12.06 ± 0.284 |
| 400 | 12655 | 10842 | 34.27 ± 0.792 | 19.35 ± 0.843 | 8.424 ± 0.733 | 6.169 ± 0.463 | 14.38 ± 0.372 | |
| 200 | 100 | 12584 | 10815 | 33.75 ± 1.388 | 19.01 ± 1.130 | 7.549 ± 0.444 | 7.306 ± 0.600 | 14.15 ± 0.332 |
| 400 | 13042 | 11241 | 31.93 ± 1.181 | 19.40 ± 0.797 | 10.53 ± 0.441 | 6.107 ± 0.381 | 14.82 ± 0.875 | |
| 500 | 100 | 12257 | 10478 | 34.95 ± 0.812 | 20.81 ± 0.885 | 9.166 ± 0.660 | 6.951 ± 0.461 | 14.56 ± 0.304 |
| 400 | 12737 | 10877 | 38.26 ± 1.353 | 20.84 ± 1.375 | 8.8 ± 0.86500 | 9.235 ± 0.411 | 14.73 ± 0.446 | |
| 1000 | 100 | 12825 | 10677 | 39.98 ± 1.556 | 26.94 ± 1.056 | 17.29 ± 1.170 | 6.053 ± 0.605 | 16.96 ± 1.068 |
| 400 | 13518 | 11638 | 45.01 ± 1.397 | 25.24 ± 0.709 | 13.87 ± 0.923 | 5.550 ± 0.797 | 13.91 ± 0.162 | |
| 2000 | 100 | 11781 | 9924 | 35.89 ± 1.418 | 27.41 ± 1.402 | 19.88 ± 0.698 | 6.16 ± 0.5460 | 15.84 ± 0.631 |
| 400 | 6857 | 5625 | 46.26 ± 2.004 | 15.85 ± 1.634 | 13.69 ± 1.949 | 5.555 ± 0.864 | 18.51 ± 1.067 | |
| 4000 | 100 | 14417 | 12548 | 41.40 ± 2.082 | 29.36 ± 2.244 | 16.28 ± 1.291 | 5.869 ± 0.650 | 13.04 ± 0.316 |
| 400 | 15046 | 12904 | 42.79 ± 0.989 | 28.36 ± 0.502 | 13.62 ± 1.048 | 5.758 ± 0.503 | 13.06 ± 0.385 | |
| 8000 | 100 | 13426 | 11611 | 41.94 ± 1.268 | 30.99 ± 1.281 | 15.01 ± 1.512 | 6.242 ± 0.920 | 13.53 ± 0.243 |
| 400 | 13751 | 11812 | 45.33 ± 1.146 | 26.58 ± 1.142 | 12.45 ± 0.845 | 5.149 ± 0.791 | 14.13 ± 0.669 |
| MP-Ps Size (nm) | Conc. (mg/L) | CM (%) | DLA (%) | CC (%) | VG (%) | LG (%) | DM (%) | DSA (%) | MP (%) |
|---|---|---|---|---|---|---|---|---|---|
| NC | 0 | ND | ND | ND | ND | ND | ND | ND | ND |
| 80 | 100 | 2.655 ± 0.465 | 1.299 ± 0.378 | 7.184 ± 0.678 | 1.739 ± 0.278 | 0.712 ± 0.083 | 3.063 ± 0.442 | 1.291 ± 0.405 | 0.736 ± 0.103 |
| 400 | 0.591 ± 0.112 | 5.213 ± 0.674 | 8.032 ± 0.908 | 1.936 ± 0.218 | 0.467 ± 0.048 | 2.115 ± 0.334 | 1.497 ± 0.208 | 0.505 ± 0.013 | |
| 100 | 100 | 1.173 ± 0.156 | 2.984 ± 0.405 | 11.86 ± 0.926 | 1.731 ± 0.212 | 0.967 ± 0.223 | 2.155 ± 0.364 | 1.182 ± 0.202 | 0.699 ± 0.172 |
| 400 | 1.689 ± 0.200 | 2.02 ± 0.2120 | 15.41 ± 0.583 | 1.978 ± 0.312 | 1.002 ± 0.082 | 2.22 ± 0.1750 | 1.907 ± 0.260 | 1.268 ± 0.149 | |
| 200 | 100 | 1.344 ± 0.166 | 2.999 ± 0.518 | 14.24 ± 0.859 | 2.276 ± 0.217 | 0.968 ± 0.195 | 1.649 ± 0.184 | 1.572 ± 0.161 | 1.009 ± 0.114 |
