Cognitive Archeology and the Attentional System: An Evolutionary Mismatch for the Genus Homo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Attention and Human Evolution
2. Cognitive Archeology and Attention
2.1. Alerting and Orienting
2.2. Executive Attention
2.3. A General View
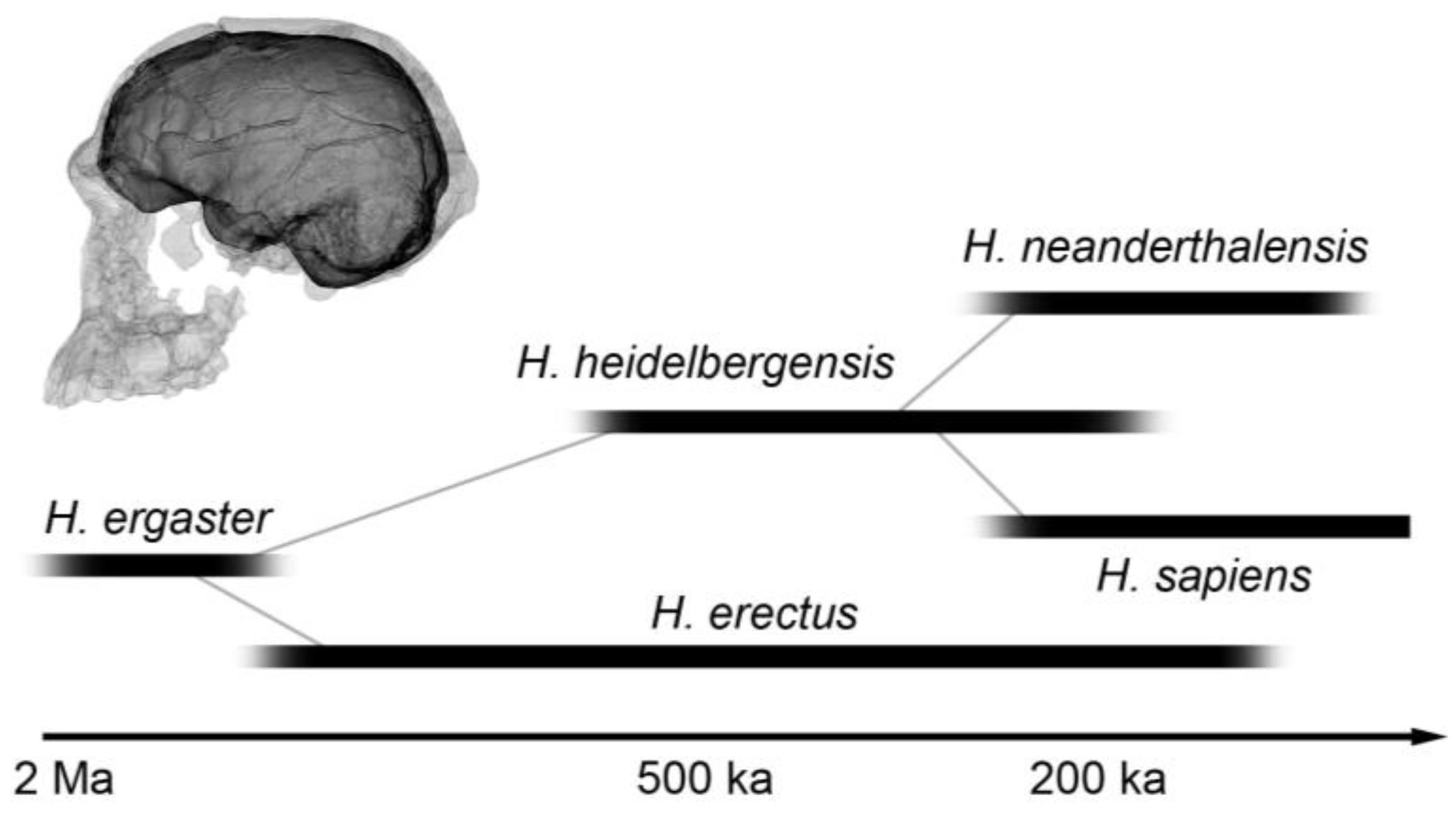
- The relative brain size (encephalization) did increase with the origin of the human genus (H. ergaster and H. erectus), then again during the Middle Pleistocene (H. heidelbergensis), and finally in modern humans (H. sapiens) and Neanderthals (H. neanderthalensis). However, it is important to take into account that this increase deals with average values, while the range of brain size in all these species does largely overlap. Furthermore, considering that correlations between brain size and most cognitive features range from null to modest, mean species differences are not really informative on possible individual differences;
- Technological complexity is similar in H. erectus and H. heildelbergensis. Then, it displays a general increase in H. neanderthalensis and early H. sapiens, and a surprising boost in late modern humans (say in the last 50,000 years). Technological complexity is supposedly linked to distinct cognitive aspects, including reasoning, memory, and attentional skills;
- Similarly, social complexity (group size, social structure, landscape use, economy, hunting strategy, etc.) underwent an apparent increase in H. neanderthalensis and early H. sapiens, and then a pronounced escalation in the latter lineage.
