
Effect of Key Phytochemicals from Andrographis paniculata, Tinospora cordifolia, and Ocimum sanctum on PLpro-ISG15 De-Conjugation Machinery—A Computational Approach

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Screening of Phytochemicals of Andrographis paniculata (AG), Tinospora cordifolia (GU), and Ocimum sanctum (TU) against SARS-CoV-2 PLpro ISG15 Site at UIM (PDB:6XA9)
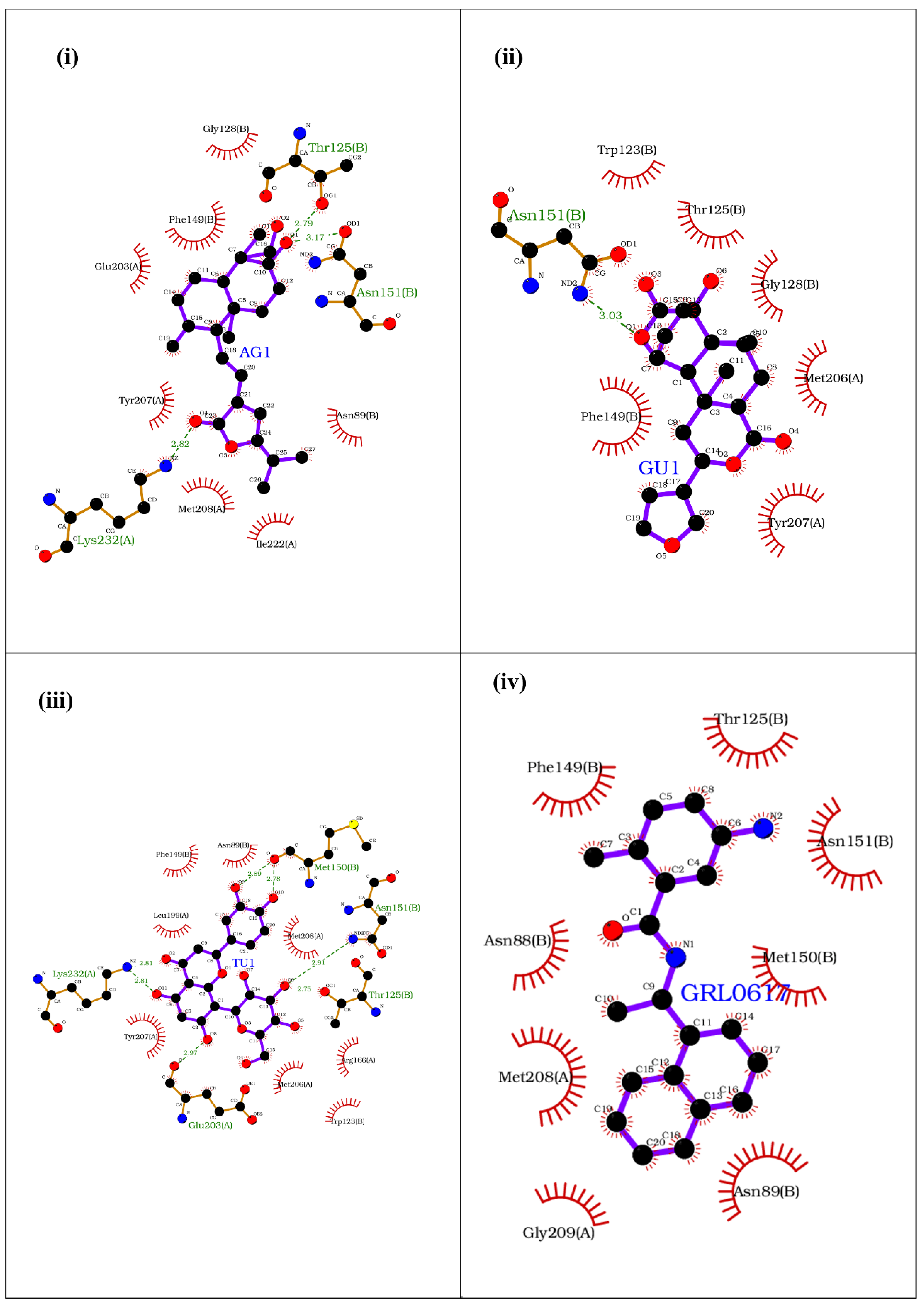
Docking against SARS-CoV-2 PLpro ISG15 Interacting Site
2.2. Physiochemical Properties Analysis of the Phytoactives Affirms Their Drug-Likeness
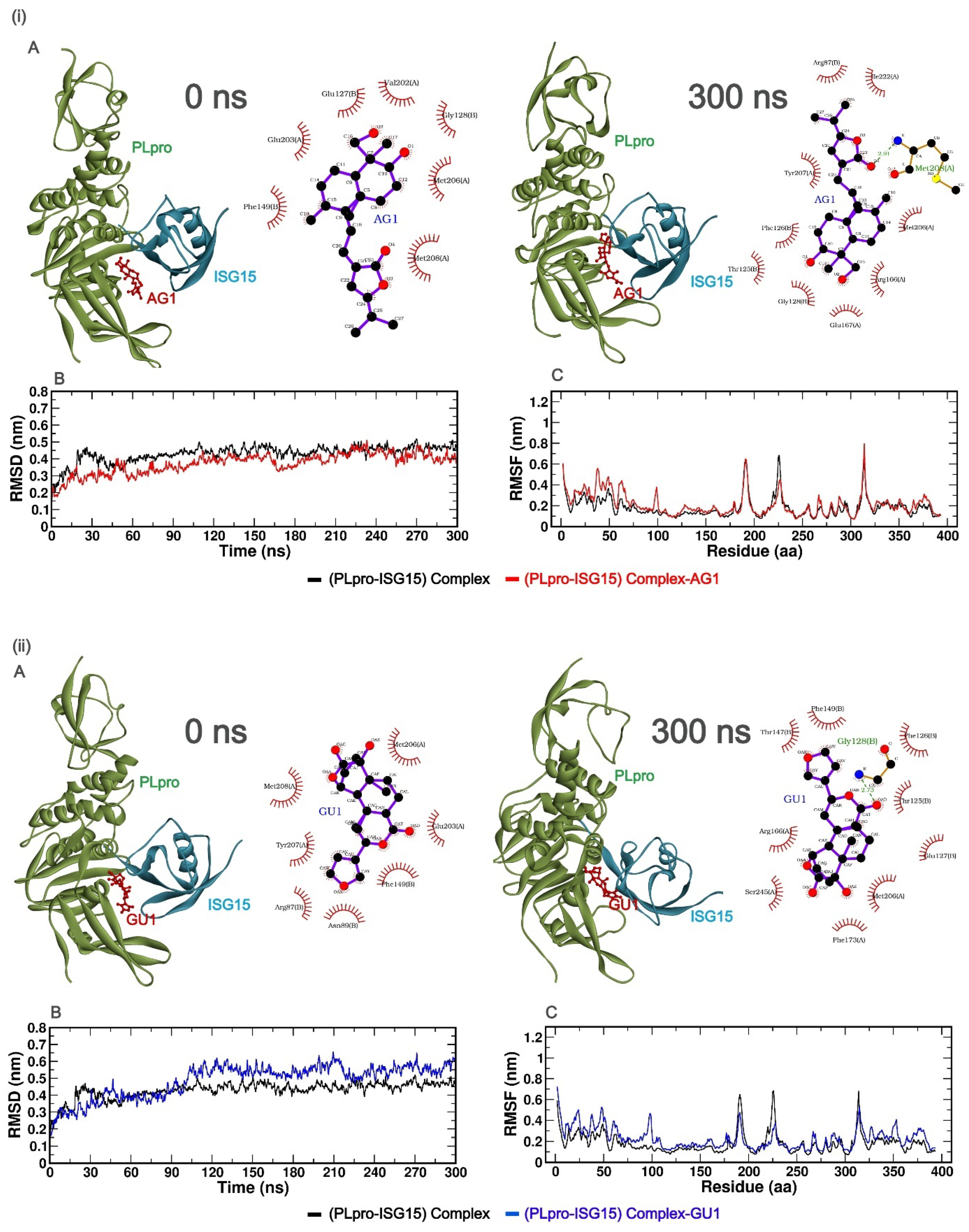
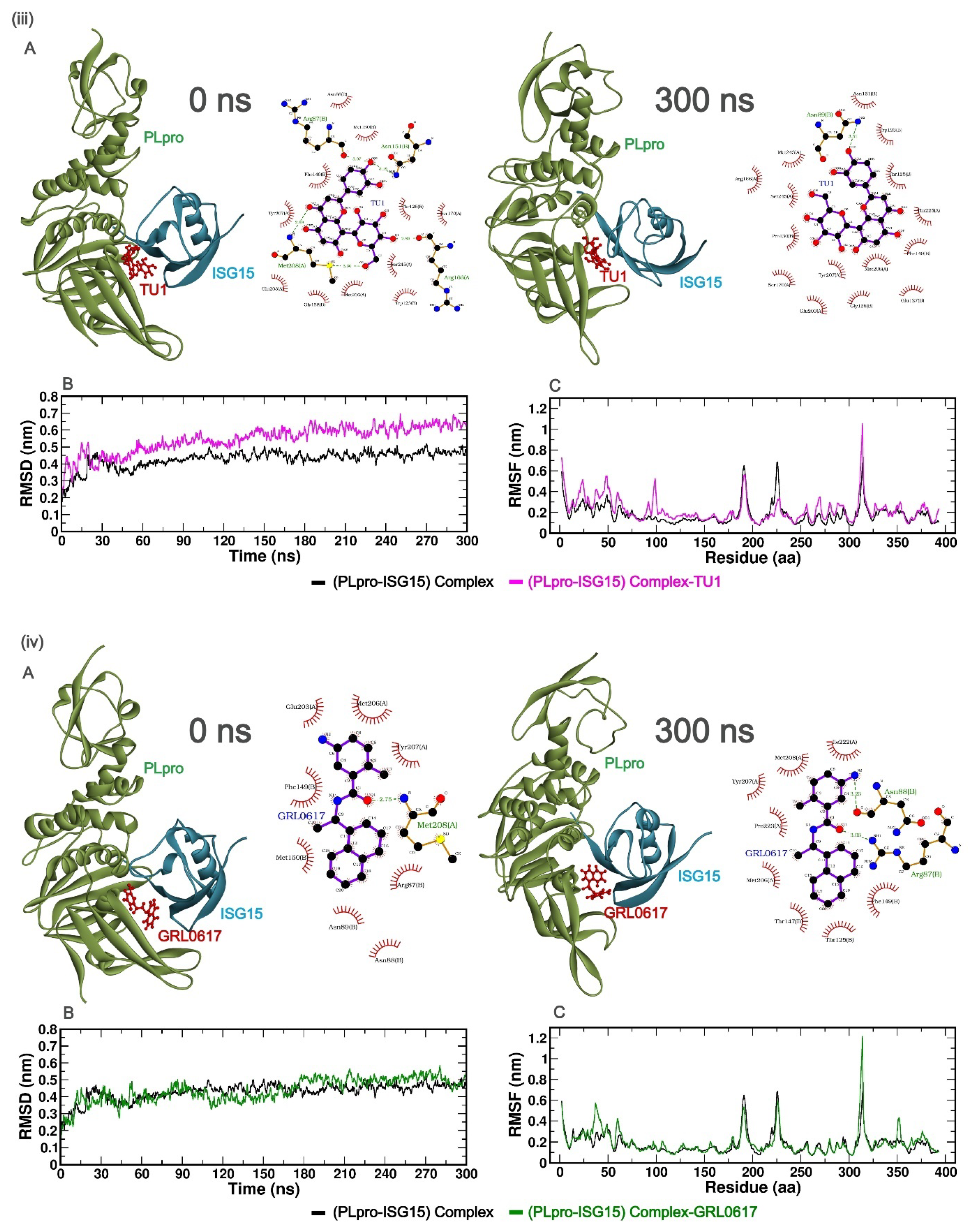
2.3. Molecular Dynamics (MD) Simulation Study
2.3.1. Stability and Fluctuations of the Protein: RMSD and RMSF Analysis of the Protein Complex
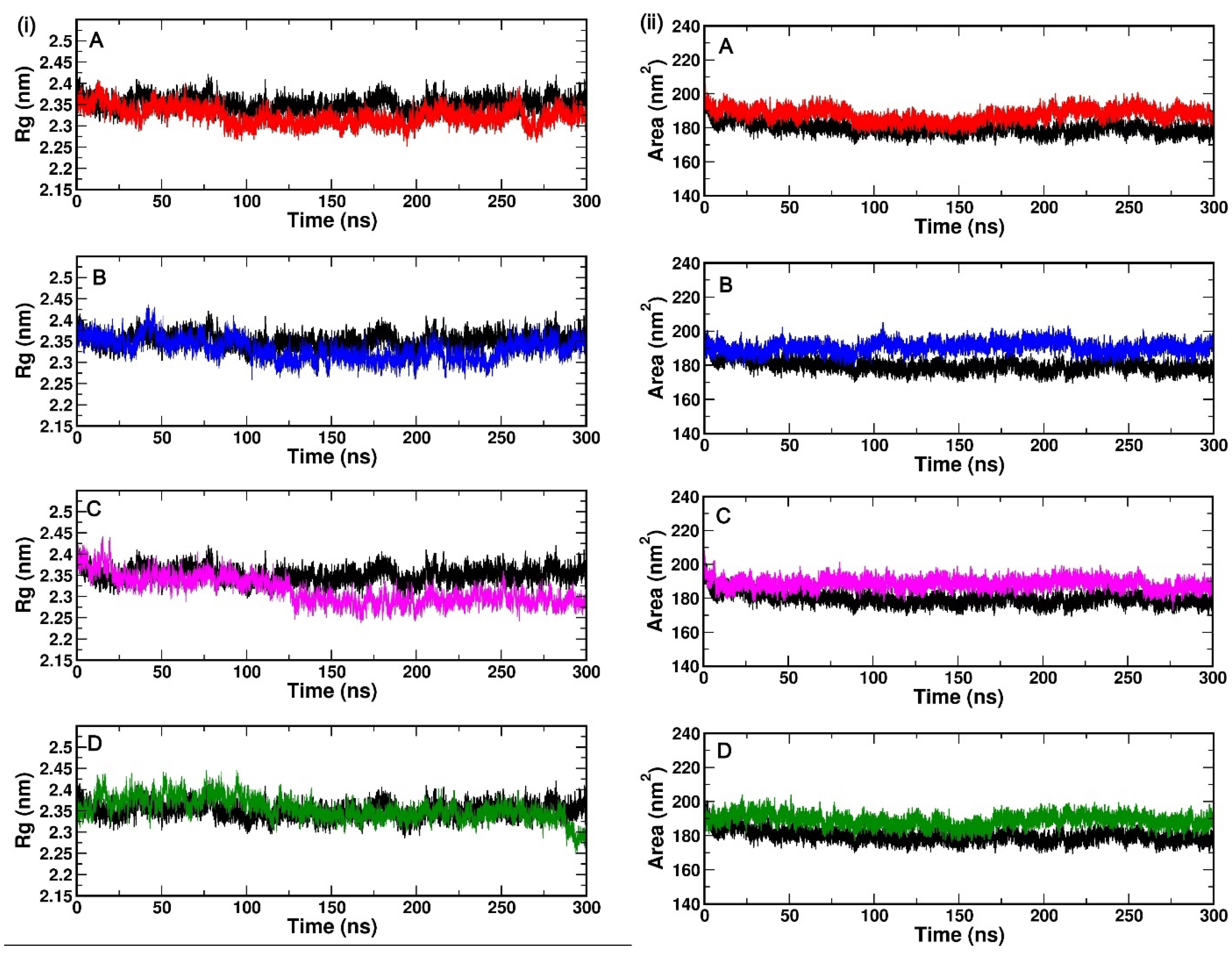
2.3.2. Compactness of Protein Complex: Radius of Gyration (Rg) and Solvent Accessible Surface Area (SASA)
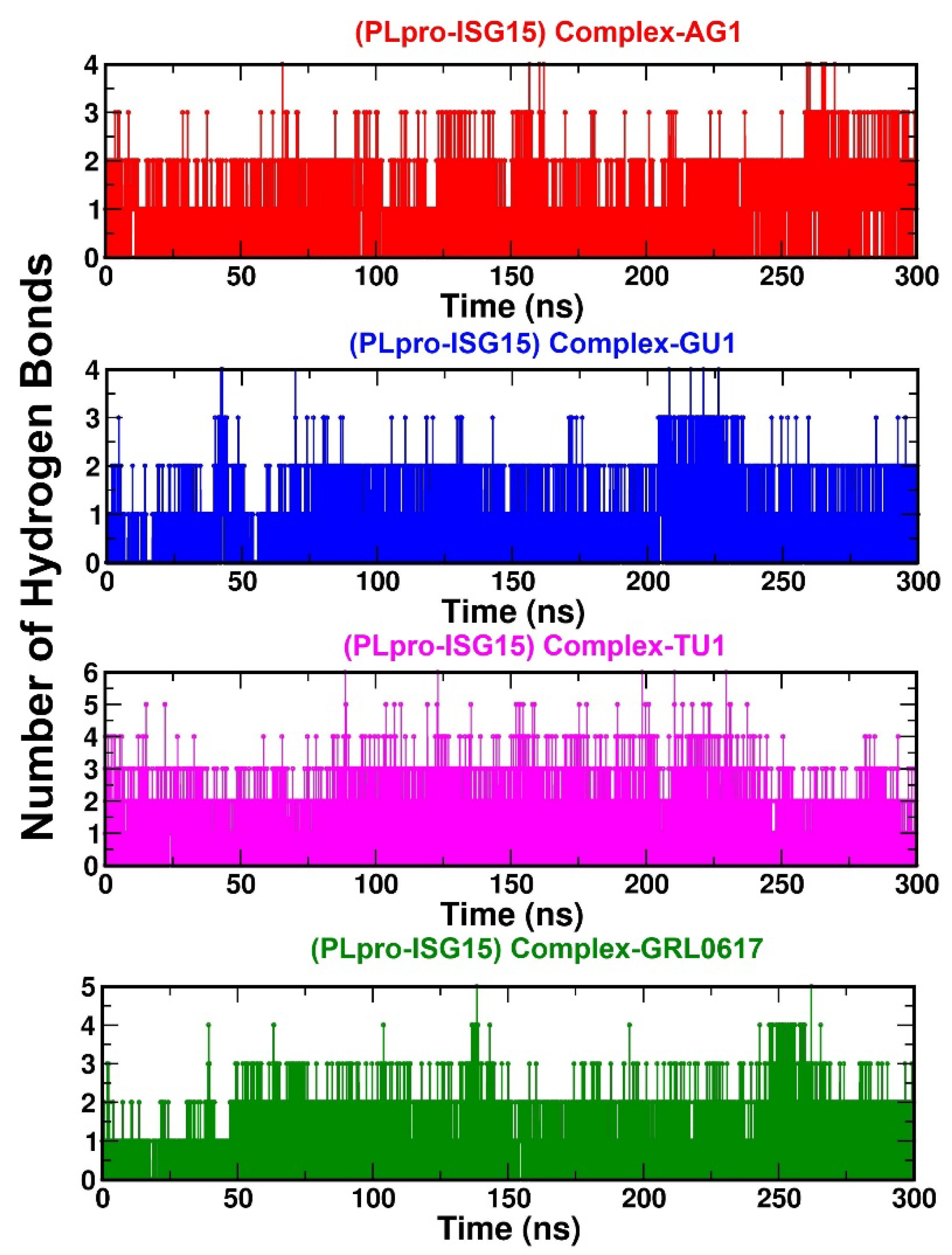
2.3.3. Interactions between the Protein–Ligand Complex: Hydrogen Bond (H-Bond)
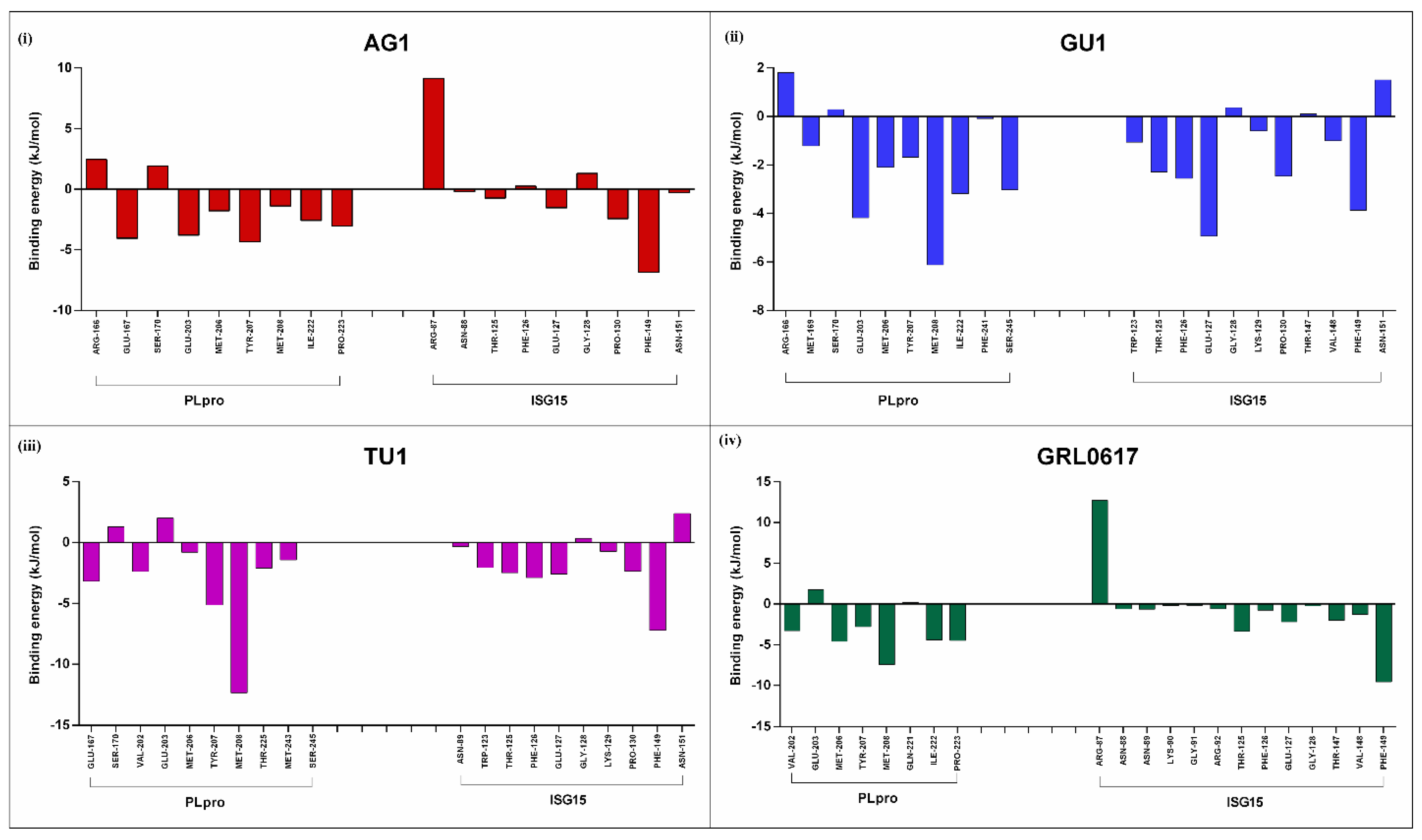
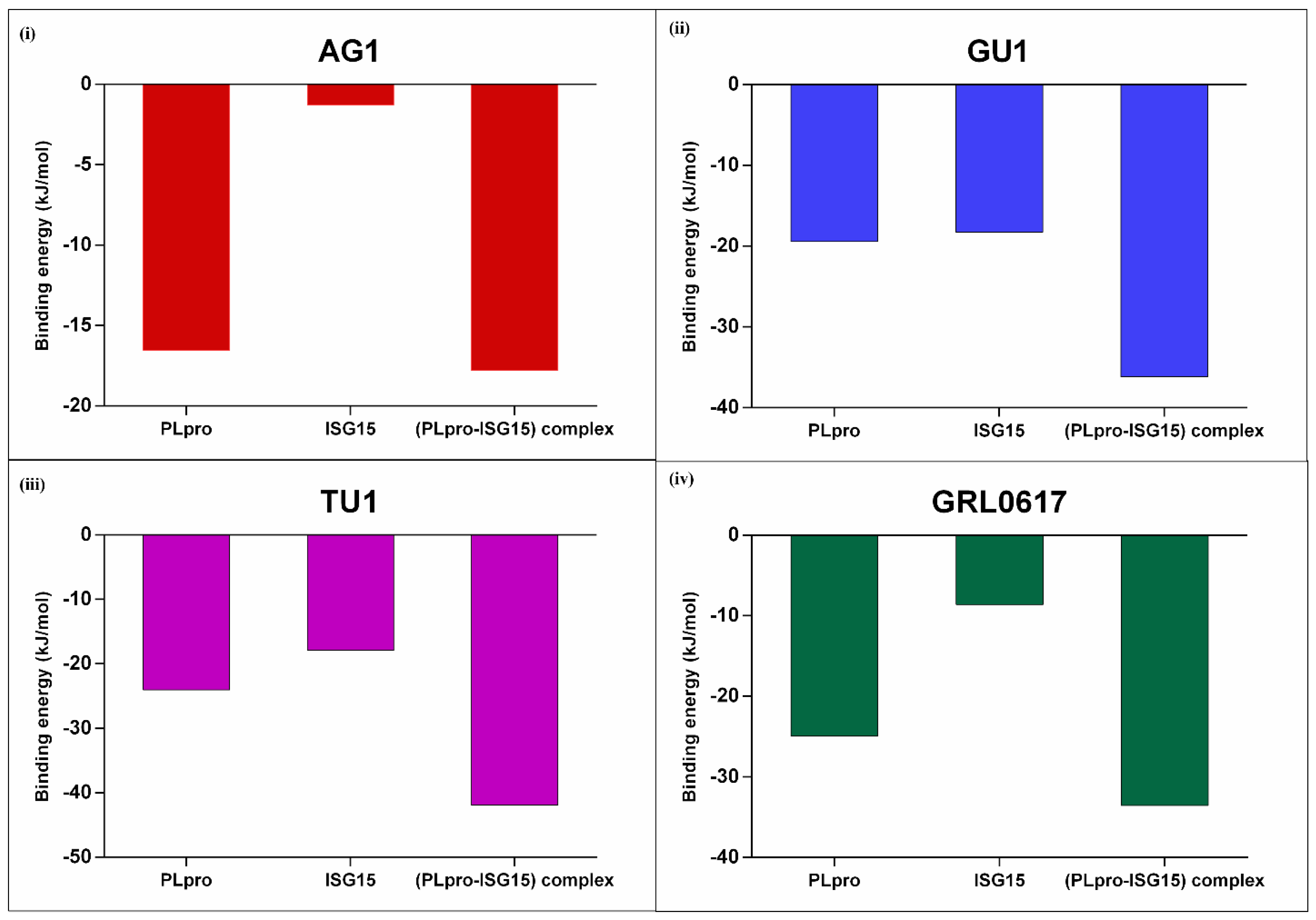
2.3.4. Binding Affinities of Phytochemicals to the Interface of PLpro-ISG15 Complex: MMPBSA Based Calculations
