Review of Copper and Copper Nanoparticle Toxicity in Fish


 ,
,  ,
, 

Abstract
1. Introduction
2. Overview of Copper-Induced Toxicity in Aquatic Model Organisms
3. Overview of Copper-Induced Organ Toxicity in Elected Fish
4. Effect of External Factors on Copper Toxicity in Aquatic Organisms
5. Effects of Copper Nanoparticles (CuNPs) on Fish
6. Overview of Organ Toxicity in Fish Induced by Copper and Copper Oxide Nanoparticles
7. Effect of External Factors on CuNP and CuO-NP Toxicity in Fish
8. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lushina, M.; Kamenev, Y.; Leonov, V.; Ostapenko, E. Use of expanded copper mesh grid for negative electrodes of sealed lead storage batteries. J. Power Sources 2005, 148, 95–104. [Google Scholar] [CrossRef]
- Salem, I.A.; Shaltout, M.H.; Zaki, A.B. Homogeneous and heterogeneous catalytic oxidation of some azo dyes using copper(II) ions. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2019, 227, 117618. [Google Scholar] [CrossRef] [PubMed]
- Mitrea, S.; Oprina, G.; Radu, E.; Marinescu, V.; Voina, A.; Lingvay, I. Corrosion of copper and carbon steel in some electrical purposes oils. Rev. Chim.(Bucharest) 2016, 67, 1707–1712. [Google Scholar]
- Gupta, N.; Siddique, R. Utilization of Copper Slag in Self-compacting Concrete—Strength and Permeation Properties. In Natural Fibres: Advances in Science and Technology Towards Industrial Applications; Springer: Berlin/Heidelberg, Germany, 2019; pp. 544–551. [Google Scholar]
- Inkinen, J.; Mäkinen, R.; Keinänen-Toivola, M.M.; Nordström, K.; Ahonen, M. Copper as an antibacterial material in different facilities. Lett. Appl. Microbiol. 2016, 64, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Dugal, S.; Mascarenhas, S. Chemical synthesis of copper nanoparticles and its antibacterial effect against gram negative pathogens. J. Adv. Sci. Res. 2015, 6, 1–4. [Google Scholar]
- Lennox, A.J.J.; Bartels, P.; Pohl, M.-M.; Junge, H.; Beller, M. In situ photodeposition of copper nanoparticles on TiO2: Novel catalysts with facile light-induced redox cycling. J. Catal. 2016, 340, 177–183. [Google Scholar] [CrossRef]
- Barot, N.; Patel, S.B.; Kaur, H. Nitro resin supported copper nanoparticles: An effective heterogeneous catalyst for C N cross coupling and oxidative C C homocoupling. J. Mol. Catal. A Chem. 2016, 423, 77–84. [Google Scholar] [CrossRef]
- Woźniak-Budych, M.J.; Langer, K.; Peplińska, B.; Przysiecka, Ł.; Jarek, M.; Jarzębski, M.; Jurga, S. Copper-gold nanoparticles: Fabrication, characteristic and application as drug carriers. Mater. Chem. Phys. 2016, 179, 242–253. [Google Scholar] [CrossRef]
- Albrecht, A.; Rivadeneyra, A.; Abdellah, A.; Lugli, P.; Fernández-Salmerón, J. Inkjet printing and photonic sintering of silver and copper oxide nanoparticles for ultra-low-cost conductive patterns. J. Mater. Chem. C 2016, 4, 3546–3554. [Google Scholar] [CrossRef]
- Tsai, C.-Y.; Chang, W.-C.; Chen, G.-L.; Chung, C.-H.; Liang, J.-X.; Ma, W.-Y.; Yang, T.-N. A Study of the Preparation and Properties of Antioxidative Copper Inks with High Electrical Conductivity. Nanoscale Res. Lett. 2015, 10, 357. [Google Scholar] [CrossRef]
- Gopalan, A.; Muthuchamy, N.; Komathi, S.; Lee, K.-P. A novel multicomponent redox polymer nanobead based high performance non-enzymatic glucose sensor. Biosens. Bioelectron. 2016, 84, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Brahman, P.K.; Suresh, L.; Lokesh, V.; Nizamuddin, S. Fabrication of highly sensitive and selective nanocomposite film based on CuNPs/fullerene-C60/MWCNTs: An electrochemical nanosensor for trace recognition of paracetamol. Anal. Chim. Acta 2016, 917, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Pourbeyram, S.; Mehdizadeh, K. Nonenzymatic glucose sensor based on disposable pencil graphite electrode modified by copper nanoparticles. J. Food Drug Anal. 2016, 24, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-W.; Lee, I.-C.; Shin, N.-R.; Jeon, C.-M.; Kwon, O.-K.; Ko, J.-W.; Kim, J.-C.; Oh, S.R.; Shin, I.-S.; Ahn, K.-S. Copper oxide nanoparticles aggravate airway inflammation and mucus production in asthmatic mice via MAPK signaling. Nanotoxicology 2015, 10, 1–8. [Google Scholar] [CrossRef]
- Montes, M.; Pierce, C.G.; Lopez-Ribot, J.L.; Bhalla, A.; Guo, R. Properties of Silver and Copper Nanoparticle Containing Aqueous Suspensions and Evaluation of their In Vitro Activity against Candida albicans and Staphylococcus aureus Biofilms. J. Nano Res. 2015, 37, 109–121. [Google Scholar] [CrossRef]
- Kalatehjari, P.; Yousefian, M.; Khalilzadeh, M.A. Assessment of antifungal effects of copper nanoparticles on the growth of the fungus Saprolegnia sp. on white fish (Rutilus frisii kutum) eggs. Egypt. J. Aquat. Res. 2015, 41, 303–306. [Google Scholar] [CrossRef]
- Majumder, S.; Neogi, S.; Soumyadip, M.; Sudarsan, N. Antimicrobial Activity of Copper Oxide Nanoparticles Coated on Cotton Fabric and Synthesized by One-Pot Method. Adv. Sci. Eng. Med. 2016, 8, 102–111. [Google Scholar] [CrossRef]
- Ma, W.; Soroush, A.; Luong, T.V.A.; Brennan, G.; Rahaman, S.; Asadishad, B.; Tufenkji, N. Spray- and spin-assisted layer-by-layer assembly of copper nanoparticles on thin-film composite reverse osmosis membrane for biofouling mitigation. Water Res. 2016, 99, 188–199. [Google Scholar] [CrossRef]
- Ben-Sasson, M.; Lu, X.; Nejati, S.; Jaramillo, H.; Elimelech, M. In situ surface functionalization of reverse osmosis membranes with biocidal copper nanoparticles. Desalination 2016, 388, 1–8. [Google Scholar] [CrossRef]
- Förstner, U.; Wittmann, G.T. Metal Pollution in the Aquatic Environment; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Lee, J.; Prohaska, J.R.; Thiele, D.J. Essential role for mammalian copper transporter Ctr1 in copper homeostasis and embryonic development. Proc. Natl. Acade. Sci. USA 2001, 98, 6842–6847. [Google Scholar] [CrossRef]
- Lee, J.; Petris, M.J.; Thiele, D.J. Characterization of mouse embryonic cells deficient in the ctr1 high affinity copper transporter Identification of a Ctr1-independent copper transport system. J. Biol. Chem. 2002, 277, 40253–40259. [Google Scholar] [CrossRef] [PubMed]
- Stern, B.R. Essentiality and Toxicity in Copper Health Risk Assessment: Overview, Update and Regulatory Considerations. J. Toxicol. Environ. Heal. Part A 2010, 73, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kiron, V.; Satoh, S. Trace minerals in fish nutrition. Aquaculture 1997, 151, 185–207. [Google Scholar] [CrossRef]
- Scott, D.M.; Major, C.W. The Effect of Copper (Ii) on Survival, Respiration, and Heart Rate in The Common Blue Mussel, Mytilus Edulis. Boil. Bull. 1972, 143, 679–688. [Google Scholar] [CrossRef]
- Flemming, C.A.; Trevors, J.T. Copper toxicity and chemistry in the environment: A review. Water Air Soil Pollut. 1989, 44, 143–158. [Google Scholar] [CrossRef]
- Richey, D.; Roseboom, D. Acute toxicity of copper to some fishes in high alkalinity water. Circular 1978. [Google Scholar] [CrossRef]
- Moore, J.W.; Ramamoorthy, S. Heavy Metals in Natural Waters: Applied Monitoring and Impact Assessment; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Tait, T.N.; McGeer, J.C.; Smith, D.S. Testing the Underlying Chemical Principles of the Biotic Ligand Model (BLM) to Marine Copper Systems: Measuring Copper Speciation Using Fluorescence Quenching. Bull. Environ. Contam. Toxicol. 2017, 100, 76–81. [Google Scholar] [CrossRef]
