
Fine-Tuning BirdNET for the Automatic Ecoacoustic Monitoring of Bird Species in the Italian Alpine Forests



Abstract

1. Introduction
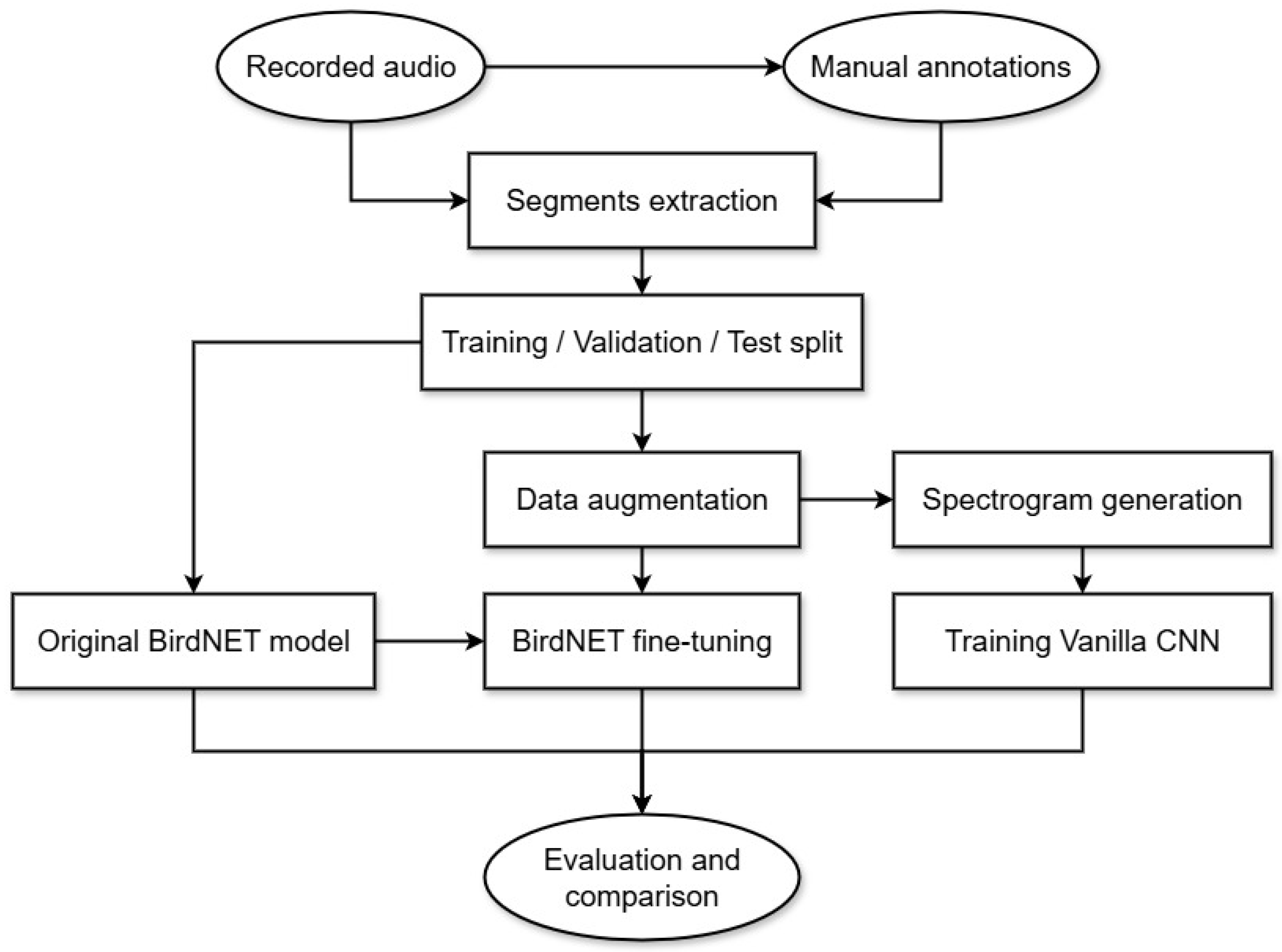
2. Materials and Methods
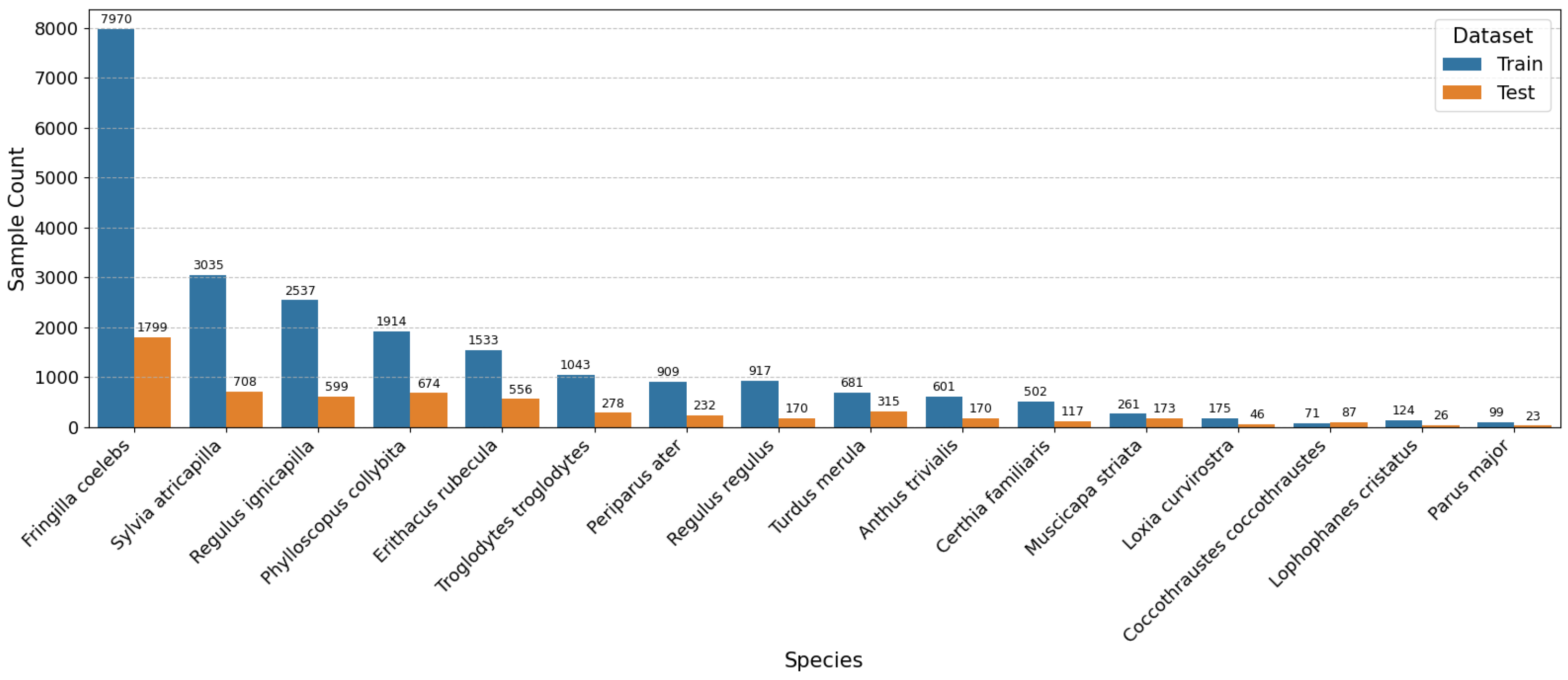
2.1. Database of Audio Recordings
2.2. Data Pre-Processing and Data Augmentation
2.3. Fine-Tuning the BirdNET Model
2.4. Baseline CNN Model
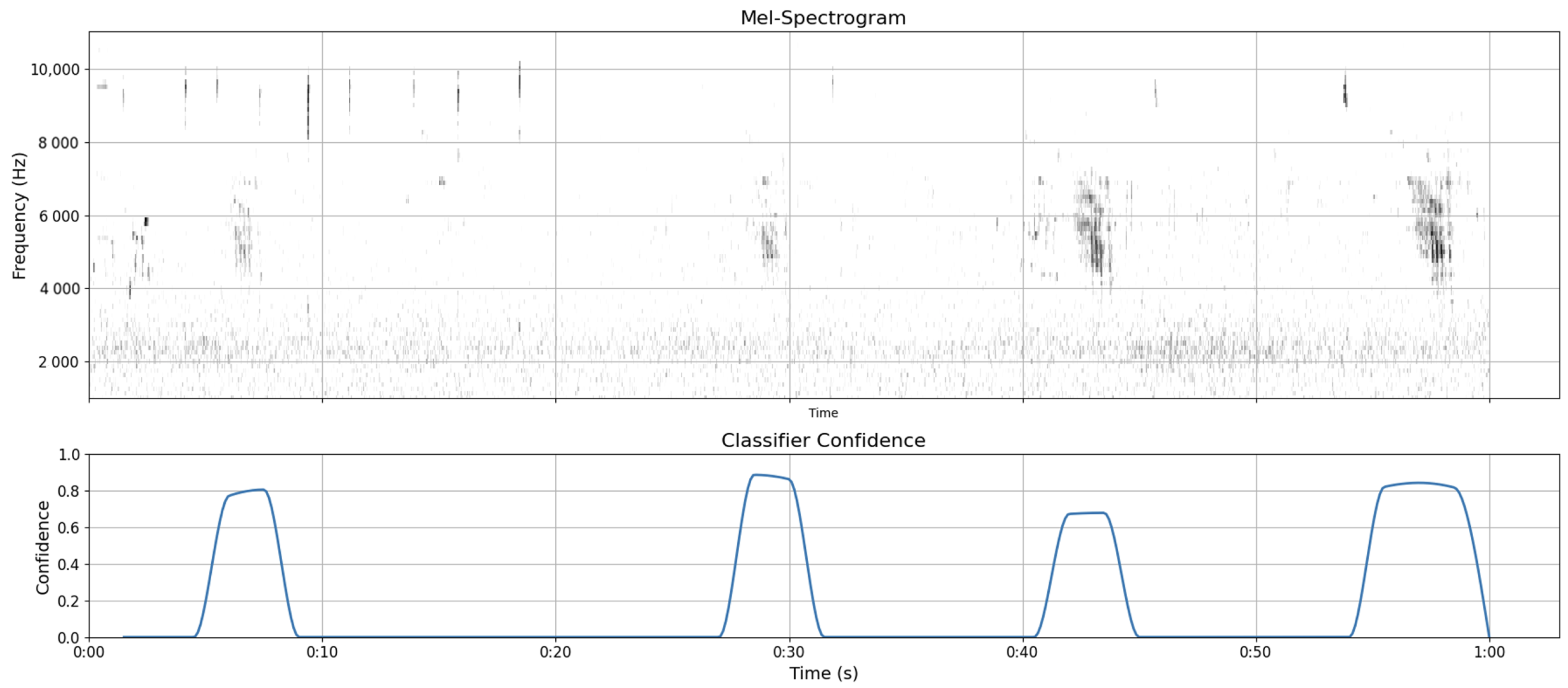
2.5. Performance Assessment
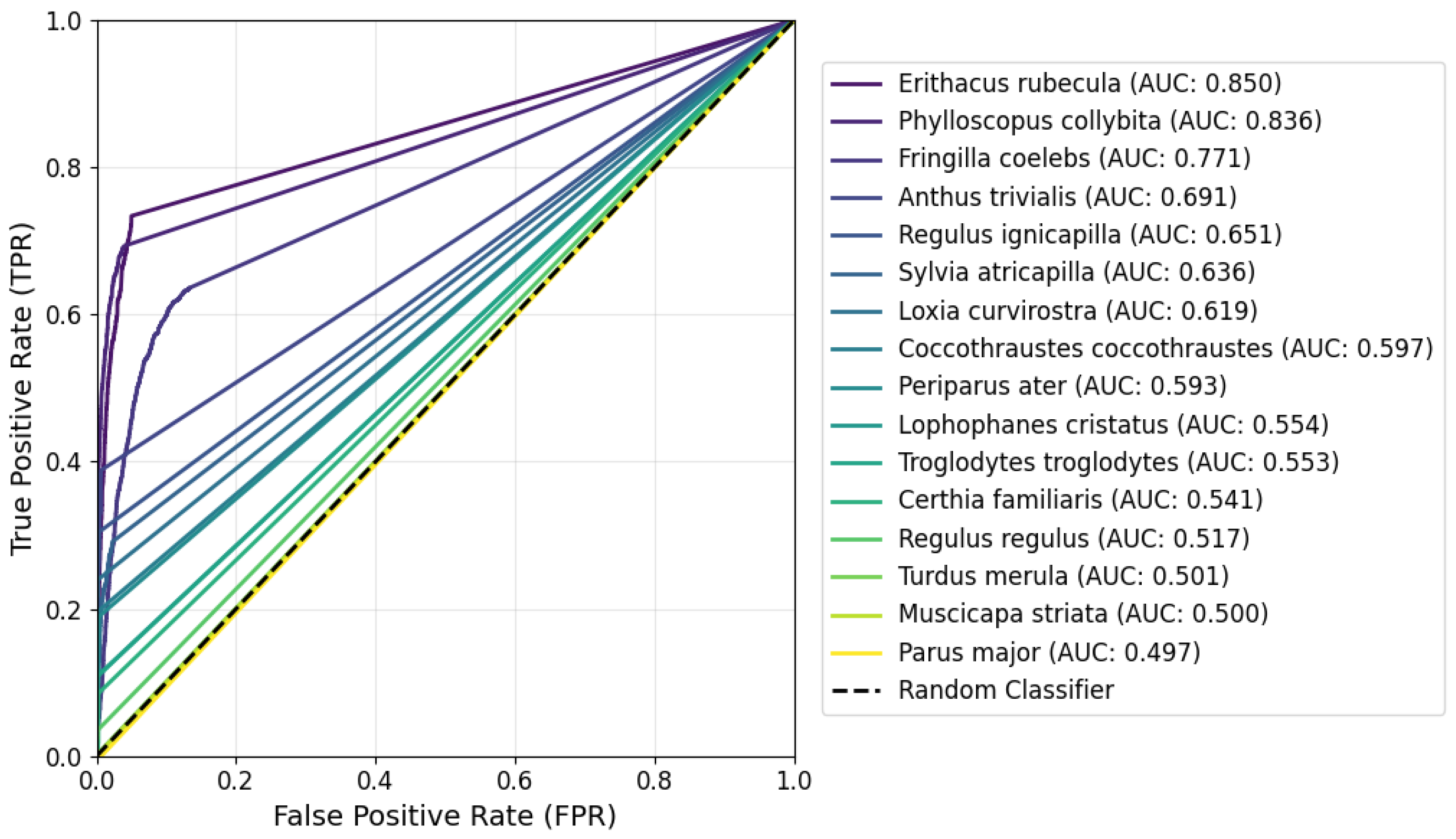
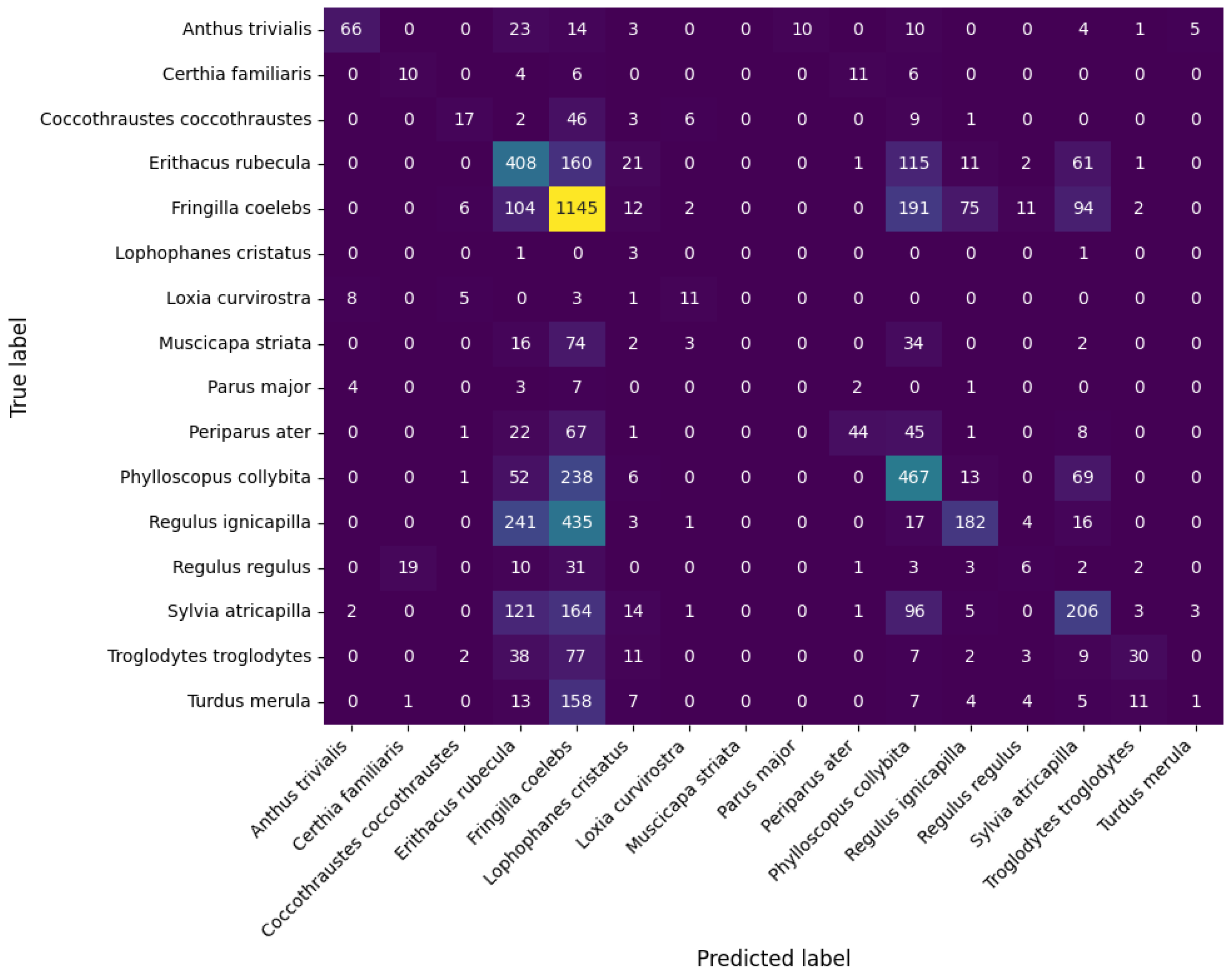
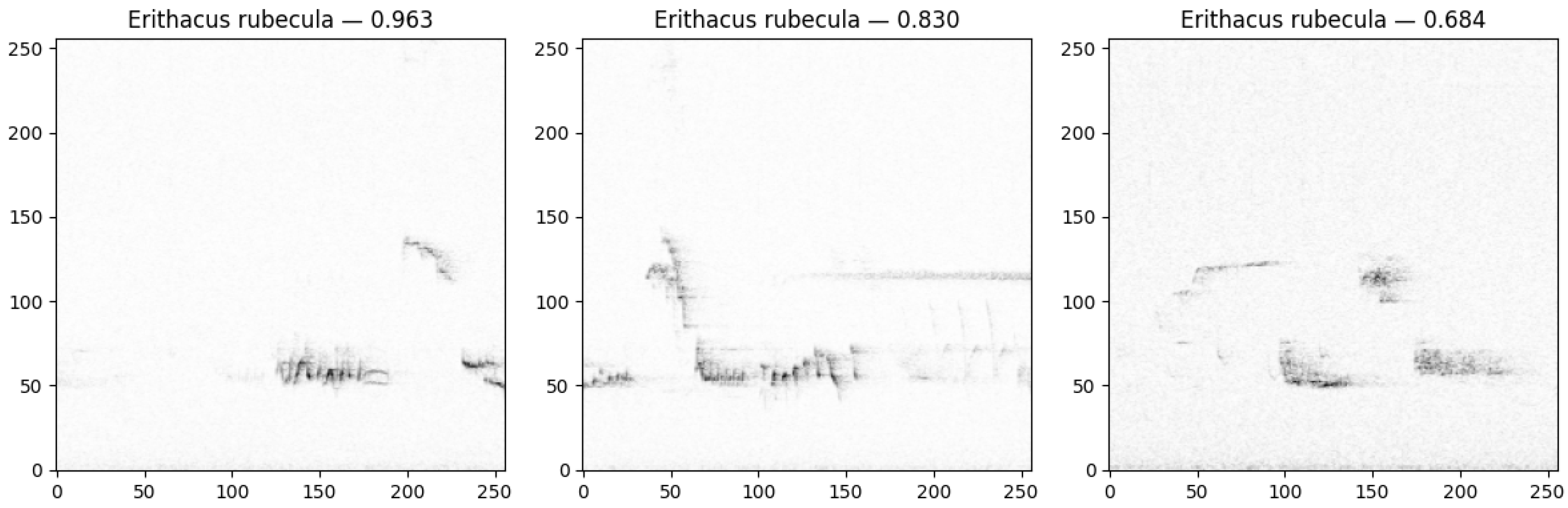
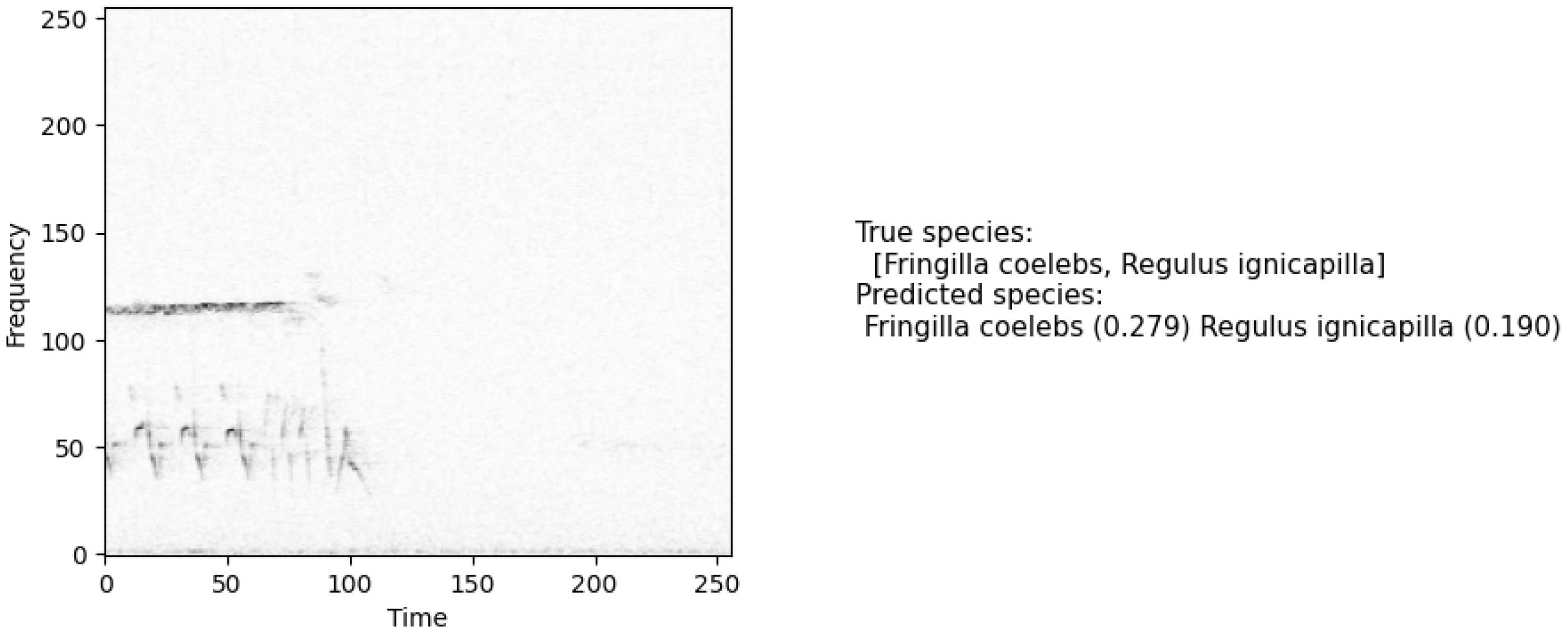
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butchart, S.H.; Walpole, M.; Collen, B.; Van Strien, A.; Scharlemann, J.P.; Almond, R.E.; Baillie, J.E.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- Scheffers, B.R.; Joppa, L.N.; Pimm, S.L.; Laurance, W.F. What we know and don’t know about Earth’s missing biodiversity. Trends Ecol. Evol. 2012, 27, 501–510. [Google Scholar] [CrossRef]
- Pocock, M.J.; Newson, S.E.; Henderson, I.G.; Peyton, J.; Sutherland, W.J.; Noble, D.G.; Ball, S.G.; Beckmann, B.C.; Biggs, J.; Brereton, T.; et al. Developing and enhancing biodiversity monitoring programmes: A collaborative assessment of priorities. J. Appl. Ecol. 2015, 52, 686–695. [Google Scholar] [CrossRef]
- Krause, B.; Farina, A. Using ecoacoustic methods to survey the impacts of climate change on biodiversity. Biol. Conserv. 2016, 195, 245–254. [Google Scholar] [CrossRef]