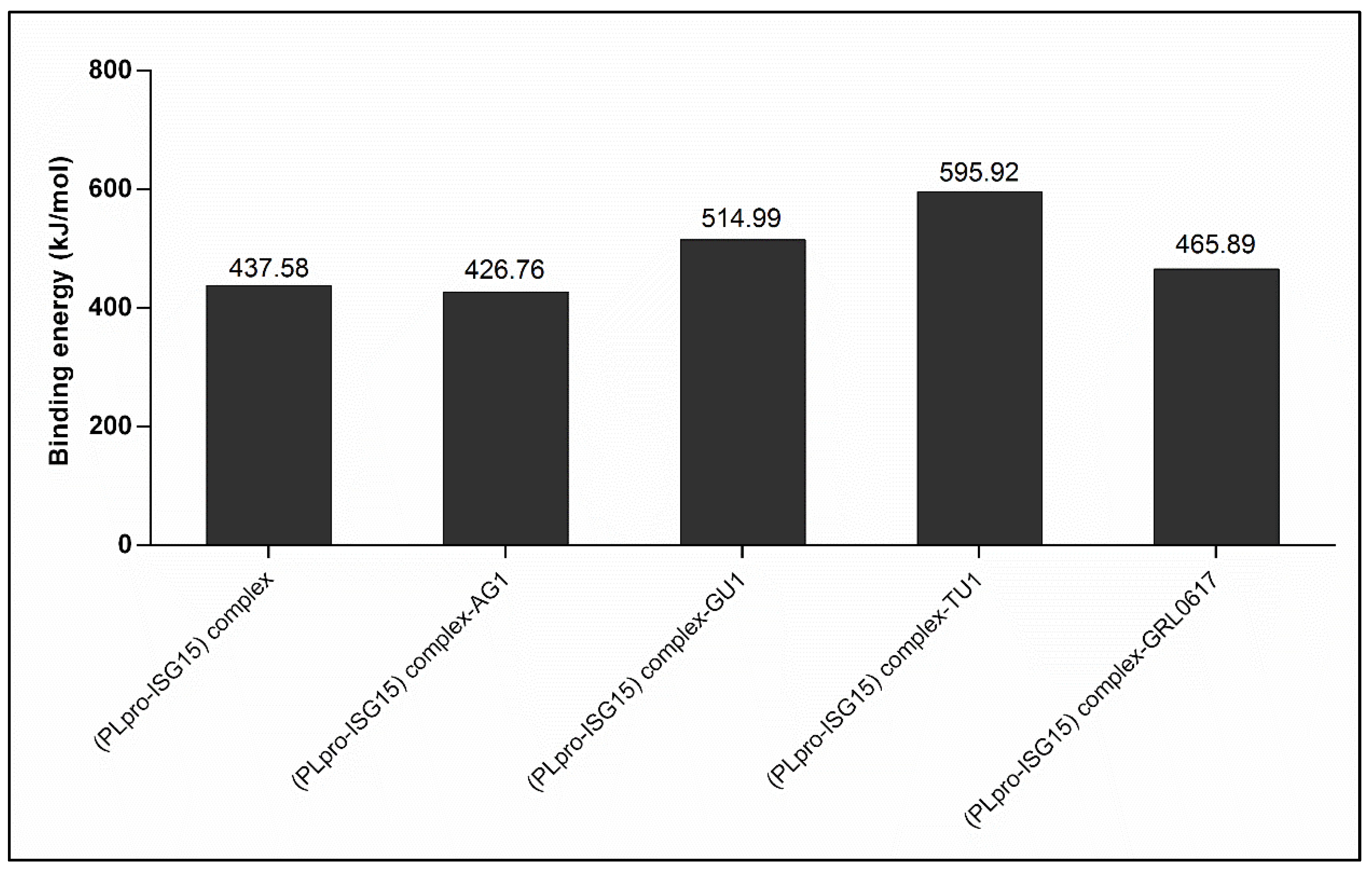
2.3.5. Effect of Phytochemicals on Protein–Protein Interactions between (PLpro-ISG15)-Complex
3. Materials and Methods
3.1. Target Enzyme Preparation
3.2. Ligand Preparation
3.3. Docking and Visualization
3.4. Drug Likeness Study and ADME Screening
3.5. Molecular Dynamic Simulations and Free Energy Calculation (MM-PBSA)
- (i)
- (PLpro-ISG15)-complex
- (ii)
- (PLpro-ISG15)-complex with AG1
- (iii)
- (PLpro-ISG15)-complex with GU1
- (iv)
- (PLpro-ISG15)-complex withTU1 and
- (v)
- (PLpro-ISG15)-complex with GRL0617
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADME | Absorption, distribution, metabolism, elimination |
| AG | Andrographis paniculata |
| AG1 | 14-deoxy-15-isopropylidene-11,12-didehydroandrographolide |
| GU | Tinospora cordifolia |
| GU1 | Isocolumbin |
| ISGs Interferon-stimulated genes | |
| MM/PBSA | Molecular mechanics/Poisson–Boltzmann surface area |
| MD | Molecular dynamics |
| PDB | Protein data bank |
| SPC | Simple point charge |
| TU | Ocimum sanctum |
| TU1 | Orientin |
| UIM | Ubiquitin-interacting motif |
References
