Walking Ecosystems in Microbiome-Inspired Green Infrastructure: An Ecological Perspective on Enhancing Personal and Planetary Health


{kind=link}
{kind=link}
Abstract
1. Introduction—The Holobiont
2. Interconnectedness and Evolutionary Biology
3. Forty-Three Percent Human
4. Functional Ecology and Food for Thought
“An animal’s behaviour tends to maximize the survival of the genes ‘for’ that behaviour, whether or not those genes happen to be in the body of the particular animal performing it”.[38] (p. 13)
5. Dysbiosis—‘Life in Distress’
6. Microbiome Rewilding and Planetary Health
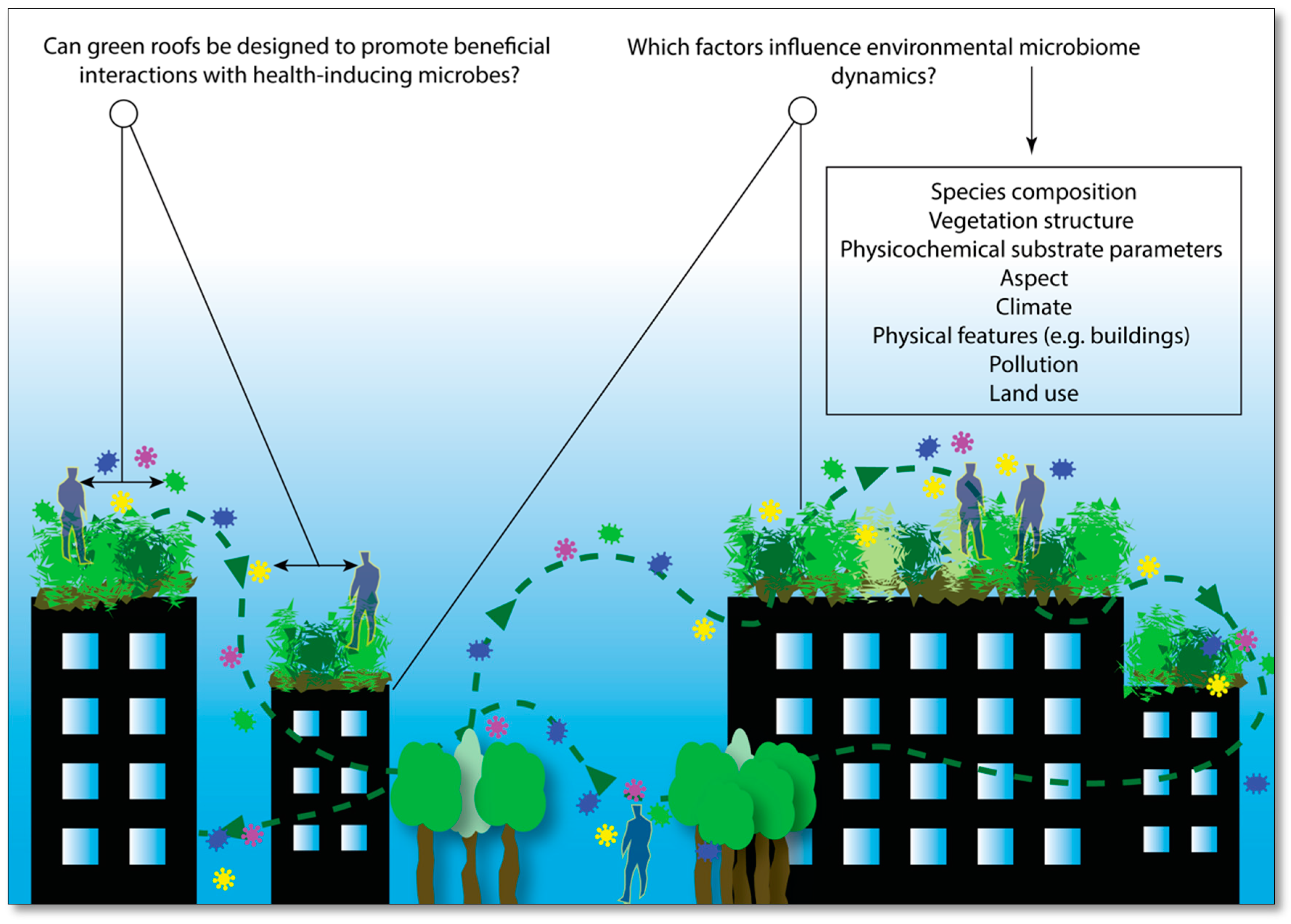
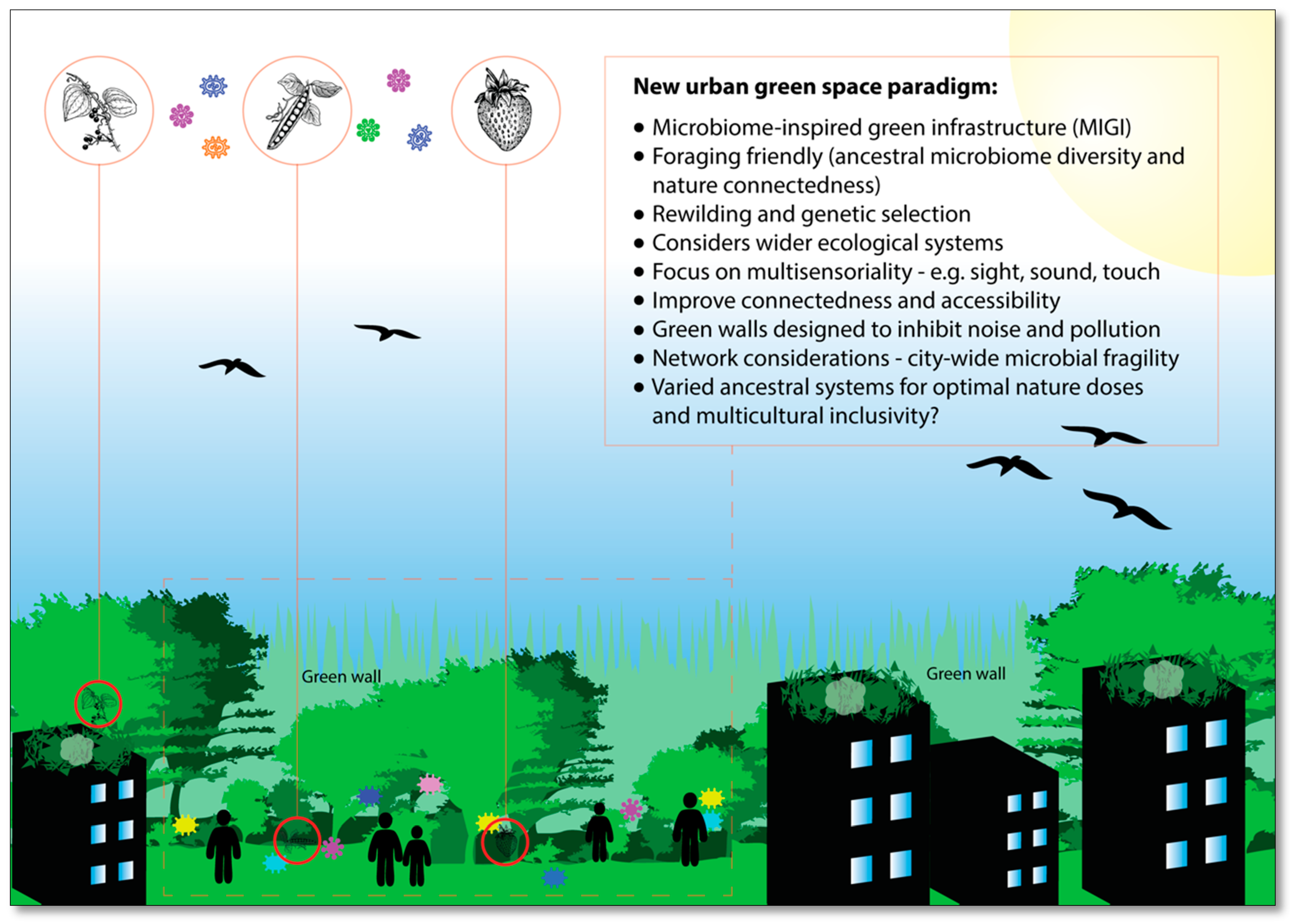
7. Microbiome-Inspired Green Infrastructure (MIGI)
7.1. Microbiome-Inspired Green Infrastructure—Foraging
7.2. Microbiome-Inspired Green Infrastructure—Green Barriers
7.3. Microbiome-Inspired Green Infrastructure—Cultural and Ancestral Microbiomes
7.4. Microbiome-Inspired Green Infrastructure—Plant Microbiome Selection and Engineering
8. Nature Connections and Green Prescriptions
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Van de Guchte, M.; Blottière, H.M.; Doré, J. Humans as holobionts: Implications for prevention and therapy. Microbiome 2018, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Dheilly, N.M. Holobiont–Holobiont interactions: Redefining host–parasite interactions. PLoS Pathog. 2014, 10, e1004093. [Google Scholar] [CrossRef] [PubMed]
- Pronin, E. How we see ourselves and how we see others. Science 2008, 320, 1177–1180. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.H.; Rubinstein, R.J.; Park, S.; Kelly, L.; Klepac-Ceraj, V. Microbiology and ecology are vitally important to premedical curricula. Evol. Med. Public Health 2015, 2015, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S.L.; Logan, A.C. Down to earth: Planetary health and biophilosophy in the symbiocene epoch. Challenges 2017, 8, 19. [Google Scholar] [CrossRef]