- Moffett, J.W.; Dupont, C. Cu complexation by organic ligands in the sub-arctic NW Pacific and Bering Sea. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 586–595. [Google Scholar] [CrossRef]
- Zitoun, R. Copper Speciation in Different Marine Ecosystems around New Zealand; University of Otago: Dunedin, New Zealand, 2019. [Google Scholar]
- Donat, J.R.; Lao, K.A.; Bruland, K.W. Speciation of dissolved copper and nickel in South San Francisco Bay: A multi-method approach. Anal. Chim. Acta 1994, 284, 547–571. [Google Scholar] [CrossRef]
- Moffett, J.W.; Tuit, C.B.; Ward, B.B. Chelator-induced inhibition of copper metalloenzymes in denitrifying bacteria. Limnol. Oceanogr. 2012, 57, 272–280. [Google Scholar] [CrossRef]
- Oldham, V.E.; Swenson, M.; Buck, K. Spatial variability of total dissolved copper and copper speciation in the inshore waters of Bermuda. Mar. Pollut. Bull. 2014, 79, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Azizi, Z.; Alamdari, A.; Malayeri, M.R. Thermal performance and friction factor of a cylindrical microchannel heat sink cooled by Cu-water nanofluid. Appl. Therm. Eng. 2016, 99, 970–978. [Google Scholar] [CrossRef]
- Keller, A.A.; Adeleye, A.S.; Conway, J.R.; Garner, K.L.; Zhao, L.; Cherr, G.N.; Hong, J.; Gardea-Torresdey, J.L.; Godwin, H.A.; Hanna, S.; et al. Comparative environmental fate and toxicity of copper nanomaterials. NanoImpact 2017, 7, 28–40. [Google Scholar] [CrossRef]
- Syed, M.; Coombs, T.L. Copper metabolism in the plaice, Pleuronectes platessa (L.). J. Exp. Mar. Boil. Ecol. 1982, 63, 281–296. [Google Scholar] [CrossRef]
- Heath, A.G. Effects of waterborne copper or zinc on the osmoregulatory response of bluegill to a hypertonic NaCl challenge. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1987, 88, 307–311. [Google Scholar] [CrossRef]
- De Romaña, D.L.; Olivares, M.; Uauy, R.; Araya, M. Risks and benefits of copper in light of new insights of copper homeostasis. J. Trace Elements Med. Boil. 2011, 25, 3–13. [Google Scholar] [CrossRef]
- Russell, R.; Beard, J.L.; Cousins, R.J.; Dunn, J.T.; Ferland, G.; Hambidge, K.; Lynch, S.; Penland, J.; Ross, A.; Stoecker, B. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; A Report of the Panel on Micronutrients; Subcommittees on Upper Reference Levels of Nutrients and of Interpretation and Uses of Dietary Reference Intakes, and the Standing Committee on the Scientific Evaluation of Dietary Reference Intakes Food and Nutrition Board Institute of Medicine: Washington, WA, USA, 2001. [Google Scholar]
- Turnlund, J.R.; Keyes, W.R.; Kim, S.K.; Domek, J.M. Long-term high copper intake: Effects on copper absorption, retention, and homeostasis in men. Am. J. Clin. Nutr. 2005, 81, 822–828. [Google Scholar] [CrossRef]
- Pelgrom, S.; Lock, R.; Balm, P.; Bonga, S. Integrated physiological response of tilapia, Oreochromis mossambicus, to sublethal copper exposure. Aquat. Toxicol. 1995, 32, 303–320. [Google Scholar] [CrossRef]
- Eisler, R. Handbook of Chemical Risk Assessment: Health Hazards to Humans, Plants, and Animals, Three Volume Set; CRC press: Boca Raton, FL, USA, 2000; Volume 1. [Google Scholar]
- Singh, A.; Singh, D.; Yadav, H. Impact and assessment of heavy metal toxicity on water quality, edible fishes and sediments in lakes: A review. Trends. Biosci. 2017, 10, 1551–1560. [Google Scholar]
- Pagenkopf, G.K. Gill surface interaction model for trace-metal toxicity to fishes: Role of complexation, pH, and water hardness. Environ. Sci. Technol. 1983, 17, 342–347. [Google Scholar] [CrossRef]
- Erickson, R.J.; Benoit, D.A.; Mattson, V.R. A Prototype Toxicity Factors Model for Site-Specific Copper Water Quality Criteria; US Environmental Protection Agency, Environmental Research Laboratory: Duluth, Minnesota, 1996. [Google Scholar]
- Erickson, R.J.; Benoit, D.A.; Mattson, V.R.; Leonard, E.N.; Nelson, H.P., Jr. The effects of water chemistry on the toxicity of copper to fathead minnows. Environ. Toxicol. Chem. Int. J. 1996, 15, 181–193. [Google Scholar] [CrossRef]
- Welsh, P.G.; Skidmore, J.F.; Spry, D.J.; Dixon, D.G.; Hodson, P.V.; Hutchinson, N.J.; Hickie, B.E. Effect of pH and Dissolved Organic Carbon on the Toxicity of Copper to Larval Fathead Minnow (Pimephales promelas) in Natural Lake Waters of Low Alkalinity. Can. J. Fish. Aquat. Sci. 1993, 50, 1356–1362. [Google Scholar] [CrossRef]
- Santore, R.C.; Di Toro, D.M.; Paquin, P.R.; Allen, H.E.; Meyer, J.S. Biotic ligand model of the acute toxicity of metals. 2. Application to acute copper toxicity in freshwater fish and Daphnia. Environ. Toxicol. Chem. Int. J. 2001, 20, 2397–2402. [Google Scholar] [CrossRef]
- Förstner, U.; Wittmann, G.T.W.; Förstner, P.D.U. Metal Pollution in the Aquatic Environment; Springer: Berlin/Heidelberg, Germany, 1981. [Google Scholar]
- Pieniak, Z.; Verbeke, W.; Scholderer, J. Health-related beliefs and consumer knowledge as determinants of fish consumption. J. Hum. Nutr. Diet. 2010, 23, 480–488. [Google Scholar] [CrossRef]
- Handy, R.D. The assessment of episodic metal pollution. I. Uses and limitations of tissue contaminant analysis in rainbow trout (Oncorhynchus mykiss) after short waterborne exposure to cadmium or copper. Arch. Environ. Contam. Toxicol. 1992, 22, 74–81. [Google Scholar] [CrossRef]
- Paquin, P.R.; Gorsuch, J.W.; Apte, S.; Batley, G.E.; Bowles, K.C.; Campbell, P.G.; Delos, C.G.; Di Toro, D.M.; Dwyer, R.L.; Galvez, F.; et al. The biotic ligand model: A historical overview. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2002, 133, 3–35. [Google Scholar] [CrossRef]
- De Schamphelaere, K.; Janssen, C.R. A Biotic Ligand Model Predicting Acute Copper Toxicity forDaphnia magna: The Effects of Calcium, Magnesium, Sodium, Potassium, and pH. Environ. Sci. Technol. 2002, 36, 48–54. [Google Scholar] [CrossRef]
- Bianchini, A.; Martins, S.E.; Barcarolli, I. Mechanism of acute copper toxicity in euryhaline crustaceans: Implications for the Biotic Ligand Model. Int. Congr. Ser. 2004, 1275, 189–194. [Google Scholar] [CrossRef]
- Playle, R.C.; Gensemer, R.W.; Dixon, D.G. Copper accumulation on gills of fathead minnows: Influence of water hardness, complexation and pH of the gill micro-environment. Environ. Toxicol. Chem. Int. J. 1992, 11, 381–391. [Google Scholar] [CrossRef]
- Playle, R.C.; Dixon, D.G.; Burnison, K. Copper and Cadmium Binding to Fish Gills: Estimates of Metal–Gill Stability Constants and Modelling of Metal Accumulation. Can. J. Fish. Aquat. Sci. 1993, 50, 2678–2687. [Google Scholar] [CrossRef]
- Davis, R.A., Jr.; Welty, A.T.; Borrego, J.; Morales, J.A.; Pendon, J.G.; Ryan, J. Rio Tinto estuary (Spain): 5000 years of pollution. Environ. Earth Sci. 2000, 39, 1107–1116. [Google Scholar] [CrossRef]
- Hall, W.S.; Bushong, S.J.; Hall, L.W.; Lenkevich, M.J.; Pinkney, A.E. Monitoring dissolved copper concentrations in Chesapeake Bay, U.S.A. Environ. Monit. Assess. 1988, 11, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, E.M. Metal Poisoning in Fish; CRC press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Carbonell, G.; Tarazona, J. Toxicokinetics of copper in rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 1994, 29, 213–221. [Google Scholar] [CrossRef]