- Jeon, Y.J.; Yoo, H.M.; Chung, C.H. ISG15 and immune diseases. Biochim. Biophys. Acta 2010, 1802, 485–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Zhang, D.E. Interferon-Stimulated Gene 15 and the Protein ISGylation System. J. Interf. Cytokine Res. 2011, 31, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, B.T.; Scholte, F.E.M.; Bergeron, É.; Pegan, S.D. How ISG15 combats viral infection. Virus Res. 2020, 286, 198036. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, J.; Yan, H.; Huang, J.; Wang, F.; Liu, T.; Zeng, L.; Zhou, F. ISGylation in Innate Antiviral Immunity and Pathogen Defense Responses: A Review. Front. Cell Dev. Biol. 2021, 9, 3196. [Google Scholar] [CrossRef] [PubMed]
- Perng, Y.C.; Lenschow, D.J. ISG15 in antiviral immunity and beyond. Nat. Rev. Microbiol. 2018, 16, 423–439. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Luan, J.; Zhang, L. Molecular docking of potential SARS-CoV-2 papain-like protease inhibitors. Biochem. Biophys. Res. Commun. 2021, 538, 72–79. [Google Scholar] [CrossRef]
- Fung, S.Y.; Yuen, K.S.; Ye, Z.W.; Chan, C.P.; Jin, D.Y. A tug-of-war between severe acute respiratory syndrome coronavirus 2 and host antiviral defence: Lessons from other pathogenic viruses. Emerg. Microbes Infect. 