- Flandroy, L.; Poutahidis, T.; Berg, G.; Clarke, G.; Dao, M.C.; Decaestecker, E.; Furman, E.; Haahtela, T.; Massart, S.; Plovier, H.; et al. The impact of human activities and lifestyles on the interlinked microbiota and health of humans and of ecosystems. Sci. Total Environ. 2018, 627, 1018–1038. [Google Scholar] [CrossRef] [PubMed]
- Pongsiri, M.J.; Gatzweiler, F.W.; Bassi, A.M.; Haines, A.; Demassieux, F. The need for a systems approach to planetary health. Lancet Planet. Health 2017, 1, e257–e259. [Google Scholar] [CrossRef]
- Seltenrich, N. Down to Earth: The Emerging Field of Planetary Health. Environ. Health Perspect. 2018, 126, 072001. [Google Scholar] [CrossRef] [PubMed]
- Tonello, L.; Gashi, B.; Scuotto, A.; Capello, G.; Cocchi, M.; Gabrielli, F.; Tuszynski, J.A. The gastrointestinal-brain axis in humans as an evolutionary advance of the root-leaf axis in plants: A hypothesis linking quantum effects of light on serotonin and auxin. J. Integr. Neurosci. 2018, 17, 227–237. [Google Scholar] [CrossRef]
- Daubin, V.; Szöllősi, G.J. Horizontal gene transfer and the history of life. Cold Spring Harbor. Perspect. Biol. 2016, 8, a018036. [Google Scholar]
- Quammen, D. The Tangled Tree: A Radical New History of Life; Simon and Schuster, Inc.: New York, NY, USA, 2018. [Google Scholar]
- Sorriento, D.; Pascale, A.V.; Finelli, R.; Carillo, A.L.; Annunziata, R.; Trimarco, B.; Iaccarino, G. Targeting mitochondria as therapeutic strategy for metabolic disorders. Sci. World J. 2014, 2014, 604685. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.W. Mitochondrial evolution. Cold Spring Harbor. Perspect. Biol. 2012, 4, a011403. [Google Scholar]