| 400 | 1.054 ± 0.142 | 3.674 ± 0.531 | 18.03 ± 0.770 | 3.586 ± 0.291 | 0.931 ± 0.049 | 1.473 ± 0.223 | 1.304 ± 0.286 | 0.82 ± 0.2040 | |
| 500 | 100 | 1.203 ± 0.317 | 1.807 ± 0.331 | 9.607 ± 0.608 | 1.565 ± 0.280 | 0.947 ± 0.188 | 1.576 ± 0.292 | 1.639 ± 0.143 | 1.116 ± 0.148 |
| 400 | ND | 2.118 ± 0.284 | 10.33 ± 1.146 | 1.257 ± 0.192 | 0.796 ± 0.187 | 0.880 ± 0.242 | 1.218 ± 0.339 | 1.567 ± 0.514 | |
| 1000 | 100 | 1.016 ± 0.269 | 2.066 ± 0.422 | 2.591 ± 0.472 | 0.978 ± 0.206 | 0.408 ± 0.000 | 1.165 ± 0.208 | 0.766 ± 0.197 | 0.501 ± 0.006 |
| 400 | 1.363 ± 0.247 | 2.223 ± 0.360 | 2.562 ± 0.378 | 1.15 ± 0.2400 | ND | 0.922 ± 0.189 | 1.025 ± 0.143 | 0.504 ± 0.007 | |
| 2000 | 100 | 1.385 ± 0.440 | 2.577 ± 0.359 | 3.405 ± 0.543 | 1.123 ± 0.155 | 0.877 ± 0.000 | 1.371 ± 0.262 | 1.016 ± 0.113 | 0.447 ± 0.009 |
| 400 | 2.161 ± 0.345 | 3.421 ± 0.495 | 4.241 ± 0.549 | 2.277 ± 0.362 | 0.819 ± 0.125 | 3.431 ± 0.637 | 1.003 ± 0.159 | 1.515 ± 0.000 | |
| 4000 | 100 | 0.456 ± 0.000 | 1.639 ± 0.352 | 2.063 ± 0.407 | 0.971 ± 0.083 | 0.944 ± 0.263 | 1.123 ± 0.267 | 0.918 ± 0.153 | 0.513 ± 0.000 |
| 400 | 0.841 ± 0.144 | 2.069 ± 0.389 | 2.518 ± 0.276 | 1.288 ± 0.138 | 0.68 ± 0.1360 | 1.101 ± 0.213 | 0.67 ± 0.1400 | 0.462 ± 0.019 | |
| 8000 | 100 | 1.012 ± 0.568 | 1.763 ± 0.270 | 2.099 ± 0.510 | 0.751 ± 0.157 | 0.764 ± 0.288 | 1.169 ± 0.324 | 1.378 ± 0.390 | 0.467 ± 0.000 |
| 400 | 1.127 ± 0.173 | 2.32 ± 0.181 | 3.02 ± 0.471 | 0.868 ± 0.199 | 0.587 ± 0.132 | 0.64 ± 0.1050 | 0.828 ± 0.161 | 0.462 ± 0.030 | |
| MP-Ps size (nm) | Conc. (mg/L) | DO (%) | BG (%) | BK (%) | RC (%) | CAI (%) * | MN (%) | NB (%) | NAI (%) * |
| NC | 0 | ND | ND | ND | ND | ND | ND | ND | ND |
| 80 | 100 | 0.926 ± 0.140 | 1.291 ± 0.248 | 0.698 ± 0.075 | ND | 20.08 ± 1.294 | 1.732 ± 0.432 | 2.238 ± 0.766 | 3.472 ± 0.952 |
| 400 | 0.752 ± 0.170 | 2.688 ± 0.235 | 0.725 ± 0.307 | ND | 22.54 ± 1.529 | 0.513 ± 0.026 | 1.087 ± 0.000 | 0.875 ± 0.378 | |
| 100 | 100 | 1.591 ± 0.288 | 1.499 ± 0.195 | 0.510 ± 0.000 | 0.508 ± 0.00 | 25.46 ± 1.782 | 1.106 ± 0.368 | 0.853 ± 0.222 | 0.843 ± 0.424 |
| 400 | 1.054 ± 0.113 | 2.118 ± 0.184 | 0.485 ± 0.000 | ND | 30.02 ± 0.411 | 0.882 ± 0.158 | 1.385 ± 0.428 | 1.763 ± 0.521 | |
| 200 | 100 | 1.757 ± 0.231 | 1.697 ± 0.299 | 0.540 ± 0.000 | ND | 29.47 ± 1.009 | 1.175 ± 0.211 | 1.635 ± 0.374 | 1.924 ± 0.549 |