2020, 9, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Shemesh, M.; Aktepe, T.E.; Deerain, J.M.; McAuley, J.L.; Audsley, M.D.; David, C.T.; Purcell, D.F.J.; Urin, V.; Hartmann, R.; Moseley, G.W.; et al. SARS-CoV-2 suppresses IFNβ production mediated by NSP1, 5, 6, 15, ORF6 and ORF7b but does not suppress the effects of added interferon. PLoS Pathog. 2021, 17, e1009800. [Google Scholar] [CrossRef]
- Kim, Y.M.; Shin, E.C. Type I and III interferon responses in SARS-CoV-2 infection. Exp. Mol. Med. 2021, 53, 750–760. [Google Scholar] [CrossRef]
- Gupta, S.; Mishra, K.P.; Ganju, L. Broad-spectrum antiviral properties of andrographolide. Arch. Virol. 2017, 162, 611–623. [Google Scholar] [CrossRef]
- Wang, W.; Wang, J.; Dong, S.F.; Liu, C.H.; Italiani, P.; Sun, S.H.; Xu, J.; Boraschi, D.; Ma, S.P.; Qu, D. Immunomodulatory activity of andrographolide on macrophage activation and specific antibody response. Acta Pharmacol. Sin. 2010, 31, 191–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niraj, S.; Varsha, S. A review on scope of immuno-modulatory drugs in Ayurveda for prevention and treatment of COVID-19. Plant. Sci. Today 2020, 7, 417–423. [Google Scholar] [CrossRef]
- Borse, S.; Joshi, M.; Saggam, A.; Bhat, V.; Walia, S.; Marathe, A.; Sagar, S.; Chavan-Gautam, P.; Girme, A.; Hingorani, L.; et al. Ayurveda botanicals in COVID-19 management: An in silico multi-target approach. PLoS ONE 2021, 16, e0248479. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.S.; Panda, S.K.; Luyten, W. Phytochemicals against SARS-CoV as potential drug leads. Biomed. J. 2021, 44, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Krupanidhi, S.; Peele, K.A.; Venkateswarulu, T.C.; Ayyagari, V.S.; Bobby, M.N.; Babu, D.J.; Narayana, A.V.; Aishwarya, G. Screening of phytochemical compounds of Tinospora cordifolia for their inhibitory activity on SARS-CoV-2: An in silico study. J. Biomol. Struct. Dyn. 2021, 39, 5799–5803. [Google Scholar] [CrossRef]
- Klemm, T.; Ebert, G.; Calleja, D.J.; Allison, C.C.; Richardson, L.W.; Bernardini, J.P.; Lu, B.G.; Kuchel, N.W.; Grohmann, C.; Shibata, Y.; et al. Mechanism and inhibition of the papain-like protease, PLpro, of SARS-CoV-2. EMBO J. 2020, 39, e106275. [Google Scholar] [CrossRef]
- Anil, K.T.J.W. Autodock vina: Improving the speed and accuracy of docking. J. Comput. Chem. 2019, 31, 455–461. [Google Scholar] [CrossRef]
- Shin, D.; Mukherjee, R.; Grewe, D.; Bojkova, D.; Baek, K.; Bhattacharya, A.; Schulz, L.; Widera, M.; Mehdipour, A.R.; Tascher, G.; et al. Papain-like protease regulates SARS-CoV-2 viral spread and innate immunity. Nature 2020, 587, 657–662. [Google Scholar] [CrossRef]