- Knight, R. Integrated Approaches to Analyze Big Data in the Perinatal/Neonatal Space. Breastfeed. Med. 2018, 13, S-5–S-6. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, S.; Cui, H.; Chen, T. Reading the Underlying Information from Massive Metagenomic Sequencing Data. Proc. IEEE 2017, 105, 459–473. [Google Scholar] [CrossRef]
- Fierer, N.; Hamady, M.; Lauber, C.L.; Knight, R. The influence of sex, handedness.; washing on the diversity of hand surface bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 17994–17999. [Google Scholar] [CrossRef] [PubMed]
- Jordán, F.; Lauria, M.; Scotti, M.; Nguyen, T.P.; Praveen, P.; Morine, M.; Priami, C. Diversity of key players in the microbial ecosystems of the human body. Sci. Rep. 2015, 5, 15920. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, M.K.; Maurice, C.F. The Mammalian Gut as a Matchmaker. Cell Host Microbe 2017, 22, 726–727. [Google Scholar] [CrossRef] [PubMed]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Mazidi, M.; Rezaie, P.; Kengne, A.P.; Mobarhan, M.G.; Ferns, G.A. Gut microbiome and metabolic syndrome. Diabetes & Metabolic Syndrome. Clin. Res. Rev. 2016, 10, S150–S157. [Google Scholar]
- Sivaprakasam, S.; Prasad, P.D.; Singh, N. Benefits of short-chain fatty acids and their receptors in inflammation and carcinogenesis. Pharmacol. Ther. 2016, 164, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shimizu, Y.; Kimura, I. Gut microbial metabolite short-chain fatty acids and obesity. Biosci. Microbiota Food Health 2017, 36, 135–140. [Google Scholar] [CrossRef] [PubMed]
- TEDx. Feed Your Microbes—Nurture Your Mind; Online Video. 2017. Available online: https://www.youtube.com/watch?v=vKxomLM7SVc (accessed on 20 August 2018).