3. Fronto-Parietal Anatomy and the Human Genus
4. The Parietal Lobe, Attention, and the History of the Self
5. Limitations and Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Ambrose, Stanley H. 2001. Paleolithic technology and human evolution. Science 291: 1748–53. [Google Scholar] [CrossRef]
- Antón, Susan C., and J. Josh Snodgrass. 2012. Origins and evolution of genus Homo: New perspectives. Current Anthropology 53: S479–S496. [Google Scholar] [CrossRef]
- Antón, Susan C., Richard Potts, and Leslie C. Aiello. 2014. Evolution of early Homo: An integrated biological perspective. Science 345: 1236828. [Google Scholar] [CrossRef] [PubMed]
- Ardesch, Dirk Jan, Lianne H. Scholtens, Longchuan Li, Todd M. Preuss, James K. Rilling, and Martijn P. van den Heuvel. 2019. Evolutionary expansion of connectivity between multimodal association areas in the human brain compared with chimpanzees. Proceedings of the National Academy of Sciences USA 116: 7101–6. [Google Scholar] [CrossRef] [PubMed]
- Baddeley, Alan. 2000. The episodic buffer: A new component of working memory? Trends in Cognitive Sciences 4: 417–23. [Google Scholar] [CrossRef] [PubMed]
- Baluch, Farhan, and Laurent Itti. 2011. Mechanisms of top-down attention. Trends in Neurosciences 34: 210–24. [Google Scholar] [CrossRef]
- Behrmann, M., J. J. Geng, and S. Shomstein. 2004. Parietal cortex and attention. Current Opinion in Neurobiology 14: 212–17. [Google Scholar] [CrossRef] [PubMed]
- Bisley, James W., and Michael E. Goldberg. 2010. Attention, intention, and priority in the parietal lobe. Annual Review of Neuroscience 33: 1–21. [Google Scholar] [CrossRef] [PubMed]
- Borghi, Anna M., and Felice Cimatti. 2010. Embodied cognition and beyond: Acting and sensing the body. Neuropsychologia 48: 763–73. [Google Scholar] [CrossRef]
- Brewer, Judson A., Patrick D. Worhunsky, Jeremy R. Gray, Yi-Yuan Tang, Jochen Weber, and Hedy Kober. 2011. Meditation experience is associated with differences in default mode network activity and connectivity. Proceedings of the National Academy of Sciences USA 108: 20254–59. [Google Scholar] [CrossRef] [PubMed]
- Bruner, Emiliano. 2004. Geometric morphometrics and paleoneurology: Brain shape evolution in the genus Homo. Journal of Human Evolution 47: 279–303. [Google Scholar] [CrossRef]
- Bruner, Emiliano. 2013. The species concept as a cognitive tool for biological anthropology. American Journal of Primatology 75: 10–15. [Google Scholar] [CrossRef] [PubMed]
- Bruner, Emiliano. 2015. Human Paleoneurology. Cham: Springer. [Google Scholar]
- Bruner, Emiliano. 2018. Human paleoneurology and the evolution of the parietal cortex. Brain Behavior and Evolution 91: 136–47. [Google Scholar] [CrossRef]
- Bruner, Emiliano. 2019. Human paleoneurology: Shaping cortical evolution in fossil hominids. Journal of Comparative Neurology 527: 1753–65. [Google Scholar] [CrossRef]
- Bruner, Emiliano. 2021. Evolving human brains: Paleoneurology and the fate of Middle Pleistocene. Journal of Archeological Method and Theory 28: 76–94. [Google Scholar] [CrossRef]
- Bruner, Emiliano. 2022a. Prehistory, neuroscience, and evolutionary anthropology: A personal journey. Journal of Anthropological Sciences 100: 173–92. [Google Scholar] [PubMed]
- Bruner, Emiliano. 2022b. A network approach to the topological organization of the Brodmann map. Anatomical Record 305: 3504–15. [Google Scholar] [CrossRef] [PubMed]
- Bruner, Emiliano, Alexandra Battaglia-Mayer, and Roberto Caminiti. 2023c. The parietal lobe evolution and the emergence of material culture in the human genus. Brain Structure and Function 228: 145–67. [Google Scholar] [CrossRef] [PubMed]
- Bruner, Emiliano, Ana Sofia Pereira-Pedro, Xu Chen, and James K. Rilling. 2017b. Precuneus proportions and cortical folding: A morphometric evaluation on a racially diverse human sample. Annals of Anatomy 211: 120–28. [Google Scholar] [CrossRef]