- Freitas, B.T.; Durie, I.A.; Murray, J.; Longo, J.E.; Miller, H.C.; Crich, D.; Hogan, R.J.; Tripp, R.A.; Pegan, S.D. Characterization and Noncovalent Inhibition of the Deubiquitinase and deISGylase Activity of SARS-CoV-2 Papain-Like Protease. ACS Infect. Dis. 2020, 6, 2099–2109. [Google Scholar] [CrossRef]
- Fu, Z.; Huang, B.; Tang, J.; Liu, S.; Liu, M.; Ye, Y.; Liu, Z.; Xiong, Y.; Zhu, W.; Cao, D.; et al. The complex structure of GRL0617 and SARS-CoV-2 PLpro reveals a hot spot for antiviral drug discovery. Nat. Commun. 2021, 12, 488. [Google Scholar] [CrossRef]
- Islam, M.T.; Bardaweel, S.K.; Mubarak, M.S.; Koch, W.; Gaweł-Beben, K.; Antosiewicz, B.; Sharifi-Rad, J. Immunomodulatory Effects of Diterpenes and Their Derivatives Through NLRP3 Inflammasome Pathway: A Review. Front. Immunol. 2020, 11, 2234. [Google Scholar] [CrossRef] [PubMed]
- Wardana, A.P.; Aminah, N.S.; Rosyda, M.; Abdjan, M.I.; Kristanti, A.N.; Tun, K.N.W.; Choudhary, M.I.; Takaya, Y. Potential of diterpene compounds as antivirals, a review. Heliyon 2021, 7, e07777. [Google Scholar] [CrossRef] [PubMed]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. LIGPLOT: A program to generate schematic diagrams of protein-ligand interactions. Protein. Eng. Des. Sel. 