- Campedelli, T.; Florenzano, G.T.; Londi, G.; Cutini, S.; Fornasari, L. Effectiveness of the italian national protected areas system in conservation of farmland birds= Efectividad del sistema nacional de protección de áreas en Italia para la conservación de aves en paisajes agrícolas: A gap analysis= un análisis de deficiencias. Ardeola 2010, 57, 51–64. [Google Scholar]
- Portaccio, A.; Basile, M.; Favaretto, A.; Campagnaro, T.; Pettenella, D.; Sitzia, T. The role of Natura 2000 in relation to breeding birds decline on multiple land cover types and policy implications. J. Nat. Conserv. 2021, 62, 126023. [Google Scholar] [CrossRef]
- European Environment Agency. State of Nature in the EU—Results from Reporting Under the Nature Directives 2013–2018; Publications Office: Luxembourg, 2020. [Google Scholar]
- Segan, D.B.; Bottrill, M.C.; Baxter, P.W.; Possingham, H.P. Using conservation evidence to guide management. Conserv. Biol. 2011, 25, 200–202. [Google Scholar] [CrossRef]
- BirdLife International. Birds and Biodiversity Targets: What Do Birds Tell Us About Progress to the Aichi Targets and Requirements for the Post-2020 Biodiversity Framework? A State of the World’s Birds Report; BirdLife International: Cambridge, UK, 2020. [Google Scholar]
- Gregory, R.D.; Noble, D.; Field, R.; Marchant, J.; Raven, M.; Gibbons, D. Using birds as indicators of biodiversity. Ornis Hung. 2003, 12, 11–24. [Google Scholar]
- Darras, K.; Batáry, P.; Furnas, B.; Celis-Murillo, A.; Van Wilgenburg, S.L.; Mulyani, Y.A.; Tscharntke, T. Comparing the sampling performance of sound recorders versus point counts in bird surveys: A meta-analysis. J. Appl. Ecol. 2018, 55, 2575–2586. [Google Scholar] [CrossRef]