- Bruner, Emiliano, and Amélie Beaudet. 2023. The brain of Homo habilis: Three decades of paleoneurology. Journal of Human Evolution 174: 103281. [Google Scholar] [CrossRef]
- Bruner, Emiliano, and Atsushi Iriki. 2016. Extending mind, visuospatial integration, and the evolution of the parietal lobes in the human genus. Quaternary International 405: 98–110. [Google Scholar] [CrossRef]
- Bruner, Emiliano, and Ben T. Gleeson. 2019. Body cognition and self-domestication in human evolution. Frontiers in Psychology 10: 1111. [Google Scholar] [CrossRef]
- Bruner, Emiliano, and Ralph L. Holloway. 2010. Bivariate approach to the widening of the frontal lobes in the genus Homo. Journal of Human Evolution 58: 138–46. [Google Scholar] [CrossRef] [PubMed]
- Bruner, Emiliano, and Roberto Colom. 2022. Can a Neandertal meditate? An evolutionary view of attention as a core component of general intelligence. Intelligence 93: 101668. [Google Scholar] [CrossRef]
- Bruner, Emiliano, Annapaola Fedato, María Silva-Gago, Rodrigo Alonso-Alcalde, Marcos Terradillos-Bernal, María Ángeles Fernández-Durantes, and Elena Martín-Guerra. 2018a. Visuospatial Integration and Hand-Tool Interaction in Cognitive Archeology. In Processes of Visuospatial Attention and Working Memory. Current Topics in Behavioral Neurosciences 41. Edited by T. Hodgson. Cham: Springer, pp. 13–36. [Google Scholar]
- Bruner, Emiliano, Annapaola Fedato, María Silva-Gago, Rodrigo Alonso-Alcalde, Marcos Terradillos-Bernal, María Ángeles Fernández-Durantes, and Elena Martín-Guerra. 2018b. Cognitive archeology, body cognition, and hand–tool interaction. Progress in Brain Research 238: 325–45. [Google Scholar]
- Bruner, Emiliano, Enza Spinapolice, Ariane Burke, and Karenleigh A. Overmann. 2018c. Visuospatial integration: Paleoanthropological and archeological perspectives. In Evolution of Primates Social Cognition. Edited by L. D. Di Paolo, F. Di Vincenzo and F. De Petrillo. Berlin: Springer, pp. 299–326. [Google Scholar]
- Bruner, Emiliano, José Manuel De la Cuétara, Michael Masters, Hideki Amano, and Naomichi Ogihara. 2014. Functional craniology and brain evolution: From paleontology to biomedicine. Frontiers in Neuroanatomy 8: 19. [Google Scholar] [CrossRef] [PubMed]
- Bruner, Emiliano, María Silva-Gago, Annapaola Fedato, Manuel Martín-Loeches, and Roberto Colom. 2023b. Psychometrics, visuospatial abilities and cognitive archeology. In Cognitive Archeology, Body Cognition, and the Evolution of Visuospatial Perception. Edited by E. Bruner. London: Academic Press, in press. [Google Scholar]
- Bruner, Emiliano, Ralph Holloway, Karen L. Baab, Michael J. Rogers, and Sileshi Semaw. 2023a. The endocast from Dana Aoule North (DAN5/P1): A 1.5 million year-old human braincase from Gona, Afar, Ethiopia. American Journal of Biological Anthropology. [Google Scholar] [CrossRef] [PubMed]
- Bruner, Emiliano, Todd M. Preuss, Xu Chen, and James K. Rilling. 2017a. Evidence for expansion of the precuneus in human evolution. Brain Structure and Function 222: 1053–60. [Google Scholar] [CrossRef]
- Buckner, Randy L., and Fenna M. Krienen. 2013. The evolution of distributed association networks in the human brain. Trends in Cognitive Sciences 17: 648–65. [Google Scholar] [CrossRef]
- Buckner, Randy L., and Lauren M. DiNicola. 2019. The brain’s default network: Updated anatomy, physiology and evolving insights. Nature Reviews Neuroscience 20: 593–608. [Google Scholar] [CrossRef]
- Caramazza, Alfonso, Stefano Anzellotti, Lukas Strnad, and Angelika Lingnau. 2014. Embodied cognition and mirror neurons: A critical assessment. Annual Review of Neuroscience 37: 1–15. [Google Scholar] [CrossRef]
- Carstensen, Laura L., and Marguerite DeLiema. 2018. The positivity effect: A negativity bias in youth fades with age. Current Opinion in Behavioral Sciences 19: 7–12. [Google Scholar] [CrossRef] [PubMed]