- Kapustka, L.A. Issue Paper on the Ecological Effects of Metals; US Environmental Protection Agency: Washington, DC, USA, 2004. [Google Scholar]
- Tierney, K.B.; Baldwin, D.H.; Hara, T.J.; Ross, P.S.; Scholz, N.L.; Kennedy, C.J. Olfactory toxicity in fishes. Aquat. Toxicol. 2010, 96, 2–26. [Google Scholar] [CrossRef] [PubMed]
- Nriagu, J.O. A History of Global Metal Pollution. Science 1996, 272, 223. [Google Scholar] [CrossRef]
- Woody, C.A.; O’Neal, S. Effects of Copper on Fish and Aquatic Resources; Fisheries Research and Consulting: Anchorage, Alaska, 2012. [Google Scholar]
- Baker, J.T.P. Histological and Electron Microscopical Observations on Copper Poisoning in the Winter Flounder (Pseudopleuronectes americanus). J. Fish. Res. Board Can. 1969, 26, 2785–2793. [Google Scholar] [CrossRef]
- Cardeilhac, P.; Simpson, C.; Lovelock, R.; Yosha, S.; Calderwood, H.; Gudat, J. Failure of osmoregulation with apparent potassium intoxication in marine teleosts: A primary toxic effect of copper. Aquaculture 1979, 17, 231–239. [Google Scholar] [CrossRef]
- Benedetti, I.; Albano, A.G.; Mola, L. Histomorphological changes in some organs of the brown bullhead, Ictalurus nebulosus LeSueur, following short- and long-term exposure to copper. J. Fish Boil. 1989, 34, 273–280. [Google Scholar] [CrossRef]
- Mazon, A.F.; Monteiro, E.A.S.; Pinheiro, G.H.D.; Fernadez, M.N. Hematological and physiological changes induced by short-term exposure to copper in the freshwater fish, Prochilodus scrofa. Braz. J. Boil. 2002, 62, 621–631. [Google Scholar] [CrossRef]
- Figueiredo-Fernandes, A.; Ferreira-Cardoso, J.V.; Santos, S.G.G.; Monteiro, S.M.; Carrola, J.; Matos, P. Histopathological changes in liver and gill epithelium of Nile tilapia, Oreochromis niloticus, exposed to waterborne copper. Pesqui. Veterinária Bras. 2007, 27, 103–109. [Google Scholar] [CrossRef]
- Shaw, B.; Handy, R.D. Dietary copper exposure and recovery in Nile tilapia, Oreochromis niloticus. Aquat. Toxicol. 2006, 76, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Kamunde, C.N.; Grosell, M.; Na Lott, J.; Wood, C.M. Copper metabolism and gut morphology in rainbow trout (Oncorhynchus mykiss) during chronic sublethal dietary copper exposure. Can. J. Fish. Aquat. Sci. 2001, 58, 293–305. [Google Scholar] [CrossRef]
- De Boeck, G.; Vlaeminck, A.; Balm, P.H.; Lock, R.A.; De Wachter, B.; Blust, R. Morphological and metabolic changes in common carp, Cyprinus carpio, during short-term copper exposure: Interactions between Cu2+ and plasma cortisol elevation. Environ. Toxicol. Chem. Int. J. 2001, 20, 374–381. [Google Scholar] [CrossRef]
- McGeer, J.C.; Szebedinszky, C.; McDonald, D.G.; Wood, C.M. The role of dissolved organic carbon in moderating the bioavailability and toxicity of Cu to rainbow trout during chronic waterborne exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2002, 133, 147–160. [Google Scholar] [CrossRef]
- Marr, J.C.; Lipton, J.; Cacela, D.; A Hansen, J.; Meyer, J.S.; Bergman, H.L. Bioavailability and acute toxicity of copper to rainbow trout (Oncorhynchus mykiss) in the presence of organic acids simulating natural dissolved organic carbon. Can. J. Fish. Aquat. Sci. 1999, 56, 1471–1483. [Google Scholar] [CrossRef]
- Schjolden, J.; Sørensen, J.; Nilsson, G.E.; Poléo, A.B. The toxicity of copper to crucian carp (Carassius carassius) in soft water. Sci. Total. Environ. 2007, 384, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Oliva, M.; Garrido-Pérez, M.D.C.; Márquez, D.S.; De Canales, M.G. Sublethal and lethal toxicity in juvenile Senegal sole (Solea senegalensis) exposed to copper: A preliminary toxicity range-finding test. Exp. Toxicol. Pathol. 2009, 61, 113–121. [Google Scholar] [CrossRef]
- Jiraungkoorskul, W.; Sahaphong, S.; Kangwanrangsan, N. Toxicity of copper in butterfish (Poronotus triacanthus): Tissues accumulation and ultrastructural changes. Environ. Toxicol. 2007, 22, 92–100. [Google Scholar] [CrossRef]
- Cerqueira, C.C.; Fernandes, M.N. Gill Tissue Recovery after Copper Exposure and Blood Parameter Responses in the Tropical Fish Prochilodus scrofa. Ecotoxicol. Environ. Saf. 2002, 52, 83–91. [Google Scholar] [CrossRef]
- Mela, M.; Guiloski, I.; Doria, H.B.; Rabitto, I.; Silva, C.; Maraschi, A.; Prodocimo, V.; Freire, C.; Randi, M.A.F.; Ribeiro, C.A.D.O.; et al. Risks of waterborne copper exposure to a cultivated freshwater Neotropical catfish (Rhamdia quelen). Ecotoxicol. Environ. Saf. 2013, 88, 108–116. [Google Scholar] [CrossRef]
- Mazon, A.D.F.; Fernandes, M.; Nolasco, M.; Severi, W. Functional morphology of gills and respiratory area of two active rheophilic fish species, Plagioscion squamosissimus and Prochilodus scrofa. J. Fish Boil. 1998, 52, 50–61. [Google Scholar] [CrossRef]
- Monteiro, S.M.; Mancera, J.M.; Fontaínhas-Fernandes, A.; Sousa, M. Copper induced alterations of biochemical parameters in the gill and plasma of Oreochromis niloticus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2005, 141, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Viarengo, A.; Pertica, M.; Mancinelli, G.; Burlando, B.; Canesi, L.; Orunesu, M. In vivo effects of copper on the calcium homeostasis mechanisms of mussel gill cell plasma membranes. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1996, 113, 421–425. [Google Scholar] [CrossRef]
- Laurén, D.J.; McDonald, D.G. Influence of Water Hardness, pH, and Alkalinity on the Mechanisms of Copper Toxicity in Juvenile Rainbow Trout, Salmo gairdneri. Can. J. Fish. Aquat. Sci. 1986, 43, 1488–1496. [Google Scholar] [CrossRef]
- Evans, D.H. The fish gill: Site of action and model for toxic effects of environmental pollutants. Environ. Health Perspect. 1987, 71, 47–58. [Google Scholar] [CrossRef]
- Eyckmans, M.; Tudorache, C.; Darras, V.M.; Blust, R.; De Boeck, G. Hormonal and ion regulatory response in three freshwater fish species following waterborne copper exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 152, 270–278. [Google Scholar] [CrossRef]
- Sola, F.; Isaia, J.; Masoni, A. Effects of copper on gill structure and transport function in the rainbow trout, oncorhynchus mykiss. J. Appl. Toxicol. 1995, 15, 391–398. [Google Scholar] [CrossRef]
- Grosell, M.; McDonald, M.; Walsh, P.; Wood, C.M. Effects of prolonged copper exposure in the marine gulf toadfish (Opsanus beta) II: Copper accumulation, drinking rate and Na+/K+-ATPase activity in osmoregulatory tissues. Aquat. Toxicol. 2004, 68, 263–275. [Google Scholar] [CrossRef]
- Monteiro, S.M.; Rocha, E.; Fontaínhas-Fernandes, A.; Sousa, M. Quantitative histopathology ofOreochromis niloticusgills after copper exposure. J. Fish Boil. 2008, 73, 1376–1392. [Google Scholar] [CrossRef]
- McNulty, H.R.; Anderson, B.S.; Hunt, J.W.; Turpen, S.L.; Singer, M.M. Age-specific toxicity of copper to larval topsmeltAther1Nops Affinis. Environ. Toxicol. Chem. 1994, 13, 487–492. [Google Scholar] [CrossRef]
- Taylor, L.N.; Wood, C.M.; McDonald, D.G. An Evaluation of Sodium Loss and Gill Metal Binding Properties in Rainbow Trout and Yellow Perch to Explain Species Differences in Copper Tolerance. Environ. Toxicol. Chem. 2003, 22, 2159–2166. [Google Scholar] [CrossRef] [PubMed]