- Celis-Murillo, A.; Deppe, J.L.; Allen, M.F. Using soundscape recordings to estimate bird species abundance, richness, and composition. J. Field Ornithol. 2009, 80, 64–78. [Google Scholar] [CrossRef]
- Sugai, L.S.M.; Silva, T.S.F.; Ribeiro, J.W., Jr.; Llusia, D. Terrestrial passive acoustic monitoring: Review and perspectives. BioScience 2019, 69, 15–25. [Google Scholar] [CrossRef]
- Abrahams, C. Bird bioacoustic surveys-developing a standard protocol. Practice 2018, 102, 20–23. [Google Scholar]
- Acevedo, M.A.; Villanueva-Rivera, L.J. From the field: Using automated digital recording systems as effective tools for the monitoring of birds and amphibians. Wildl. Soc. Bull. 2006, 34, 211–214. [Google Scholar] [CrossRef]
- Priyadarshani, N.; Marsland, S.; Castro, I. Automated birdsong recognition in complex acoustic environments: A review. J. Avian Biol. 2018, 49, jav-01447. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, S.; Xu, Z.; Bellisario, K.; Dai, N.; Omrani, H.; Pijanowski, B.C. Automated bird acoustic event detection and robust species classification. Ecol. Inform. 2017, 39, 99–108. [Google Scholar] [CrossRef]
- Nieto-Mora, D.; Rodríguez-Buritica, S.; Rodríguez-Marín, P.; Martínez-Vargaz, J.; Isaza-Narváez, C. Systematic review of machine learning methods applied to ecoacoustics and soundscape monitoring. Heliyon 2023, 9, e20275. [Google Scholar] [CrossRef]