- Cavanna, Andrea E., and Michael R. Trimble. 2006. The precuneus: A review of its functional anatomy and behavioural correlates. Brain 129: 564–83. [Google Scholar] [CrossRef] [PubMed]
- Chaieb, Leila, Christian Hoppe, and Juergen Fell. 2022. Mind wandering and depression: A status report. Neuroscience & Biobehavioral Reviews 133: 104505. [Google Scholar]
- Chica, Ana B., Paolo Bartolomeo, and Juan Lupiáñez. 2013. Two cognitive and neural systems for endogenous and exogenous spatial attention. Behavioural Brain Research 237: 107–23. [Google Scholar] [PubMed]
- Christoff, Kalina, Zachary C. Irving, Kieran CR Fox, R. Nathan Spreng, and Jessica R. Andrews-Hanna. 2016. Mind-wandering as spontaneous thought: A dynamic framework. Nature Reviews Neuroscience 17: 718–31. [Google Scholar] [PubMed]
- Cohen, Michael A., Patrick Cavanagh, Marvin M. Chun, and Ken Nakayama. 2012. The attentional requirements of consciousness. Trends in Cognitive Sciences 16: 411–17. [Google Scholar] [CrossRef] [PubMed]
- Coolidge, Frederick L., and Thomas Wynn. 2005. Working memory its executive functions and the emergence of modern thinking. Cambridge Archeological Journal 15: 5–26. [Google Scholar] [CrossRef]
- Coolidge, Frederick L., Karenleigh A. Overmann, and Thomas Wynn. 2011. Recursion: What is it, who has it, and how did it evolve? Wiley Interdisciplinary Reviews: Cognitive Science 2: 547–54. [Google Scholar] [CrossRef]
- Corbetta, M., M. J. Kincade, C. Lewis, A. Z. Snyder, and A. Sapir. 2005. Neural basis and recovery of spatial attention deficits in spatial neglect. Nature Neuroscience 8: 1603–10. [Google Scholar] [CrossRef]
- Corbey, R. 2020. Baldwin effects in early stone tools. Evolutionary Anthropology 29: 237–44. [Google Scholar] [CrossRef]
- Dahl, Cortland J., Antoine Lutz, and Richard J. Davidson. 2015. Reconstructing and deconstructing the self: Cognitive mechanisms in meditation practice. Trends in Cognitive Sciences 19: 515–23. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, Robin IM. 2018. The anatomy of friendship. Trends in Cognitive Sciences 22: 32–51. [Google Scholar] [PubMed]
- Edlow, Brian L., Emi Takahashi, Ona Wu, Thomas Benner, Guangping Dai, Lihong Bu, Patricia Ellen Grant, David M. Greer, Steven M. Greenberg, Hannah C. Kinney, and et al. 2012. Neuroanatomic connectivity of the human ascending arousal system critical to consciousness and its disorders. Journal of Neuropathology & Experimental Neurology 71: 531–46. [Google Scholar]
- Eibl-Eibesfeldt, I. 1989. Human Ethology. New York: Aldine de Gruyter. [Google Scholar]
- Faivre, Nathan, Roy Salomon, and Olaf Blanke. 2015. Visual consciousness and bodily self-consciousness. Current Opinion in Neurology 28: 23–28. [Google Scholar] [CrossRef]
- Federico, Giovanni, Emanuelle Reynaud, Jordan Navarro, Mathieu Lesourd, Vivien Gaujoux, Franck Lamberton, Danièle Ibarrola, Carlo Cavaliere, Vincenzo Alfano, Marco Aiello, and et al. 2022. The cortical thickness of the area PF of the left inferior parietal cortex mediates technical-reasoning skills. Scientific Reports 12: 11840. [Google Scholar] [CrossRef]
- Federico, Giovanni, François Osiurak, Emanuelle Reynaud, and Maria A. Brandimonte. 2021. Semantic congruency effects of prime words on tool visual exploration. Brain and Cognition 15: 105758. [Google Scholar]
- Gibson, James J. 1979. The Ecological Approach to Visual Perception. New York: Psychology Press. [Google Scholar]
- Goldring, A. B., and L. A. Krubitzer. 2017. Evolution of the parietal cortex in mammals: From manipulation to tool use. In Evolution of the Nervous System, 2nd ed. Edited by J. Kaas. Amsterdam: Elsevier, pp. 259–86. [Google Scholar]
- Hagmann, Patric, Leila Cammoun, Xavier Gigandet, Reto Meuli, Christopher J. Honey, Van J. Wedeen, and Olaf Sporns. 2008. Mapping the structural core of human cerebral cortex. PLoS Biology 6: e159. [Google Scholar] [CrossRef]