| 400 | 1.523 ± 0.192 | 1.642 ± 0.275 | ND | ND | 31.65 ± 1.380 | 0.640 ± 0.171 | 0.487 ± 0.029 | 0.376 ± 0.106 | |
| 500 | 100 | 1.346 ± 0.201 | 1.693 ± 0.240 | 0.861 ± 0.141 | ND | 22.24 ± 0.633 | 1.698 ± 0.964 | 4.93 ± 0.9770 | 5.873 ± 1.149 |
| 400 | 1.815 ± 0.313 | 1.306 ± 0.153 | 0.490 ± 0.005 | 0.461 ± 0.00 | 20.44 ± 0.830 | 1.388 ± 0.557 | 1.846 ± 0.472 | 2.412 ± 0.540 | |
| 1000 | 100 | 0.683 ± 0.150 | 0.821 ± 0.150 | ND | ND | 9.429 ± 0.496 | 0.507 ± 0.000 | 0.741 ± 0.184 | 0.303 ± 0.330 |
| 400 | 1.246 ± 0.210 | 1.191 ± 0.196 | ND | 0.53 ± 0.029 | 9.892 ± 0.601 | 0.627 ± 0.173 | 0.69 ± 0.2140 | 0.438 ± 0.124 | |
| 2000 | 100 | 0.797 ± 0.129 | 0.651 ± 0.103 | 0.555 ± 0.099 | 0.416 ± 0.00 | 10.3 ± 0.8480 | 0.88 ± 0.1040 | 0.563 ± 0.083 | 0.704 ± 0.247 |
| 400 | 0.869 ± 0.000 | 1.003 ± 0.253 | 1.013 ± 0.143 | ND | 17.29 ± 1.645 | 2.43 ± 0.5890 | 1.127 ± 0.407 | 2.081 ± 0.855 | |
| 4000 | 100 | 0.487 ± 0.018 | 2.072 ± 1.479 | ND | 0.502 ± 0.00 | 6.759 ± 0.562 | 0.683 ± 0.186 | 0.497 ± 0.029 | 0.338 ± 0.273 |
| 400 | 0.565 ± 0.095 | 0.635 ± 0.085 | ND | ND | 9.210 ± 0.563 | 0.588 ± 0.127 | ND | 0.261 ± 0.128 | |
| 8000 | 100 | 0.528 ± 0.013 | 1.111 ± 0.219 | ND | ND | 7.05 ± 0.8750 | 0.511 ± 0.019 | ND | 0.171 ± 0.019 |
| 400 | 0.461 ± 0.020 | 1.18 ± 0.214 | 0.469 ± 0.000 | 0.417 ± 0.00 | 10.16 ± 0.503 | 0.467 ± 0.028 | 0.555 ± 0.000 | 0.217 ± 0.030 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, M.; Xu, M.; Wang, L. Cyto–Genotoxic Effect Causing Potential of Polystyrene Micro-Plastics in Terrestrial Plants. Nanomaterials 2022, 12, 2024. https://doi.org/10.3390/nano12122024
Kaur M, Xu M, Wang L. Cyto–Genotoxic Effect Causing Potential of Polystyrene Micro-Plastics in Terrestrial Plants. Nanomaterials. 2022; 12(12):2024. https://doi.org/10.3390/nano12122024
Chicago/Turabian StyleKaur, Mandeep, Ming Xu, and Lin Wang. 2022. "Cyto–Genotoxic Effect Causing Potential of Polystyrene Micro-Plastics in Terrestrial Plants" Nanomaterials 12, no. 12: 2024. https://doi.org/10.3390/nano12122024
APA StyleKaur, M., Xu, M., & Wang, L. (2022). Cyto–Genotoxic Effect Causing Potential of Polystyrene Micro-Plastics in Terrestrial Plants. Nanomaterials, 12(12), 2024. https://doi.org/10.3390/nano12122024