- Nur Korkmaz, B.; Diamant, R.; Danino, G.; Testolin, A. Automated detection of dolphin whistles with convolutional networks and transfer learning. Front. Artif. Intell. 2023, 6, 1099022. [Google Scholar] [CrossRef]
- Sprengel, E.; Jaggi, M.; Kilcher, Y.; Hofmann, T. Audio based bird species identification using deep learning techniques. In Proceedings of the Conference and Labs of the Evaluation Forum (CLEF) 2016, Evora, Portugal, 5–8 September 2016; pp. 547–559. [Google Scholar]
- Maegawa, Y.; Ushigome, Y.; Suzuki, M.; Taguchi, K.; Kobayashi, K.; Haga, C.; Matsui, T. A new survey method using convolutional neural networks for automatic classification of bird calls. Ecol. Inform. 2021, 61, 101164. [Google Scholar] [CrossRef]
- Madhusudhana, S.; Shiu, Y.; Klinck, H.; Fleishman, E.; Liu, X.; Nosal, E.M.; Helble, T.; Cholewiak, D.; Gillespie, D.; Širović, A.; et al. Improve automatic detection of animal call sequences with temporal context. J. R. Soc. Interface 2021, 18, 20210297. [Google Scholar] [CrossRef]
- Kahl, S.; Wood, C.M.; Eibl, M.; Klinck, H. BirdNET: A deep learning solution for avian diversity monitoring. Ecol. Inform. 2021, 61, 101236. [Google Scholar] [CrossRef]
- Wood, C.M.; Kahl, S.; Rahaman, A.; Klinck, H. The machine learning–powered BirdNET App reduces barriers to global bird research by enabling citizen science participation. PLoS Biol. 2022, 20, e3001670. [Google Scholar] [CrossRef]
- Pérez-Granados, C. BirdNET: Applications, performance, pitfalls and future opportunities. Ibis 2023, 165, 1068–1075. [Google Scholar] [CrossRef]
- Pérez-Granados, C. A first assessment of BirdNET performance at varying distances: A playback experiment. Ardeola 2023, 70, 257–269. [Google Scholar] [CrossRef]