1995, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, Q.; Yuan, Z.F.; Zhang, L.; Xu, L.; Cui, Y.; Duan, K. Pharmacokinetics and tissue distribution study of orientin in rat by liquid chromatography. J. Pharm. Biomed. Anal. 2008, 47, 429–434. [Google Scholar] [CrossRef]
- Wang, C.; Nguyen, P.H.; Pham, K.; Huynh, D.; Le, T.B.N.; Wang, H.; Ren, P.; Luo, R. Calculating protein-ligand binding affinities with MMPBSA: Method and error analysis. J. Comput. Chem. 2016, 37, 2436–2446. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.R.; Lightstone, F.C.; Cheng, F. In Silico Insights into Protein–Protein Interaction Disruptive Mutations in the PCSK9-LDLR Complex. Int. J. Mol. Sci. 2020, 21, 1550. [Google Scholar] [CrossRef] [Green Version]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An Open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Biovia: Discovery Studio Modeling Environment-Google Scholar. Available online: https://scholar.google.com/scholar?cluster=17675170202455151209&hl=en&as_sdt=2005&sciodt=0,5 (accessed on 8 June 2022).
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- PreADME in EuroQSAR 2004–PreADMET|Prediction of ADME/Tox. 2004. Available online: https://preadmet.webservice.bmdrc.org/2004/09/27/200409-preadme-in-euroqsar-2004/ (accessed on 7 June 2022).