- Roméo, M.; Cosson, R.P.; Gnassia-Barelli, M.; Risso, C.; Stien, X.; Lafaurie, M. Metallothionein determination in the liver of the sea bass Dicentrarchus labrax treated with copper and B(a)P. Mar. Environ. Res. 1997, 44, 275–284. [Google Scholar] [CrossRef]
- Segner, H. Response of fed and starved roach, Rutilus rutilus, to sublethal copper contamination. J. Fish Boil. 1987, 30, 423–437. [Google Scholar] [CrossRef]
- Paris-Palacios, S.; Biagianti-Risbourg, S.; Vernet, G. Biochemical and (ultra)structural hepatic perturbations of Brachydanio rerio (Teleostei, Cyprinidae) exposed to two sublethal concentrations of copper sulfate. Aquat. Toxicol. 2000, 50, 109–124. [Google Scholar] [CrossRef]
- Al-Bairuty, G.A.; Shaw, B.J.; Handy, R.D.; Henry, T.B. Histopathological effects of waterborne copper nanoparticles and copper sulphate on the organs of rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2013, 126, 104–115. [Google Scholar] [CrossRef]
- Carvalho, C.D.S.; Fernandes, M.N. Effect of copper on liver key enzymes of anaerobic glucose metabolism from freshwater tropical fish Prochilodus lineatus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 151, 437–442. [Google Scholar] [CrossRef]
- Patel, J.; Bahadur, A. Histopathological manifestations of sub lethal toxicity of copper ions in Catla catla. Am. Eurasian J. Toxicol. Sci. 2011, 3, 1–5. [Google Scholar]
- Sanchez, W.; Palluel, O.; Meunier, L.; Coquery, M.; Porcher, J.-M.; Aït-Aïssa, S. Copper-induced oxidative stress in three-spined stickleback: Relationship with hepatic metal levels. Environ. Toxicol. Pharmacol. 2005, 19, 177–183. [Google Scholar] [CrossRef]
- Romeo, M.; Bennani, N.; Gnassia-Barelli, M.; Lafaurie, M.; Girard, J. Cadmium and copper display different responses towards oxidative stress in the kidney of the sea bass Dicentrarchus labrax. Aquat. Toxicol. 2000, 48, 185–194. [Google Scholar] [CrossRef]
- Kondera, E.; Witeska, M. Cadmium and copper reduce hematopoietic potential in common carp (Cyprinus carpio L.) head kidney. Fish Physiol. Biochem. 2012, 39, 755–764. [Google Scholar] [CrossRef]
- Kondera, E.; Ługowska, K.; Sarnowski, P. High affinity of cadmium and copper to head kidney of common carp (Cyprinus carpio L.). Fish Physiol. Biochem. 2013, 40, 9–22. [Google Scholar] [CrossRef] [PubMed]
- James, R.; Sampath, K.; Jothilakshmi, S.; Vasudhevan, I.; Thangarathinam, R. Effects of copper toxicity on growth, reproduction and metal accumulation in chosen ornamental fishes. Ecohydrol. Hydrobiol. 2008, 8, 89–97. [Google Scholar] [CrossRef]
- Wong, P.; Chu, L.; Wong, C. Study of toxicity and bioaccumulation of copper in the silver sea bream Sparus sarba. Environ. Int. 1999, 25, 417–422. [Google Scholar] [CrossRef]
- Papagiannis, I.; Kagalou, I.; Leonardos, J.; Petridis, D.; Kalfakakou, V. Copper and zinc in four freshwater fish species from Lake Pamvotis (Greece). Environ. Int. 2004, 30, 357–362. [Google Scholar] [CrossRef]
- Kumar, S.; Pant, S. Comparative effects of the sublethal poisoning of zinc, copper and lead on the gonads of the teleost Puntius conchonius ham. Toxicol. Lett. 1984, 23, 189–194. [Google Scholar] [CrossRef]
- Ellenberger, S.A.; Baumann, P.C.; May, T.W. Evaluation of Effects Caused by High Copper Concentrations in Torch Lake, Michigan, on Reproduction of Yellow Perch. J. Great Lakes Res. 1994, 20, 531–536. [Google Scholar] [CrossRef]
- Kotze, P.; Du Preez, H.; Van Vuren, J. Bioaccumulation of copper and zinc in Oreochromis mossambicus and Clarias gariepinus, from the Olifants River, Mpumalanga, South Africa. Water Sa Pretoria 1999, 25, 99–110. [Google Scholar]
- Zubcov, E.; Zubcov, N.; Ene, A.; Biletchi, L. Assessment of copper and zinc levels in fish from freshwater ecosystems of Moldova. Environ. Sci. Pollut. Res. 2012, 19, 2238–2247. [Google Scholar] [CrossRef]
- Suvi, R.; Giovanna, M.; Katja, A. Transgenerational endocrine disruption? Experimental copper exposure, but not heat stress, leads to elevated egg thyroid hormone levels. bioRxiv 2019, 717157. [Google Scholar] [CrossRef]
- Hughes, G.; Nemcsók, J. Effects of low pH alone and combined with copper sulphate on blood parameters of rainbow trout. Environ. Pollut. 1988, 55, 89–95. [Google Scholar] [CrossRef]
- Nemcsók, J.; Nemeth, A.; Buzás, Z.; Boross, L. Effects of copper, zinc and paraquat on acetylcholinesterase activity in carp (Cyprinus carpio L.). Aquat. Toxicol. 1984, 5, 23–31. [Google Scholar] [CrossRef]
- Stasiūnaitė, P. Toxicity of Copper to Embryonic Development of Rainbow Trout (Oncorhynchus Mykiss). Acta Zool. Litu. 2005, 15, 259–265. [Google Scholar] [CrossRef]
- Wilson, R.W.; Taylor, E.W. The physiological responses of freshwater rainbow trout, Oncorhynchus mykiss, during acutely lethal copper exposure. J. Comp. Physiol. B 1993, 163, 38–47. [Google Scholar] [CrossRef]
- Johnson, A.; Carew, E.; Sloman, K.A. The effects of copper on the morphological and functional development of zebrafish embryos. Aquat. Toxicol. 2007, 84, 431–438. [Google Scholar] [CrossRef]
- De Boeck, G.; Nilsson, G.E.; Elofsson, U.; Vlaeminck, A.; Blust, R. Brain monoamine levels and energy status in common carp (Cyprinus carpio) after exposure to sublethal levels of copper. Aquat. Toxicol. 1995, 33, 265–277. [Google Scholar] [CrossRef]
- Scarfe, A.; Jones, K.; Steele, C.; Kleerekoper, H.; Corbett, M. Locomotor behavior of four marine teleosts in response to sublethal copper exposure. Aquat. Toxicol. 1982, 2, 335–353. [Google Scholar] [CrossRef]
- Steele, C.W. Effects of exposure to sublethal copper on the locomotor behavior of the sea catfish, Arius felis. Aquat. Toxicol. 1983, 4, 83–93. [Google Scholar] [CrossRef]
- Koltes, K.H. Effects of Sublethal Copper Concentrations on the Structure and Activity of Atlantic Silverside Schools. Trans. Am. Fish. Soc. 1985, 114, 413–422. [Google Scholar] [CrossRef]
- Lett, P.F.; Farmer, G.J.; Beamish, F.W.H. Effect of Copper on Some Aspects of the Bioenergetics of Rainbow Trout (Salmo gairdneri). J. Fish. Res. Board Can. 1976, 33, 1335–1342. [Google Scholar] [CrossRef]
- Drummond, R.A.; Spoor, W.A.; Olson, G.F. Some Short-term Indicators of Sublethal Effects of Copper on Brook Trout, Salvelinus fontinalis. J. Fish. Res. Board Can. 1973, 30, 698–701. [Google Scholar] [CrossRef]
- Sandheinrich, M.B.; Atchison, G.J. Sublethal Copper Effects on Bluegill, Lepomis macrochirus, Foraging Behavior. Can. J. Fish. Aquat. Sci. 1989, 46, 1977–1985. [Google Scholar] [CrossRef]
- Pyle, G.G.; Mirza, R.S. Copper-Impaired Chemosensory Function and Behavior in Aquatic Animals. Hum. Ecol. Risk Assessment: Int. J. 2007, 13, 492–505. [Google Scholar] [CrossRef][Green Version]
- Kleerekoper, H.; Waxman, J.B.; Matis, J. Interaction of Temperature and Copper Ions as Orienting Stimuli in the Locomotor Behavior of the Goldfish (Carassius auratus). J. Fish. Res. Board Can. 1973, 30, 725–728. [Google Scholar] [CrossRef]
- Benoit, D. Chronic Effects of Copper on Survival, Growth, and Reproduction of the Bluegill (Lepomis macrochirus). Trans. Am. Fish. Soc. 1975, 104, 353–358. [Google Scholar] [CrossRef]
- Sloman, K.A.; Baker, D.W.; Wood, C.M.; McDonald, G. Social interactions affect physiological consequences of sublethal copper exposure in rainbow trout, Oncorhynchus mykiss. Environ. Toxicol. Chem. Int. J. 2002, 21, 1255–1263. [Google Scholar] [CrossRef]