- Matsinos, Y.G.; Tsaligopoulos, A. Hot spots of ecoacoustics in Greece and the issue of background noise. J. Ecoacoustics 2018, 2, 14. [Google Scholar] [CrossRef]
- Joppa, L.N.; Pfaff, A. High and far: Biases in the location of protected areas. PLoS ONE 2009, 4, e8273. [Google Scholar] [CrossRef]
- Haley, S.M.; Madhusudhana, S.; Branch, C.L. Comparing detection accuracy of mountain chickadee (Poecile gambeli) song by two deep-learning algorithms. Front. Bird Sci. 2024, 3, 1425463. [Google Scholar] [CrossRef]
- Hardersen, S.; Mason, F.; Viola, F.; Campedel, D.; Lasen, C.; Cassol, M. Research on the natural heritage of the reserves Vincheto di Celarda and Val Tovanella (Belluno province, Italy). In Conservation of Two Protected Areas in the Context of a LIFE Project. Quaderni Conservazione Habitat Invertebrati; Arti Grafiche Fiorini: Verona, Italy, 2008; Volume 5. [Google Scholar]
- Lasen, C.; Scariot, A.; Sitzia, T. Natura 2000 Habitats map, forest types and vegetation outline of Val Tovanella Nature Reserve. In Research on the Natural Heritage of the Reserves Vincheto di Celarda and Val Tovanella (Belluno Province Italy). Conservation of Two Protected Areas in the Context of a LIFE Project; Arti Grafiche Fiorini: Verona, Italy, 2008; pp. 325–334. [Google Scholar]
- Hill, A.P.; Prince, P.; Snaddon, J.L.; Doncaster, C.P.; Rogers, A. AudioMoth: A low-cost acoustic device for monitoring biodiversity and the environment. HardwareX 2019, 6, e00073. [Google Scholar] [CrossRef]