- Hasenkamp, Wendy, and Lawrence W. Barsalou. 2012. Effects of meditation experience on functional connectivity of distributed brain networks. Frontiers in Human Neuroscience 6: 38. [Google Scholar] [CrossRef] [PubMed]
- Hasenkamp, Wendy, Christine D. Wilson-Mendenhall, Erica Duncan, and Lawrence W. Barsalou. 2012. Mind wandering and attention during focused meditation: A fine-grained temporal analysis of fluctuating cognitive states. Neuroimage 59: 750–60. [Google Scholar] [CrossRef]
- Hecht, Erin E., David A. Gutman, Todd M. Preuss, Mar M. Sanchez, Lisa A. Parr, and James K. Rilling. 2013. Process versus product in social learning: Comparative diffusion tensor imaging of neural systems for action execution–observation matching in macaques, chimpanzees, and humans. Cerebral Cortex 23: 1014–24. [Google Scholar] [CrossRef] [PubMed]
- Hills, Thomas T., Peter M. Todd, David Lazer, A. David Redish, and Iain D. Couzin. 2015. Exploration versus exploitation in space, mind, and society. Trends in Cognitive Sciences 19: 46–54. [Google Scholar] [CrossRef] [PubMed]
- Hommel, Bernhard, Craig S. Chapman, Paul Cisek, Heather F. Neyedli, Joo-Hyun Song, and Timothy N. Welsh. 2019. No one knows what attention is. Attention, Perception, & Psychophysics 81: 2288–303. [Google Scholar]
- Huntenburg, Julia M., Pierre-Louis Bazin, and Daniel S. Margulies. 2017. Large-scale gradients in human cortical organization. Trends in Cognitive Science 22: 21–31. [Google Scholar] [CrossRef] [PubMed]
- Japyassú, Hilton F., and Kevin N. Laland. 2017. Extended spider cognition. Animal Cognition 20: 375–95. [Google Scholar] [CrossRef]
- Kaplan, David Michael. 2012. How to demarcate the boundaries of cognition. Biology and Philosophy 27: 545–70. [Google Scholar] [CrossRef]
- Kastner, Sabine, and Mark A. Pinsk. 2004. Visual attention as a multilevel selection process. Cognitive, Affective, & Behavioral Neuroscience 4: 483–500. [Google Scholar]
- Killingsworth, Matthew A., and Daniel T. Gilbert. 2010. A wandering mind is an unhappy mind. Science 330: 932–32. [Google Scholar] [CrossRef] [PubMed]
- Klein, Raymond M., and Michael A. Lawrence. 2011. The modes and domains of attention. In Cognitive Neuroscience of Attention, 2nd ed. Edited by M. I. Posner. New York: Guilford Press, pp. 11–28. [Google Scholar]
- Krauzlis, Richard J., Anil Bollimunta, Fabrice Arcizet, and Lupeng Wang. 2014. Attention as an effect not a cause. Trends in Cognitive Sciences 18: 457–64. [Google Scholar] [CrossRef]
- Krauzlis, Richard J., Lupeng Wang, Gongchen Yu, and Leor N. Katz. 2021. What is attention? WIRE Cognitive Science, e1570. [Google Scholar] [CrossRef]
- Lamme, Victor AF. 2003. Why visual attention and awareness are different. Trends in Cognitive Sciences 7: 12–18. [Google Scholar] [CrossRef] [PubMed]
- Land, Michael F. 2014. Do we have an internal model of the outside world? Philosophical Transactions of the Royal Society B: Biological Sciences 369: 20130045. [Google Scholar] [CrossRef]
- Leary, Mark R., and Nicole R. Buttermore. 2003. The evolution of the human self: Tracing the natural history of self-awareness. Journal for the Theory of Social Behaviour 33: 365–404. [Google Scholar] [CrossRef]
- Lou, Hans C., Bruce Luber, Michael Crupain, Julian P. Keenan, Markus Nowak, Troels W. Kjaer, Harold A. Sackeim, and Sarah H. Lisanby. 2004. Parietal cortex and representation of the mental self. Proceedings of the National Academy of Sciences USA 101: 6827–32. [Google Scholar] [CrossRef] [PubMed]
- Lutz, Antoine, Heleen A. Slagter, John D. Dunne, and Richard J. Davidson. 2008. Attention regulation and monitoring in meditation. Trends in Cognitive Sciences 12: 163–69. [Google Scholar] [CrossRef]
- MacLeod, Jeffrey W., Michael A. Lawrence, Meghan M. McConnell, Gail A. Eskes, Raymond M. Klein, and David I. Shore. 2010. Appraising the ANT: Psychometric and theoretical considerations of the Attention Network Test. Neuropsychology 24: 637. [Google Scholar] [CrossRef]