- Audira, G.; Sampurna, B.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A Simple Setup to Perform 3D Locomotion Tracking in Zebrafish by Using a Single Camera. Inventions 2018, 3, 11. [Google Scholar] [CrossRef]
- Audira, G.; Sampurna, B.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A Versatile Setup for Measuring Multiple Behavior Endpoints in Zebrafish. Inventions 2018, 3, 75. [Google Scholar] [CrossRef]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Han, L.-W.; Hsiao, C.-D. Establishing simple image-based methods and a cost-effective instrument for toxicity assessment on circadian rhythm dysregulation in fish. Boil. Open 2019, 8, bio041871. [Google Scholar] [CrossRef]
- Thi, N.B.; Anh, N.T.N.; Audira, G.; Juniardi, S.; Liman, R.; Villaflores, O.; Lai, Y.-H.; Chen, J.-R.; Liang, S.-T.; Huang, J.-C.; et al. Development of a Modified Three-Day T-maze Protocol for Evaluating Learning and Memory Capacity of Adult Zebrafish. Int. J. Mol. Sci. 2020, 21, 1464. [Google Scholar] [CrossRef]
- Carvalho, C.; Fernandes, M. Effect of temperature on copper toxicity and hematological responses in the neotropical fish Prochilodus scrofa at low and high pH. Aquaculture 2006, 251, 109–117. [Google Scholar] [CrossRef]
- Takasusuki, J.; Araujo, M.R.R.; Fernandes, M.N. Effect of water pH on copper toxicity in the neotropical fish, Prochilodus scrofa (Prochilodondidae). Bull. Environ. Contam. Toxicol. 2004, 72, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimpour, M.; Alipour, H.; Rakhshah, S. Influence of water hardness on acute toxicity of copper and zinc on fish. Toxicol. Ind. Heal. 2010, 26, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Simonato, J.D.; Mela, M.; Doria, H.B.; Guiloski, I.C.; Randi, M.A.F.; Carvalho, P.S.M.; Meletti, P.C.; De Assis, H.C.S.; Bianchini, A.; Martinez, C.B. Biomarkers of waterborne copper exposure in the Neotropical fish Prochilodus lineatus. Aquat. Toxicol. 2016, 170, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, F.G.; Boijink, C.; Dos Santos, L.R.B.; Oba, E.T.; Kalinin, A.L.; Rantin, F. The combined effect of copper and low pH on antioxidant defenses and biochemical parameters in neotropical fish pacu, Piaractus mesopotamicus (Holmberg, 1887). Ecotoxicology 2010, 19, 963–976. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, F.G.; Boijink, C.; Dos Santos, L.R.B.; Oba, E.T.; Kalinin, A.L.; Luiz, A.J.B.; Rantin, F.T. Antioxidant defenses and biochemical changes in the neotropical fish pacu, Piaractus mesopotamicus: Responses to single and combined copper and hypercarbia exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 156, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Tamilvanan, A.; Balamurugan, K.; Ponappa, K.; Kumar, B.M. Copper Nanoparticles: Synthetic Strategies, Properties and Multifunctional Application. Int. J. Nanosci. 2014, 13, 1430001. [Google Scholar] [CrossRef]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Gracia-Pinilla, M.A.; Martínez, E.; Vidaurri, G.S.; Pérez-Tijerina, E. Deposition of Size-Selected Cu Nanoparticles by Inert Gas Condensation. Nanoscale Res. Lett. 2009, 5, 180–188. [Google Scholar] [CrossRef]
- Cioffi, N.; DiTaranto, N.; Torsi, L.; Picca, R.A.; De Giglio, E.; Sabbatini, L.; Novello, L.; Tantillo, G.M.; Bleve-Zacheo, T.; Zambonin, P.G. Synthesis, analytical characterization and bioactivity of Ag and Cu nanoparticles embedded in poly-vinyl-methyl-ketone films. Anal. Bioanal. Chem. 2005, 382, 1912–1918. [Google Scholar] [CrossRef]
- Mishra, G.; Verma, S.K.; Singh, D.; Yadawa, P.K.; Yadav, R.R. Synthesis and Ultrasonic Characterization of Cu/PVP Nanoparticles-Polymer Suspensions. Open J. Acoust. 2011, 1, 9–14. [Google Scholar] [CrossRef]
- Pham, L.Q.; Shon, J.-H.; Kim, C.W.; Park, J.H.; Kang, H.S.; Lee, B.C.; Kang, Y.S. Copper nanoparticles incorporated with conducting polymer: Effects of copper concentration and surfactants on the stability and conductivity. J. Colloid Interface Sci. 2012, 365, 103–109. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.; Pang, J.; Lin, J. Fabrication and performance of carbon coated copper nanoparticles. Mater. Sci. Pol. 2010, 28, 181–187. [Google Scholar]
- Kobayashi, Y.; Ishida, S.; Ihara, K.; Yasuda, Y.; Morita, T.; Yamada, S. Synthesis of metallic copper nanoparticles coated with polypyrrole. Colloid Polym. Sci. 2009, 287, 877–880. [Google Scholar] [CrossRef]
- Wang, S.; Liu, Y.; Miao, H.; Tan, G.; Zhao, F.; Gao, Y.; Luo, H. Synthesizing and stabilizing copper nanoparticles by coating with a silica layer in aqueous solution. J. Ceram. Soc. Jpn. 2012, 120, 248–250. [Google Scholar] [CrossRef][Green Version]
- Xuan, Y.; Li, Q. Heat transfer enhancement of nanofluids. Int. J. Heat Fluid Flow 2000, 21, 58–64. [Google Scholar] [CrossRef]
- Kantam, M.L.; Jaya, V.S.; Lakshmi, M.J.; Reddy, B.R.; Choudary, B.; Bhargava, S.K. Alumina supported copper nanoparticles for aziridination and cyclopropanation reactions. Catal. Commun. 2007, 8, 1963–1968. [Google Scholar] [CrossRef]
- Karthik, A.D.; Geetha, K. Synthesis of copper precursor, copper and its oxide nanoparticles by green chemical reduction method and its antimicrobial activity. J. Appl. Pharm. Sci. 2013, 3, 16–21. [Google Scholar]
- Ruparelia, J.; Chatterjee, A.K.; Duttagupta, S.P.; Mukherji, S. Strain specificity in antimicrobial activity of silver and copper nanoparticles. Acta Biomater. 2008, 4, 707–716. [Google Scholar] [CrossRef]
- Huang, T.-K.; Lin, K.-W.; Tung, S.-P.; Cheng, T.-M.; Chang, I.-C.; Hsieh, Y.-Z.; Lee, C.-Y.; Chiu, H.-T. Glucose sensing by electrochemically grown copper nanobelt electrode. J. Electroanal. Chem. 2009, 636, 123–127. [Google Scholar] [CrossRef]
- Majzlik, P.; Strasky, A.; Adam, V.; Nemec, M.; Trnkova, L.; Zehnalek, J.; Hubalek, J.; Provaznik, I.; Kizek, R. Influence of zinc (II) and copper (II) ions on Streptomyces bacteria revealed by electrochemistry. Int. J. Electrochem. Sci. 2011, 6, 2171–2191. [Google Scholar]
- Ramyadevi, J.; Jeyasubramanian, K.; Marikani, A.; Rajakumar, G.; Rahuman, A.A. Synthesis and antimicrobial activity of copper nanoparticles. Mater. Lett. 2012, 71, 114–116. [Google Scholar] [CrossRef]
- Wei, Y.; Chen, S.; Kowalczyk, B.; Huda, S.; Gray, T.P.; Grzybowski, B.A. Synthesis of Stable, Low-Dispersity Copper Nanoparticles and Nanorods and Their Antifungal and Catalytic Properties. J. Phys. Chem. C 2010, 114, 15612–15616. [Google Scholar] [CrossRef]
- Griffitt, R.J.; Weil, R.; Hyndman, K.A.; Denslow, N.D.; Powers, K.; Taylor, D.; Barber, D.S. Exposure to Copper Nanoparticles Causes Gill Injury and Acute Lethality in Zebrafish (Danio rerio). Environ. Sci. Technol. 2007, 41, 8178–8186. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-X.; Zhang, T.; Sun, H.-J.; Liu, J.-X. Copper nanoparticles induce zebrafish intestinal defects via endoplasmic reticulum and oxidative stress. Metallomics 2020, 12, 12–22. [Google Scholar] [CrossRef]
- Brun, N.; Varela, M.; Peijnenburg, W.J.G.M.; Spaink, H.P.; Koch, B.E.V.; Vijver, M.G. Nanoparticles induce dermal and intestinal innate immune system responses in zebrafish embryos. Environ. Sci. Nano 2018, 5, 904–916. [Google Scholar] [CrossRef]