- Planqué, B.; Vellinga, W.P. Xeno-Canto: A 21st century way to appreciate neotropical bird song. Neotrop. Bird 2008, 3, 17–23. [Google Scholar]
- Vellinga, W.P.; Planqué, R. The Xeno-canto Collection and its Relation to Sound Recognition and Classification. In Proceedings of the CLEF (Working Notes), Toulouse, France, 8–11 September 2015. [Google Scholar]
- Krause, B. Bioacoustics, habitat ambience in ecological balance. Whole Earth Rev. 1987, 57, 14–18. [Google Scholar]
- Diamant, R.; Testolin, A.; Shachar, I.; Galili, O.; Scheinin, A. Observational study on the non-linear response of dolphins to the presence of vessels. Sci. Rep. 2024, 14, 6062. [Google Scholar] [CrossRef]
- Nanni, L.; Maguolo, G.; Paci, M. Data augmentation approaches for improving animal audio classification. Ecol. Inform. 2020, 57, 101084. [Google Scholar] [CrossRef]
- Stowell, D.; Wood, M.D.; Pamuła, H.; Stylianou, Y.; Glotin, H. Automatic acoustic detection of birds through deep learning: The first bird audio detection challenge. Methods Ecol. Evol. 2019, 10, 368–380. [Google Scholar] [CrossRef]
- Florentin, J.; Dutoit, T.; Verlinden, O. Detection and identification of European woodpeckers with deep convolutional neural networks. Ecol. Inform. 2020, 55, 101023. [Google Scholar] [CrossRef]
- Slabbekoorn, H.; Yeh, P.; Hunt, K. Sound transmission and song divergence: A comparison of urban and forest acoustics. Condor 2007, 109, 67–78. [Google Scholar] [CrossRef]
- Hoefer, S.; McKnight, D.T.; Allen-Ankins, S.; Nordberg, E.J.; Schwarzkopf, L. Passive acoustic monitoring in terrestrial vertebrates: A review. Bioacoustics 2023, 32, 506–531. [Google Scholar] [CrossRef]