- Maister, Lara, Mel Slater, Maria V. Sanchez-Vives, and Manos Tsakiris. 2015. Changing bodies changes minds: Owning another body affects social cognition. Trends in Cognitive Sciences 19: 6–12. [Google Scholar] [CrossRef]
- Malafouris, Lambros. 2010. The brain–artefact interface (BAI): A challenge for archeology and cultural neuroscience. Social Cognitive and Affective Neuroscience 5: 264–73. [Google Scholar] [CrossRef] [PubMed]
- Margulies, Daniel S., Justin L. Vincent, Clare Kelly, Gabriele Lohmann, Lucina Q. Uddin, Bharat B. Biswal, Arno Villringer, F. Xavier Castellanos, Michael P. Milham, and Michael Petrides. 2009. Precuneus shares intrinsic functional architecture in humans and monkeys. Proceedings of the National Academy of Sciences USA 106: 20069–74. [Google Scholar] [CrossRef]
- Meteyard, Lotte, Sara Rodriguez Cuadrado, Bahador Bahrami, and Gabriella Vigliocco. 2012. Coming of age: A review of embodiment and the neuroscience of semantics. Cortex 48: 788–804. [Google Scholar] [CrossRef]
- Northoff, Georg, and Felix Bermpohl. 2004. Cortical midline structures and the self. Trends in Cognitive Sciences 8: 102–7. [Google Scholar] [CrossRef] [PubMed]
- Osiurak, François, Nicolas Claidière, Alexandre Bluet, Joël Brogniart, Salomé Lasserre, Timothé Bonhoure, Laura Di Rollo, Néo Gorry, Yohann Polette, Alix Saude, and et al. 2022. Technical reasoning bolsters cumulative technological culture through convergent transformations. Science Advances 8: eabl7446. [Google Scholar] [CrossRef]
- Peer, Michael, Roy Salomon, Ilan Goldberg, Olaf Blanke, and Shahar Arzy. 2015. Brain system for mental orientation in space, time, and person. Proceeding of the National Academy of Science USA 112: 11072–77. [Google Scholar] [CrossRef]
- Pereira-Pedro, Ana Sofia, Emiliano Bruner, Philipp Gunz, and Simon Neubauer. 2020. A morphometric comparison of the parietal lobe in modern humans and Neanderthals. Journal of Human Evolution 142: 102770. [Google Scholar] [CrossRef] [PubMed]
- Petersen, Steven E., and Michael I. Posner. 2012. The attention system of the human brain: 20 years after. Annual Review of Neuroscience 35: 73–89. [Google Scholar] [CrossRef] [PubMed]
- Pilacinski, Artur, Stella De Haan, Rita Donato, and Jorge Almeida. 2021. Tool heads prime saccades. Scientific Reports 11: 1–8. [Google Scholar] [CrossRef]
- Ponce de León, Marcia S., Thibault Bienvenu, Assaf Marom, Silvano Engel, Paul Tafforeau, José Luis Alatorre Warren, David Lordkipanidze, Iwan Kurniawan, Delta Bayu Murti, Rusyad Adi Suriyanto, and et al. 2021. The primitive brain of early Homo. Science 372: 165–71. [Google Scholar] [CrossRef]
- Posner, Michael I. 2023. The evolution and future development of attention networks. Journal of Intelligence 11: 98. [Google Scholar] [CrossRef]
- Posner, Michael I., and Mary K. Rothbart. 2023. Fifty years integrating neurobiology and psychology to study attention. Biological Psychology 180: 108574. [Google Scholar] [CrossRef]
- Posner, Michael I., John A. Walker, Frances J. Friedrich, and Robert D. Rafal. 1984. Effects of parietal injury on covert orienting of attention. Journal of Neuroscience 4: 1863–74. [Google Scholar] [CrossRef] [PubMed]
- Raz, Amir. 2004. Anatomy of attentional networks. The Anatomical Record 281: 21–36. [Google Scholar] [PubMed]
- Robbins, Trevor W. 1997. Arousal systems and attentional processes. Biological Psychology 45: 57–71. [Google Scholar]
- Robertson, Lynn C. 2003. Binding, spatial attention and perceptual awareness. Nature Reviews Neuroscience 4: 93–102. [Google Scholar] [CrossRef] [PubMed]
- Rueda, M. Rosario. 2018. Attention in the heart of intelligence. Trends in Neuroscience and Education 13: 26–33. [Google Scholar]
- Rueda, M. Rosario, Sebastian Moyano, and Josué Rico-Picó. 2023. Attention: The grounds of self-regulated cognition. WIRE Cognitive Science 14: e1582. [Google Scholar] [CrossRef]