- Thit, A.; Huggins, K.; Selck, H.; Baun, A. Acute toxicity of copper oxide nanoparticles to Daphnia magna under different test conditions. Toxicol. Environ. Chem. 2017, 99, 665–679. [Google Scholar] [CrossRef]
- Shaw, B.J.; Al-Bairuty, G.; Handy, R.D. Effects of waterborne copper nanoparticles and copper sulphate on rainbow trout, (Oncorhynchus mykiss): Physiology and accumulation. Aquat. Toxicol. 2012, 116, 90–101. [Google Scholar] [CrossRef]
- Wang, T.; Long, X.H.; Cheng, Y.; Liu, Z.; Yan, S. The potential toxicity of copper nanoparticles and copper sulphate on juvenile Epinephelus coioides. Aquat. Toxicol. 2014, 152, 96–104. [Google Scholar] [CrossRef]
- Tesser, M.E.; De Paula, A.A.; Risso, W.E.; Monteiro, R.A.; Pereira, A.D.E.S.; Fraceto, L.F.; Martinez, C.B.D.R. Sublethal effects of waterborne copper and copper nanoparticles on the freshwater Neotropical teleost Prochilodus lineatus: A comparative approach. Sci. Total. Environ. 2020, 704, 135332. [Google Scholar] [CrossRef]
- Black, M.N.; Henry, E.F.; Adams, O.A.; Bennett, J.C.F.; MacCormack, T. Environmentally relevant concentrations of amine-functionalized copper nanoparticles exhibit different mechanisms of bioactivity inFundulus Heteroclitusin fresh and brackish water. Nanotoxicology 2017, 11, 1070–1085. [Google Scholar] [CrossRef]
- Abdollahzadeh, F.; Khayatzadeh, J.; Ghasemzadeh, F. Comparing study of the effects of copper nanoparticles and copper sulphate on gill histopathology and growth rate of grass carp (Ctenopharyngodon idella). Iran. Sci. Fish. J. 2018, 27, 81–90. [Google Scholar]
- Aghamirkarimi, S.; Mashinchian, M.A.; Sharifpour, I.; Jamili, S.; Ghavam, M.P. Sublethal Effects of Copper Nanoparticles on the Histology of Gill, Liver and Kidney of the Caspian Roach, Rutilus Rutilus Caspicus (Case Study). Glob. J. Environ. Sci. Manag. 2017. [Google Scholar] [CrossRef]
- Sahraei, H.; Hoseini, S.A.; Hedayati, S.A.; Ghorbani, R. Gill Histopathological Changes of Common Carp (Cyprinus carpio) During Exposure to Sub-Lethal Concentrations of Copper Oxide Nanoparticles. J. Environ. Sci. Technol. 2018, 2, 93–103. [Google Scholar]
- Noureen, A.; Jabeen, F.; A Tabish, T.; Ali, M.; Iqbal, R.; Yaqub, S.; Chaudhry, A.S. Histopathological changes and antioxidant responses in common carp (Cyprinus carpio) exposed to copper nanoparticles. Drug Chem. Toxicol. 2019, 1–8. [Google Scholar] [CrossRef]
- Mota, S.B.; Campos, D.; MacCormack, T.; Duarte, R.; Val, A.L.; Almeida-Val, V.M.F. Mechanisms of toxic action of copper and copper nanoparticles in two Amazon fish species: Dwarf cichlid (Apistogramma agassizii) and cardinal tetra (Paracheirodon axelrodi). Sci. Total. Environ. 2018, 630, 1168–1180. [Google Scholar] [CrossRef]
- Al Ghais, S.; Bhardwaj, V.; Kumbhar, P.; Al Shehhi, O. Effect of copper nanoparticles and organometallic compounds (dibutyltin) on tilapia fish. J. Basic Appl. Zool. 2019, 80, 32. [Google Scholar] [CrossRef]
- Aghamirkarimi, S.; Mashinchian Moradi, A.; Sharifpour, I.; Jamili, S.; Ghavam Mostafavi, P. Effect of copper nanoparticles in the Caspian Roach (Rutillus rutillus caspicus), changing antioxidant activities and liver histopathology. Iran. Sci. Fish. J. 2019, 27, 125–134. [Google Scholar]
- Gupta, Y.R.; Sellegounder, D.; Kannan, M.; Deepa, S.; Senthilkumaran, B.; Basavaraju, Y. Effect of copper nanoparticles exposure in the physiology of the common carp (Cyprinus carpio): Biochemical, histological and proteomic approaches. Aquac. Fish. 2016, 1, 15–23. [Google Scholar] [CrossRef]
- Wang, T.; Wen, X.; Hu, Y.; Zhang, X.; Wang, D.; Yin, S. Copper nanoparticles induced oxidation stress, cell apoptosis and immune response in the liver of juvenile Takifugu fasciatus. Fish Shellfish. Immunol. 2019, 84, 648–655. [Google Scholar] [CrossRef]
- Tunçsoy, M.; Erdem, C. Copper Accumulation in Tissues of Oreochromis niloticus Exposed to Copper Oxide Nanoparticles and Copper Sulphate with Their Effect on Antioxidant Enzyme Activities in Liver. Water Air Soil Pollut. 2018, 229, 269. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Hedayati, A.; Mirghaed, A.T.; Ghelichpour, M. Toxic effects of copper sulfate and copper nanoparticles on minerals, enzymes, thyroid hormones and protein fractions of plasma and histopathology in common carp Cyprinus carpio. Exp. Toxicol. Pathol. 2016, 68, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Vajargah, M.F.; Yalsuyi, A.M.; Sattari, M.; Prokić, M.D.; Faggio, C. Effects of Copper Oxide Nanoparticles (CuO-NPs) on Parturition Time, Survival Rate and Reproductive Success of Guppy Fish, Poecilia reticulata. J. Clust. Sci. 2019, 31, 499–506. [Google Scholar] [CrossRef]
- Mansano, A.S.; Souza, J.P.; Cancino-Bernardi, J.; Venturini, F.P.; Marangoni, V.S.; Zucolotto, V. Toxicity of copper oxide nanoparticles to Neotropical species Ceriodaphnia silvestrii and Hyphessobrycon eques. Environ. Pollut. 2018, 243, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Murugananthkumar, R.; Rajesh, D.; Senthilkumaran, B. Copper Nanoparticles Differentially Target Testis of the Catfish, Clarias batrachus: In vivo and In vitro Study. Front. Environ. Sci. 2016, 4, 43. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, Q.; Li, X.; Zhan, S.; Wang, L.; Chen, N. The effects of copper oxide nanoparticles on dorsoventral patterning, convergent extension, and neural and cardiac development of zebrafish. Aquat. Toxicol. 2017, 188, 130–137. [Google Scholar] [CrossRef]
- McNeil, P.L.; Boyle, D.; Henry, T.B.; Handy, R.D.; Sloman, K.A. Effects of metal nanoparticles on the lateral line system and behaviour in early life stages of zebrafish (Danio rerio). Aquat. Toxicol. 2014, 152, 318–323. [Google Scholar] [CrossRef]
- Sovová, T.; Boyle, D.; Sloman, K.A.; Pérez, C.V.; Handy, R.D. Impaired behavioural response to alarm substance in rainbow trout exposed to copper nanoparticles. Aquat. Toxicol. 2014, 152, 195–204. [Google Scholar] [CrossRef]
- Bai, W.; Tian, W.; Zhang, Z.; He, X.; Ma, Y.; Liu, N.; Chai, Z. Effects of copper nanoparticles on the development of zebrafish embryos. J. Nanosci. Nanotechnol. 2010, 10, 8670–8676. [Google Scholar] [CrossRef]
- Torres-Duarte, C.; Adeleye, A.S.; Pokhrel, S.; Mädler, L.; Keller, A.A.; Cherr, G.N. Developmental effects of two different copper oxide nanomaterials in sea urchin ( Lytechinus pictus ) embryos. Nanotoxicology 2015, 10, 1–9. [Google Scholar] [CrossRef]
- Ates, M.; Dugo, M.; Demir, V.; Arslan, Z.; Tchounwou, P. Effect of Copper Oxide Nanoparticles to Sheepshead Minnow (Cyprinodon Variegatus) at Different Salinities. Dig J Nanomater. Bios 2014, 9, 369–377. [Google Scholar]
- Quik, J. Fate of nanoparticles in the aquatic environment. Fate of nanoparticles in the aquatic environment. Removal of engineered nanomaterials from the water phase under environmental conditions. Ph.D. Thesis, Radboud University Nijmegen, Nijmegen, The Netherlands, 2013. [Google Scholar]
- Song, L.; Vijver, M.G.; Peijnenburg, W.J.G.M.; Galloway, T.S.; Tyler, C.R. A comparative analysis on the in vivo toxicity of copper nanoparticles in three species of freshwater fish. Chemosphere 2015, 139, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Khabbazi, M.; Harsij, M.; Hedayati, S.A.A.; Gholipoor, H.; Gerami, M.H.; Ghafari Farsani, H. Effect of CuO nanoparticles on some hematological indices of rainbow trout oncorhynchus mykiss and their potential toxicity. Nanomed. J. 2015, 2, 67–73. [Google Scholar]