- Pérez-Granados, C.; Funosas, D.; Morant, J.; Acosta, M.; Tarifa, R.; Devenish, D.K.N.; Fléau, H.L.; Rocha, P.A.; Bělka, T.; Desmet, L.; et al. Optimisation of passive acoustic bird surveys: A global assessment of BirdNET settings. Res. Sq. 2025. preprint (Version 1). [Google Scholar] [CrossRef]
- Staicer, C.A.; Spector, D.A.; Horn, A.G. The dawn chorus and other diel patterns in acoustic signaling. In Ecology and Evolution of Acoustic Communication in Birds; Comstock Pub. Associates: Ithaca, NY, USA, 1996; pp. 426–453. [Google Scholar]
- Luo, Y.; Mesgarani, N. Conv-TasNet: Surpassing ideal time-frequency magnitude masking for speech separation. IEEE/ACM Trans. Audio Speech Lang. Process. 2019, 27, 1256–1266. [Google Scholar] [CrossRef]
- Funosas, D.; Barbaro, L.; Schillé, L.; Elger, A.; Castagneyrol, B.; Cauchoix, M. Assessing the potential of BirdNET to infer European bird communities from large-scale ecoacoustic data. Ecol. Indic. 2024, 164, 112146. [Google Scholar] [CrossRef]
- Bota, G.; Manzano-Rubio, R.; Catalán, L.; Gómez-Catasús, J.; Pérez-Granados, C. Hearing to the unseen: AudioMoth and BirdNET as a cheap and easy method for monitoring cryptic bird species. Sensors 2023, 23, 7176. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Precision | Recall | F1-Score |
|---|---|---|---|
| Original BirdNET | 0.796 | 0.168 | 0.278 |
| Fine-tuned BirdNET | 0.590 | 0.666 | 0.626 |
| Fine-tuned BirdNET + data augmentation | 0.641 | 0.652 | 0.647 |