- Rushworth, Matthew FS, T. Paus, and Perttu K. Sipila. 2001. Attention systems and the organization of the human parietal cortex. Journal of Neuroscience 21: 5262–71. [Google Scholar] [CrossRef] [PubMed]
- Scheperjans, Filip, Klaudia Hermann, Simon B. Eickhoff, Katrin Amunts, Axel Schleicher, and Karl Zilles. 2008. Observer-independent cytoarchitectonic mapping of the human superior parietal cortex. Cerebral Cortex 18: 846–67. [Google Scholar] [PubMed]
- Scolari, Miranda, Katharina N. Seidl-Rathkopf, and Sabine Kastner. 2015. Functions of the human frontoparietal attention network: Evidence from neuroimaging. Current Opinion in Behavioral Sciences 1: 32–39. [Google Scholar] [CrossRef] [PubMed]
- Semendeferi, Katerina, A. Lu, Natalie Schenker, and Hanna Damásio. 2002. Humans and great apes share a large frontal cortex. Nature Neuroscience 5: 272–76. [Google Scholar] [CrossRef]
- Sharon, Gonen, and Maya Oron. 2014. The lithic tool arsenal of a Mousterian hunter. Quaternary International 331: 167–85. [Google Scholar] [CrossRef]
- Shea, John J. 2013. Lithic modes A–I: A new framework for describing global-scale variation in stone tool technology illustrated with evidence from the East Mediterranean Levant. Journal of Archeological Method and Theory 20: 151–86. [Google Scholar]
- Shea, John J. 2017. Occasional, obligatory, and habitual stone tool use in hominin evolution. Evolutionary Anthropology 26: 200–17. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, Chet C., and Aida Gómez-Robles. 2017. Brain plasticity and human evolution. Annual Review of Anthropology 46: 399–419. [Google Scholar] [CrossRef]
- Sherwood, Chet C., and Jeroen B. Smaers. 2013. What’s the fuss over human frontal lobe evolution? Trends in Cognitive Sciences 17: 432–33. [Google Scholar] [CrossRef] [PubMed]
- Shumaker, Robert W., Kristina R. Walkup, and Benjamin B. Beck. 2011. Animal Tool Behavior: The Use and Manufacture of Tools by Animals. Baltimore: Johns Hopkins University Press. [Google Scholar]
- Silva-Gago, María, and Emiliano Bruner. 2023. Cognitive archeology, attention and visual behaviour. In Cognitive Archeology, Body Cognition, and the Evolution of Visuospatial Perception. Edited by E. Bruner. London: Academic Press, in press. [Google Scholar]
- Silva-Gago, María, Annapaola Fedato, Timothy Hodgson, Marcos Terradillos-Bernal, Rodrigo Alonso-Alcalde, and Emiliano Bruner. 2021. Visual attention reveals affordances during Lower Palaeolithic stone tool exploration. Archeological and Anthropological Sciences 13: 145. [Google Scholar] [CrossRef]
- Silva-Gago, María, Flora Ioannidou, Annapaola Fedato, Timothy Hodgson, and Emiliano Bruner. 2022. Visual attention and cognitive archeology: An eye-tracking study of Palaeolithic stone tools. Perception 51: 3–24. [Google Scholar] [CrossRef]
- Stout, D. 2010. The evolution of cognitive control. Topics in Cognitive Science 2: 614–30. [Google Scholar] [CrossRef] [PubMed]
- Sumantry, David, and Kathleen E. Stewart. 2021. Meditation, mindfulness, and attention: A meta-analysis. Mindfulness 12: 1332–49. [Google Scholar]
- Sznajder, Beata, Maurice W. Sabelis, and Martijn Egas. 2012. How adaptive learning affects evolution: Reviewing theory on the Baldwin effect. Evolutionary Biology 39: 301–10. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, Yoshinori, Satoshi Nobusako, Yusaku Takamura, Yu Miyawaki, Moe Terada, and Shu Morioka. 2020. Effects of tool novelty and action demands on gaze searching during tool observation. Frontiers in Psychology 11: 587270. [Google Scholar] [CrossRef]
- Tang, Yi-Yuan, and Michael I. Posner. 2009. Attention training and attention state training. Trends in Cognitive Sciences 13: 222–27. [Google Scholar] [CrossRef] [PubMed]
- Tang, Yi-Yuan, Britta K. Hölzel, and Michael I. Posner. 2015. The neuroscience of mindfulness meditation. Nature Reviews Neuroscience 16: 213–25. [Google Scholar] [CrossRef] [PubMed]