- Lin, S.; Taylor, A.A.; Ji, Z.; Chang, C.H.; Kinsinger, N.M.; Ueng, W.; Walker, S.L.; Nel, A.E.; Zhaoxia, J. Understanding the Transformation, Speciation, and Hazard Potential of Copper Particles in a Model Septic Tank System Using Zebrafish to Monitor the Effluent. ACS Nano 2015, 9, 2038–2048. [Google Scholar] [CrossRef] [PubMed]


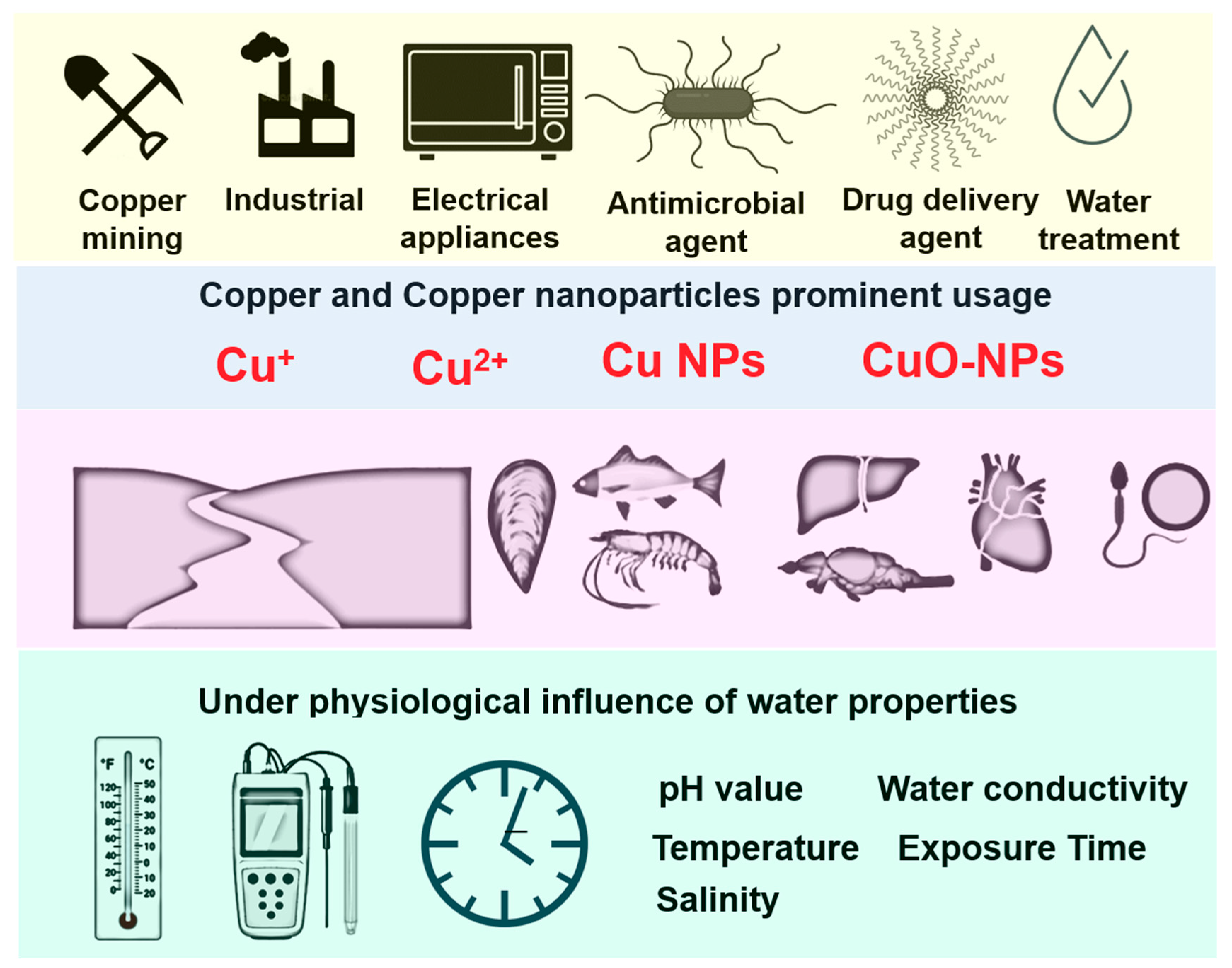{kind=link}
| Organs | Model Organism | Toxicity Response | Copper Concentration | Exposure Time | References |
|---|---|---|---|---|---|
| Gills | Oreochromis niloticus | Inhibition of Na+/K+-ATPase activity | Cu2+, 40 and 400 µg/L | 0, 3, 7, 14, and 21 days | [83] |
| Oncorhynchus mykiss | Inhibition of Na+/K+-ATPase activity (LC50 10 µM) | 1 mM | 30 min | [88] | |
| Oncorhynchusmykiss Perca flavescens | Different mechanisms of copper tolerance or copper burden do not necessarily translate into toxicity. LC50 Hard Water 1.05 vs. 4.16 mM. LC50 Soft Water 0.10 vs. 0.44 mM | CuSO4. Hard water: 0.38 to 15.35 µM. Soft water: 0.11 to 0.77 µM | 3 h | [92] | |
| Oncorhynchus mykiss | Decline of plasma Na+ and Cl− and arterial O2 tension, cell swelling, thickening, and curling of the gill lamellae | 4.9 µmol/L | 24 h | [114] | |
| Oncorhynchus mykiss | Hyperplasia, aneurisms, and necrosis in secondary lamellae of the gills | CuSO4, 20 or 100 µg/L | 0, 4, and 10 days | [96] | |
| Opsanus beta | No inhibition of Na+/K+-ATPase | Copper, 12.8 ± 1.6 µM, 55.2 ± 5.0 µM | 30 days | [89] | |
| Liver | Rutilus rutilus | Combined effect of nutrition and copper in toxicity | 80 µg Cu/L 0, 25, 100, and 200 µg/L, and 20 or 100 µg/L | 7 days | [94] |
| Brachydanio rerio | Copper accumulation. Large lysed area, hepatocyte alterations, and increase in antioxidative defense | CuSO4, 40 ± 5 or 140 ± 30 µg Cu/L | 14 days | [95] | |
| Oncorhynchusmykiss | Hepatitis injuries and cells with pyknotic nuclei in the liver | CuSO4, 20 or 100 µg/L | 0, 4, and 10 days | [96] | |
| Prochilodun lineatus | Activity of key liver enzymes change with ambient water surroundings. Copper accumulation in the liver was high in fish at 20 °C and pH 8.0 | 98 ± 0.8, 16 ± 0.2, 88 ± 0.8, 14 ± 0.5 µg Cu/L (pH 7.0, 4.5, 8.0). Temperatures of 20 and 30 °C | 96 h | [97] | |
| Catla catla | Cytolysis, necrosis, pyknosis, and fibrosis at 24 h. LC50 0.75 mg/L | 0.1 and 0.3 mg/L | 3 weeks | [98] | |
| Gasterosteus aculeatus | Copper accumulation at day 8 and 12, induced oxidative stress | 0, 25, 100, and 200 µg/L | 4, 8, 12, and 21 days | [99] | |
| Dicentrarchus labrax | Metallothionine reduction with copper injection, depicting toxicity | 500 ng/g | 24 h | [93] | |
| Kidney | Oncorhynchusmykiss | Damage to epithelium of some renal tubules and increased Bowman’s space | CuSO4, 20 or 100 µg/L | 0, 4, and 10 days | [96] |
| Dicentrarchus labrax | Damage to pronephros and lysosomal membranes. Oxi-radical generation. Copper depicted a high binding affinity with the head kidney of common carp. LC50 2500 ng/g | Cu injection, 50, 250, and 1000 ng/g | 48 h | [100] | |
| Cyprinus carpio L | Buffering capacity of copper was high in fish. Increase in number of blast cells, proliferating cell nuclear antigen (PCNA), and apoptotic cells. Similar effects for short term and long term exposure | Short term: 0.75 mg/dm3 Cu. Long term: 0.075 mg/dm3 Cu | Short term: 3 h. Long term: 4 weeks | [101,102] | |
| Reproductive organs | Carassius auratus and Xiphophorus helleri | Copper accumulation, effect on growth rate, and reproductive performance. Reduction in mean diameter and weight of eggs. 96 h LC50 0.30 and 0.36 ppm | 0, 0.15, 0.20, 0.25, 0.30, 0.35, and 0.40 mg/dm3. 0, 0.18, 0.24, 0.30, 0.36, 0.42, 0.48, and 0.54 mg.dm3 | 96 h | [103] |
| Sparus sarba | Accumulation was dose-dependent with similar accumulations in fingerlings and subadults; 24 h, 48 h, 72 h, and 96 h LC50 of 2.01, 1.28, 1.17, 1.03 mg/L for fingerlings and 2.36, 1.52, 1.34, and 1.24 mg/L for subadults, respectively | 0.15, 0.30, and 0.45 mg Cu/L | 30 days | [104] | |
| Cyprinus carpio, Silurus aristotelis, Rutilus ylikiensis, and Carassius gibelio | C. carpio and R. ylikiensis presented the highest metal content in gonads | Fish from Lake Pamvotis | - | [105] | |
| Puntius conchonius | Copper interfered with spermatogenesis temporarily, induced atresia in ovaries; 96 h Median Tolerance Limit (TLm) 0.571 ± 0.020 mg/L | Lake Nainital | 2, 3, 4 months | [106] | |
| Cyprinus carpio, Carassius auratus gibelio, Rutilus rutilus heckeli, Abramis brama, Aristichthys nobilis, Hypophtalmichthys molitrix, and Sander lucioperca | Accumulation pattern was similar in the investigated species and increased greatly in the prespawning period | Freshwater ecosystem of Moldova | - | [109] | |
| Gasterosteus aculeatus | Copper increased T4 and T3 level in ovaries | 100 µg/L | 2 h, 1 week | [110] | |
| Danio rerio | Hatching inhibition and slow development | 50–1000 µg Cu/L, 68.35 ± 4.27 and 244.36 ± 17.40 g Cu/L | 48, 72, and 96 hpf | [115] | |
| Lepomis macrochirus | Survival was reduced, growth was retarded, and spawning was inhibited; 96 h TL50 1100 µg Cu/L. Maximum Acceptable Toxicant Concentration (MATC)/96 h TL50 0.02 and 0.04 | 40–162 µg Cu/L | 22 month | [125] | |
| Heart | Cyprinus carpio L | Increase in acetylcholine esterase (AChE) | 1.0, 10.0, or 50.0 ppm | 2 h | [112] |
| Oncorhynchusmykiss | Heart rate decreased, LC50 1.33 to 0.06 mg Cu/L. Measured Toxicity Equivalency Concentration (TEC) 0.09, 0.04, 0.02 mg/L, copper. No Observed Effect Concentration (NOEC) 0.06, 0.03, 0.015 mg/L, respectively | 0.008 mg/L to 8.0 mg/L | 96 h and 65 days | [113] | |
| Oncorhynchusmykiss | Doubling of mean arterial blood pressure and cardiac failure | 4.9 µmol copper/L | 24 h | [114] | |
| Daniorerio | Fastest heart rates at 28 hpf for stress response | 50–1000 µg Cu/L, 68.35 ± 4.27 and 244.36 ± 17.40 g Cu/L | 24, 48, 72, and 96 hpf | [115] | |
| Brain | Cyprinus carpio | Decrease in brain 5-HT and dopamine levels, affecting behavior and locomotor control | 0.22, 0.34, and 0.84 µM | 1 week | [116] |
| Lagodon rhomboids, Micropogon undulates, Archosargus probatocephalus, and Arius felis | Behavioral variables, general activity, swimming speeds, and angular orientation of movements affected. Locomotor activity and angular orientation of movements | 0.1 mg/L | 72 h | [117] | |
| Arius felis | Here, 0.005, 0.01, and 0.05 mg, Cu2+/L hypoactive, 0.1 and 0.2 mg Cu2+/L elicited hyperactivity and an increase in orientation angle of movement | 0.0, 0.005, 0.01, 0.05, 0.1, or 0.2 mg Cu2+/L | 72 h | [118] | |
| Saluelinus jontinalis | Increase locomotor activity and decrease in feeding response | 6–60 µg/L | 2 h | [121] | |
| Menidia menidia | At low copper concentrations, an increased swimming speed, decreased rate of change of direction and their nearest-neighbor distances. Swimming in a parallel orientation were observed; 96 h LC50 136 µg/L Cu | 0–100 µg Cu/L | 4 days | [119] | |
| Salmo gairdneri | Increased swimming speed | 0–200 µg Cu/L | 96 h–30 days | [120] | |