| Vanilla CNN + data augmentation | 0.445 | 0.435 | 0.440 |
| Model | Precision | Recall | F1-Score |
|---|---|---|---|
| Original BirdNET | 0.440 | 0.168 | 0.232 |
| Fine-tuned BirdNET | 0.612 | 0.666 | 0.601 |
| Fine-tuned BirdNET + data augmentation | 0.639 | 0.652 | 0.619 |
| Vanilla CNN + data augmentation | 0.600 | 0.435 | 0.477 |
| Model | Precision | Recall | F1-Score |
|---|---|---|---|
| Original BirdNET | 0.177 | 0.154 | 0.161 |
| Fine-tuned BirdNET | 0.621 | 0.673 | 0.632 |
| Fine-tuned BirdNET + data augmentation | 0.648 | 0.663 | 0.643 |
| Vanilla CNN + data augmentation | 0.379 | 0.443 | 0.393 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiavo, G.; Portaccio, A.; Testolin, A. Fine-Tuning BirdNET for the Automatic Ecoacoustic Monitoring of Bird Species in the Italian Alpine Forests. Information 2025, 16, 628. https://doi.org/10.3390/info16080628
Schiavo G, Portaccio A, Testolin A. Fine-Tuning BirdNET for the Automatic Ecoacoustic Monitoring of Bird Species in the Italian Alpine Forests. Information. 2025; 16(8):628. https://doi.org/10.3390/info16080628
Chicago/Turabian StyleSchiavo, Giacomo, Alessia Portaccio, and Alberto Testolin. 2025. "Fine-Tuning BirdNET for the Automatic Ecoacoustic Monitoring of Bird Species in the Italian Alpine Forests" Information 16, no. 8: 628. https://doi.org/10.3390/info16080628
APA StyleSchiavo, G., Portaccio, A., & Testolin, A. (2025). Fine-Tuning BirdNET for the Automatic Ecoacoustic Monitoring of Bird Species in the Italian Alpine Forests. Information, 16(8), 628. https://doi.org/10.3390/info16080628