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindah, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Schüttelkopf, A.W.; Van Aalten, D.M.F. PRODRG: A tool for high-throughput crystallography of protein-ligand complexes. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 1355–1363. [Google Scholar] [CrossRef] [Green Version]
- Gangadharappa, B.S.; Sharath, R.; Revanasiddappa, P.D.; Chandramohan, V.; Balasubramaniam, M.; Vardhineni, T.P. Structural insights of metallo-beta-lactamase revealed an effective way of inhibition of enzyme by natural inhibitors. J. Biomol. Struct. Dyn. 2020, 38, 3757–3771. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Kumar, R.; Lynn, A. g_mmpbsa–a GROMACS tool for high-throughput MM-PBSA calculations. J. Chem. Inf. Model. 2014, 54, 1951–1962. [Google Scholar] [CrossRef] [PubMed]
- Villarroya-Beltri, C.; Guerra, S.; Sánchez-Madrid, F. ISGylation-a key to lock the cell gates for preventing the spread of threats. J. Cell Sci. 2017, 130, 2961–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Herb | Phytochemical | PubChem CID | Canonical SMILES | ∆G (kcal/mol) |
|---|---|---|---|---|---|
| Plpro ISG15 Interaction Site at UIM (PDB: 6XA9) | |||||
| 1 | Andrographis paniculata | 14-deoxy-15-isopropylidene-11,12-didehydroandrographolide | 637300 | CC(=C1C=C(C(=O)O1)C=CC2C(=C)CCC3C2(CCC(C3(C)CO)O)C)C | −9.4 |
| 2 | Andrographolactone | 44206466 | CC1=CC2=C(CCC1)C(=C(C(=C2)C)CCC3=CCOC3=O)C | −9.2 | |
| 3 | Neoandrographolide | 9848024 | CC1(CCCC2(C1CCC(=C)C2CCC3=CCOC3=O)C)COC4C(C(C(C(O4)CO)O)O)O | −9 | |
| 4 | 14-Deoxy-11,12-didehydroandrographolide | 5708351 | CC12CCC(C(C1CCC(=C)C2C=CC3=CCOC3=O)(C)CO)O | −8.8 | |
| 5 | Andrographolide | 5318517 | CC12CCC(C(C1CCC(=C)C2CC=C3C(COC3=O)O)(C)CO)O | −8.7 | |
| 6 | Tinospora cordifolia | Isocolumbin | 24721165 | CC12CCC3C(=O)OC(CC3(C1C4C=CC2(C(=O)O4)O)C)C5=COC=C5 | −9.9 |
| 7 | Berberin | 2353 | COC1=C(C2=C[N+]3=C(C=C2C=C1)C4=CC5=C(C=C4CC3)OCO5)OC | −9.4 | |
| 8 | Ecdysterone | 12304165 | CC12CCC3C(=CC(=O)C4C3(CC(C(C4)O)O)C)C1(CCC2C(C)(C(CCC(C)(C)O)O)O)O | −9 | |
| 9 | Magnoflorine | 73337 | C[N+]1(CCC2=CC(=C(C3=C2C1CC4=C3C(=C(C=C4)OC)O)O)OC)C | −9 | |
| 10 | Beta-Sitosterol | 222284 | CCC(CCC(C)C1CCC2C1(CCC3C2CC=C4C3(CCC(C4)O)C)C)C(C)C | −8.4 | |
| 11 | Ocimum sanctum | Orientin | 5281675 | C1=CC(=C(C=C1C2=CC(=O)C3=C(O2)C(=C(C=C3O)O)C4C(C(C(C(O4)CO)O)O)O)O)O | −9.4 |
| 12 | Isoorientin | 114776 | C1=CC(=C(C=C1C2=CC(=O)C3=C(O2)C=C(C(=C3O)C4C(C(C(C(O4)CO)O)O)O)O)O)O | −9.2 | |
| 13 | Vitexin | 5280441 | C1=CC(=CC=C1C2=CC(=O)C3=C(O2)C(=C(C=C3O)O)C4C(C(C(C(O4)CO)O)O)O)O | −9.1 | |
| 14 | Isovitexin | 162350 | C1=CC(=CC=C1C2=CC(=O)C3=C(O2)C=C(C(=C3O)C4C(C(C(C(O4)CO)O)O)O)O)O | −9 | |
| 15 | Molludistin | 44258315 | COC1=C(C2=C(C(=C1)O)C(=O)C=C(O2)C3=CC=C(C=C3)O)C4C(C(C(CO4)O)O)O | −8.8 | |
| 16 | Positive control | GRL0617 | 24941262 | CC1=C(C=C(C=C1)N)C(=O)NC(C)C2=CC=CC3=CC=CC=C32 | −8.5 |
| Interacting Partner | Ligands/Phytochemicals Bound | Common Amino Acids | Number of Common Amino Acids |
|---|---|---|---|
| PLpro | AG1, GRL0617, GU1 TU1 | Glu-203, Met-208, Met-206, Tyr-207 | 4 |
| AG1, GU1, TU1 | Ser-170 | 1 | |
| AG1, GRL0617, GU1 | Ile-222 | 1 | |
| AG1, GU1 | Arg-166 | 1 | |
| AG1, TU1 | Glu-167 | 1 | |
| AG1, GRL0617 | Pro-223 | 1 | |
| GU1, TU1 | Ser-245 | 1 | |
| GRL0617, TU1 | Val-202 | 1 | |
| GU1 | Met-169, Phe-241 | 2 | |
| TU1 | Thr-225, Met-243 | 2 | |
| GRL0617 | Gln-221 | 1 | |
| ISG15 | AG1, GRL0617, GU1, TU1 | Glu-127, Gly-128, Phe-149, Thr-125, Phe-126 | 5 |
| AG1, GU1, TU1 | Pro-130, Asn-151 | 2 | |
| AG1, GRL0617 | Arg-87, Asn-88 | 2 | |
| GU1, TU1 | Lys-129, Trp-123 | 2 | |
| GRL0617, GU1 | Thr-147, Val-148 | 2 | |
| GRL0617, TU1 | Asn-89 | 1 | |
| GRL0617 | Arg-92, Gly-91, Lys-90 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, P.; Bhat, S.S.; Punnapuzha, A.; Bhagavatula, A.; Venkanna, B.U.; Mohamed, R.; Rao, R.P. Effect of Key Phytochemicals from Andrographis paniculata, Tinospora cordifolia, and Ocimum sanctum on PLpro-ISG15 De-Conjugation Machinery—A Computational Approach. Computation 2022, 10, 109. https://doi.org/10.3390/computation10070109
Singh P, Bhat SS, Punnapuzha A, Bhagavatula A, Venkanna BU, Mohamed R, Rao RP. Effect of Key Phytochemicals from Andrographis paniculata, Tinospora cordifolia, and Ocimum sanctum on PLpro-ISG15 De-Conjugation Machinery—A Computational Approach. Computation. 2022; 10(7):109. https://doi.org/10.3390/computation10070109
Chicago/Turabian StyleSingh, Prachi, Shruthi S. Bhat, Ardra Punnapuzha, Amrutha Bhagavatula, Babu U. Venkanna, Rafiq Mohamed, and Raghavendra P. Rao. 2022. "Effect of Key Phytochemicals from Andrographis paniculata, Tinospora cordifolia, and Ocimum sanctum on PLpro-ISG15 De-Conjugation Machinery—A Computational Approach" Computation 10, no. 7: 109. https://doi.org/10.3390/computation10070109
APA StyleSingh, P., Bhat, S. S., Punnapuzha, A., Bhagavatula, A., Venkanna, B. U., Mohamed, R., & Rao, R. P. (2022). Effect of Key Phytochemicals from Andrographis paniculata, Tinospora cordifolia, and Ocimum sanctum on PLpro-ISG15 De-Conjugation Machinery—A Computational Approach. Computation, 10(7), 109. https://doi.org/10.3390/computation10070109