- Tobias, Phillip V. 1987. The brain of Homo habilis: A new level of organization in cerebral evolution. Journal of Human Evolution 16: 741–61. [Google Scholar] [CrossRef]
- Turq, Alain, Wil Roebroeks, Laurence Bourguignon, and Jean-Philippe Faivre. 2013. The fragmented character of Middle Palaeolithic stone tool technology. Journal of Human Evolution 65: 641–55. [Google Scholar] [CrossRef] [PubMed]
- Vaishnavi, S. Neil, Andrei G. Vlassenko, Melissa M. Rundle, Abraham Z. Snyder, Mark A. Mintun, and Marcus E. Raichle. 2010. Regional aerobic glycolysis in the human brain. Proceedings of the National Academy of Sciences USA 107: 17757–62. [Google Scholar] [CrossRef] [PubMed]
- Whiten, Andrew, Jane Goodall, William C. McGrew, Toshisada Nishida, Vernon Reynolds, Yukimaru Sugiyama, Caroline EG Tutin, Richard W. Wrangham, and Christophe Boesch. 1999. Cultures in chimpanzees. Nature 399: 682–85. [Google Scholar] [CrossRef]
- Willbrand, Ethan H., Samira A. Maboudian, Joseph P. Kelly, Benjamin J. Parker, Brett L. Foster, and Kevin S. Weiner. 2023. Sulcal morphology of posteromedial cortex substantially differs between humans and chimpanzees. Communications Biology 6: 586. [Google Scholar] [CrossRef]
- Wilson, Margaret. 2002. Six views of embodied cognition. Psychonomic Bulletin & Review 9: 625–36. [Google Scholar]
- Wilson, Margaret. 2010. The re-tooled mind: How culture re-engineers cognition. Social Cognitive and Affective Neuroscience 5: 180–87. [Google Scholar] [CrossRef]
- Wood, Bernard. 2014. Human evolution: Fifty years after Homo habilis. Nature 508: 31–33. [Google Scholar] [CrossRef][Green Version]
- Wynn, Thomas, and Frederick L. Coolidge. 2004. The expert Neandertal mind. Journal of Human Evolution 46: 467–87. [Google Scholar] [CrossRef]
- Wynn, Thomas, and Frederick L. Coolidge. 2010. Beyond symbolism and language. Current Anthropology 51: S5–S16. [Google Scholar] [CrossRef]
- Wynn, Thomas, and Frederick L. Coolidge. 2016. Archeological insights into hominin cognitive evolution. Evolutionary Anthropology 25: 200–13. [Google Scholar] [CrossRef] [PubMed]
- Wynn, Thomas, and John Gowlett. 2018. The handaxe reconsidered. Evolutionary Anthropology 27: 21–29. [Google Scholar] [CrossRef] [PubMed]
- Wynn, Thomas, Karenleigh A. Overmann, and Frederick L. Coolidge. 2016. The false dichotomy: A refutation of the Neandertal indistinguishability claim. Journal of Anthropological Sciences 94: 1–22. [Google Scholar]
- Wynn, Thomas, R. Adriana Hernandez-Aguilar, Linda F. Marchant, and William C. McGrew. 2011. “An ape’s view of the Oldowan” revisited. Evolutionary Anthropology 20: 181–97. [Google Scholar] [CrossRef]
- Zhang, Sheng, and Chiang-shan R. Li. 2012. Functional connectivity mapping of the human precuneus by resting state fMRI. Neuroimage 59: 3548–62. [Google Scholar] [CrossRef]
- Zollikofer, Christoph PE, Thibault Bienvenu, Yonas Beyene, Gen Suwa, Berhane Asfaw, Tim D. White, and Marcia S. Ponce de León. 2022. Endocranial ontogeny and evolution in early Homo sapiens: The evidence from Herto, Ethiopia. Proceedings of the National Academy of Sciences USA 119: e2123553119. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruner, E. Cognitive Archeology and the Attentional System: An Evolutionary Mismatch for the Genus Homo. J. Intell. 2023, 11, 183. https://doi.org/10.3390/jintelligence11090183
Bruner E. Cognitive Archeology and the Attentional System: An Evolutionary Mismatch for the Genus Homo. Journal of Intelligence. 2023; 11(9):183. https://doi.org/10.3390/jintelligence11090183
Chicago/Turabian StyleBruner, Emiliano. 2023. "Cognitive Archeology and the Attentional System: An Evolutionary Mismatch for the Genus Homo" Journal of Intelligence 11, no. 9: 183. https://doi.org/10.3390/jintelligence11090183
APA StyleBruner, E. (2023). Cognitive Archeology and the Attentional System: An Evolutionary Mismatch for the Genus Homo. Journal of Intelligence, 11(9), 183. https://doi.org/10.3390/jintelligence11090183