| Lepomis macmchirus | Decreased prey consumption and increased handling times of preying on five different invertebrate prey | 31, 1 80, 1 710 µg/L | 1 week | [122] | |
| Pimephales promelas | Copper may disrupt important developmental stages in the embryonic olfactory system, also chemosensory impairment | 0 or 10 μg Cu/L | 5–7 days | [123] | |
| Carassius auratus | Frequency of entry and time spent increased at a temperature of 21.5 ± 0.1 °C | 0.010 ppm, temperatures of 21.1 ± 0.1 and 21.5 ± 0.1 °C | 24 h | [124] |
| Organs Affected | Organism | CuNPs Tested | Adverse Outcome | Time | Characteristic | References |
|---|---|---|---|---|---|---|
| Gills | Danio rerio | CuNPs | Toxicity in gills, not adequately explained by dissolution of particles alone; 48 h LC50 1.5 mg/L | 48 h | 80 nm | [154] |
| Oncorhynchus mykiss | CuNPs | Reduction in branchial Na+/K+-ATPase, increase in Thiobarbituric acid reactive substances (TBARS) | 0, 4, 10 days | 87 ± 27 nm, 20 and 100 µg/L | [158] | |
| Fundulus heteroclitus | CuNPs | >40% reduction in Na+/K+-ATPase in FW | 48 h | 5–10 nm, 10 µg/L | [161] | |
| Ctenopharyngodon idella | CuO-NPs | Growth reduction | 30 days | 50 nm, 100 and 200 mg/L | [162] | |
| Cyprinus carpio | CuO-NPs | Cell swelling, aourtism, edema, hyperplasia | 6 weeks | 10, 40, 80 mg/L | [164] | |
| Apistogramma agassizii | CuO-NPs | Oxidative stress | 96 h | HD 50 nm, 58.31, 69.6 µg/L | [166] | |
| Plotosus lineatus | CuNPs | Increase in H+ and Na+/K+ pump activity | 96 h | 40–60 nm, 12.16 ± 1.77 µg/L | [160] | |
| Rutilus rutilus caspicus | CuNPs | Fish became anemic | 21 days | 40 nm, 0.1, 0.2, and 0.5 mg/L | [163] | |
| Ceriodaphnia silvestrii and Hyphessobrycon eques | CuO-NPs | Fusion of secondary lamellae, cellular hyperplasia. Increase in reactive oxygen species (ROS). Apoptosis and necrosis | Acute-24 h Chronic-8 days | HD 30 nm, acute, 0, 0.7, 10.0, 13.0, 16.0, 19.0 µg/L, chronic, 0, 0.5, 1.0, 2.0, 4.0, 8.0, 10.0 µg/L | [174] | |
| Liver | Rutilus rutilus caspicus | CuNPs | Deformation of nuclei, cytoplasmic vacuolation, cellular degeneration, congestion in the blood sinusoids, and necrosis of the hepatocytes | 21 days | 40 nm, 0.1, 0.2, and 0.5 mg/L | [163] |
| Rutilus rutilus caspicus | CuNPs | Blood congestion in central veins and necrosis of hepatocytes. Pyknotic nuclei | 21 days | 0.1, 0.2, and 0.5 mg/L | [168] | |
| Cyprinus carpio | CuNPs | Increase in oxidative stress markers MDA, T-SOD, GSH, CAT, and caspases | 7 days | <50 nm, 20, and 100 µg/L | [169] | |
| Trachidermus fasciatus | CuNPs | Increase in MDA, T-SOD, CAT, GSH, Casp-3, 9, decrease in Na+/K+-ATPase, and cyt c | 30 days | 10–30 nm, 20 and 100 µg/L | [170] | |
| Oreochromis niloticus | CuO-NPs | Increase in SOD, CAT, GPX | 1, 7 and 15 days | <50 nm, 10, 50 and 100 µg/L | [171] | |
| Kidney | Rutilus rutilus caspicus | CuNPs | Degeneration in the tubule cells, interstitial tissue, glomerular shrinkage, increase in interstitial tissue cells, and macrophages aggregation | 21 days | 40 nm, 0.1, 0.2, 0.5 mg/L | [163] |
| Oreochromis niloticus | CuO-NPs | Excess copper bound to metallothionine (MT) | 1, 7 and 15 days | <50 nm, 10, 50, and 100 µg Cu/L | [171] | |
| Cyprinus carpio | CuNPs | Tubular vacuolization and necrosis, melano–macrophage center, Bowman’s capsule edema, glomerulus degeneration, and hyperemia | 14 days | 40 nm, 0.25 and 25 mg/L | [172] | |
| Reproductive Organs | Poecilia reticulata | CuO-NPs | Lowered reproductive stress, prolonged parturition time, and increased mortality rate | 96 h | PS, 40 nm 0.5–45 mg/L, 0.5–10 mg/L | [173] |
| Ceriodaphnia silvestrii and Hyphessobrycon eques | CuO-NPs | Decrease in reproduction, feeding inhibition, increased reactive oxygen species (ROS) | 0, 24 and 48 h | HD, 30 nm, acute, 0, 0.7, 10.0, 13.0, 16.0, 19.0 µg/L, chronic, 0, 0.5, 1.0, 2.0, 4.0, 8.0, 10.0 µg/L | [174] | |
| Clarias batrachus | CuNPs | Testis-related genes showed upregulation, increase in level of androgens, and disruption in structural analysis | 21 days | <50 nm, 100 µg/L | [175] | |
| Danio rerio | CuNPs | Retarded hatching embryos, causing morphological malformation and mortality in the gastrula stage | 96 h | 69 ± 18 nm, 0.01, 0.05, 0.1, 0.5, 1 mg/L | [179] | |
| Leiarius pictus | CuO-NPs | The developmental abnormalities caused by dissolution of internalized CuO-NPs | 96 hpf | ~10 nm, 20–2000 ppb commercial nano-CuO, 380–38,000 ppb synthesized nano-CuO | [180] | |
| Heart | Danio rerio | CuO-NPs | Prevention of looping of the heart tube during cardiogenesis, also disturbing the dorsoventral patterning, increasing wnt8 and vent expression | 120 hpf | 100–500 nm, 0.1, 0.5, 5, and 50 mg/L | [176] |
| Brain | Tilapia mossambica | CuNPs | Reduction in oxidative stress enzymes glutathione (GSH), glutathione-s-transferase (GST), and acetylcholine esterase (AChE) | 6 days | 15 mg/L | [167] |
| Danio rerio | CuNPs | Reduced lateral line neuromasts (LLN) and performance of rheotaxis | 96 h | 20 ± 9 nm, 50 and 225 µg/L | [177] | |
| Oncorhynchus mykiss | CuNPs | Oxidative stress—increment in the ratio of oxidized to reduced glutathione | 12 h | PS, <50 nm, 50 µg/L | [178] | |
| Cyprinodon variegatus | CuO-NPs | Behavioral changes, increase in mucus secretion, and loss in equilibrium | 7 days | PS, 40 nm, 5 and 50 mg/L | [181] |
| Tissues | Copper Ions | Copper Nanoparticles |
|---|---|---|
| Brain | Disruption in behavioral, olfactory, and chemosensory impairments | Increase in oxidative stress, behavioural impairments |
| Liver | Liver is the major site for copper accumulation Cytolysis, necrosis, pyknosis, fibrosis, and induced oxidative stress | Increase in oxidative stress markers MDA, T-SOD, GSH, CAT & Caspase Necrosis, and pyknotic nuclei |
| Heart | Increase in AChE Abrupt changes in heart beats | Prevention of looping of the heart tube during cardiogenesis |
| Reproduction organs | Copper accumulation, interference with egg hatching and spermatogenesis, Increase in T3&T4 in ovary | Developmental abnormalities Retard hatching embryos Increase in ROS |
| Gill | Inhibition of Na+/K+ ATPase activity | Reduction in branchial Na+/K+ ATPase, Increased oxidative stress, Inhibition of AChE |
| Kidney | Damage to renal tubules, pronephros, high buffering capacity of copper | Tubular vacuolization and necrosis Glomerular shrinkage Macrophages aggregation |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malhotra, N.; Ger, T.-R.; Uapipatanakul, B.; Huang, J.-C.; Chen, K.H.-C.; Hsiao, C.-D. Review of Copper and Copper Nanoparticle Toxicity in Fish. Nanomaterials 2020, 10, 1126. https://doi.org/10.3390/nano10061126
Malhotra N, Ger T-R, Uapipatanakul B, Huang J-C, Chen KH-C, Hsiao C-D. Review of Copper and Copper Nanoparticle Toxicity in Fish. Nanomaterials. 2020; 10(6):1126. https://doi.org/10.3390/nano10061126
Chicago/Turabian StyleMalhotra, Nemi, Tzong-Rong Ger, Boontida Uapipatanakul, Jong-Chin Huang, Kelvin H.-C. Chen, and Chung-Der Hsiao. 2020. "Review of Copper and Copper Nanoparticle Toxicity in Fish" Nanomaterials 10, no. 6: 1126. https://doi.org/10.3390/nano10061126
APA StyleMalhotra, N., Ger, T.-R., Uapipatanakul, B., Huang, J.-C., Chen, K. H.-C., & Hsiao, C.-D. (2020). Review of Copper and Copper Nanoparticle Toxicity in Fish. Nanomaterials, 10(6), 1126. https://doi.org/10.3390/nano10061126