- Dobson, A.; Lodge, D.; Alder, J.; Cumming, G.S.; Keymer, J.; McGlade, J.; Mooney, H.; Rusak, J.A.; Sala, O.; Wolters, V.; et al. Habitat loss, trophic collapse, and the decline of ecosystem services. Ecology 2006, 87, 1915–1924. [Google Scholar] [CrossRef]
- Mahoney, P.C.; Bishop, M.J. Assessing risk of estuarine ecosystem collapse. Ocean Coast. Manag. 2017, 140, 46–58. [Google Scholar] [CrossRef]
- Ruokolainen, L.; von Hertzen, L.; Fyhrquist, N.; Laatikainen, T.; Lehtomäki, J.; Auvinen, P.; Karvonen, A.M.; Hyvärinen, A.; Tillmann, V.; Niemelä, O.; et al. Green areas around homes reduce atopic sensitisation in children. Allergy 2015, 70, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Clapp, M.; Aurora, N.; Herrera, L.; Bhatia, M.; Wilen, E.; Wakefield, S. Gut microbiota’s effect on mental health: The gut-brain axis. Clin. Pract. 2017, 7, 987. [Google Scholar] [CrossRef] [PubMed]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; FitzGerald, M.G.; Fulton, R.S.; et al. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar]
- Ferreyra, J.A.; Ng, K.M.; Sonnenburg, J.L. The enteric two-step: Nutritional strategies of bacterial pathogens within the gut. Cell. Microbiol. 2014, 16, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Guyot, V.; Castagneyrol, B.; Vialatte, A.; Deconchat, M.; Selvi, F.; Bussotti, F.; Jactel, H. Tree diversity limits the impact of an invasive forest pest. PLoS ONE 2015, 10, e0136469. [Google Scholar] [CrossRef] [PubMed]
- Alcock, J.; Maley, C.C.; Aktipis, C.A. Is eating behavior manipulated by the gastrointestinal microbiota? Evolutionary pressures and potential mechanisms. Bioessays 2014, 36, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota diurnal rhythmicity programs host transcriptome oscillations. Cell 2016, 167, 1495–1510. [Google Scholar] [CrossRef] [PubMed]
- Bagga, D.; Reichert, J.L.; Koschutnig, K.; Aigner, C.S.; Holzer, P.; Koskinen, K.; Moissl-Eichinger, C.; Schöpf, V. Probiotics drive gut microbiome triggering emotional brain signatures. Gut Microbes 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, P.; Spano, A.J.; Bowers, D.; Ren, T.; Moore, D.J.; Timko, M.P.; Wu, M.; Brayman, K.L. Evidence for the Role of the Cecal Microbiome in Maintenance of Immune Regulation and Homeostasis. Ann. Surg. 2018, 268, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar] [PubMed]
- Bonaz, B.; Bazin, T.; Pellissier, S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front. Neurosci. 2018, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, R. The Extended Phenotype; Oxford University Press: Oxford, UK, 1989; p. 13. ISBN 0-19-288051-9. [Google Scholar]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef] [PubMed]
- Ohri, N.; Gerich, M.E.; Fennimore, B.P.; Ir, D.; Robertson, C.E.; Regener, E.; Caplan, L.; Wagner, B.D.; Frank, D.N.; Kuhn, K. Bacterial Dysbiosis Associates with Functional Intraepithelial Lymphocyte Changes in Inflammatory Bowel Disease and Spondyloarthritis. J. Immunol. 2017, 198, 46–55. [Google Scholar]
- Logan, A.C. Dysbiotic drift: Mental health, environmental grey space.; microbiota. J. Physiol. Anthropol. 2015, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Polz, M.F.; Mazel, F.; Albright, M.B.; Huber, J.A.; O’Connor, M.I.; Ackermann, M.; Hahn, A.S.; Srivastava, D.S.; Crowe, S.A.; et al. Function and functional redundancy in microbial systems. Nat. Ecol. Evol. 2018, 2, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Ma, W.; Li, F.Y.; Zhang, Q.; Niu, J.; Ding, Y.; Han, F.; Sun, X. Functional redundancy instead of species redundancy determines community stability in a typical steppe of Inner Mongolia. PLoS ONE 2015, 10, e0145605. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.A.; Brunet, L.R. Old friends for breakfast. Clin. Exp. Allergy 2005, 35, 841–842. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.A.W.; Brunet, L.R. Microbes, immunoregulation, and the gut. Gut 2005, 54, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Haahtela, T.; Holgate, S.; Pawankar, R.; Akdis, C.A.; Benjaponpitak, S.; Caraballo, L.; Demain, J.; Portnoy, J.; von Hertzen, L. The biodiversity hypothesis and allergic disease: World allergy organization position statement. World Allergy Organ. J. 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, S.F.; Rook, G.A.; Scott, E.A.; Shanahan, F.; Stanwell-Smith, R.; Turner, P. Time to abandon the hygiene hypothesis: New perspectives on allergic disease, the human microbiome, infectious disease prevention and the role of targeted hygiene. Perspect. Public Health 2016, 136, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Pearson, D.G.; Craig, T. The great outdoors? Exploring the mental health benefits of natural environments. Front. Psychol. 2014, 5, 1178. [Google Scholar] [CrossRef] [PubMed]
- Snell, T.L.; Lam, J.C.; Lau, W.W.Y.; Lee, I.; Maloney, E.M.; Mulholland, N.; Wilson, L.; Wynne, L.J. Contact with nature in childhood and adult depression. Child. Youth Environ. 2016, 26, 111–124. [Google Scholar] [CrossRef]
- Mills, J.G.; Weinstein, P.; Gellie, N.J.C.; Weyrich, L.S.; Lowe, A.J.; Breed, M.F. Urban habitat restoration provides a human health benefit through microbiome rewilding: The Microbiome Rewilding Hypothesis. Restor. Ecol. 2017, 25, 866–872. [Google Scholar] [CrossRef]
- Stein, M.M.; Hrusch, C.L.; Gozdz, J.; Igartua, C.; Pivniouk, V.; Murray, S.E. Innate immunity and asthma risk in Amish and Hutterite farm children. N. Engl. J. Med. 2016, 375, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Brindley, P.; Jorgensen, A.; Maheswaran, R. Domestic gardens and self-reported health: A national population study. Int. J. Health Geogr. 2018, 17, 31. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Del Rio, T.G.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Lemieux-Labonté, V.; Tromas, N.; Shapiro, B.J.; Lapointe, F.-J. Environment and host species shape the skin microbiome of captive neotropical bats. PeerJ 2016, 4, e2430. [Google Scholar] [CrossRef] [PubMed]
- Laforest-Lapointe, I.; Paquette, A.; Messier, C.; Kembel, S.W. Leaf bacterial diversity mediates plant diversity and ecosystem function relationships. Nature 2017, 546, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Lehtimäki, J.; Karkman, A.; Laatikainen, T.; Paalanen, L.; Von Hertzen, L.; Haahtela, T.; Hanski, I.; Ruokolainen, L. Patterns in the skin microbiota differ in children and teenagers between rural and urban environments. Sci. Rep. 2017, 7, 45651. [Google Scholar] [CrossRef] [PubMed]
- Goddard, M.A.; Dougill, A.J.; Benton, T.G. Scaling up from gardens: Biodiversity conservation in urban environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Jansson, Å. Reaching for a sustainable, resilient urban future using the lens of ecosystem services. Ecol. Econ. 2013, 86, 285–291. [Google Scholar] [CrossRef]
- Weinstein, N.; Balmford, A.; Dehaan, C.R.; Gladwell, V.; Bradbury, R.B.; Amano, T. Seeing community for the trees: The links among contact with natural environments, community cohesion.; crime. Bioscience 2015, 65, 1141–1153. [Google Scholar] [CrossRef]
- Bögeholz, S. Nature experience and its importance for environmental knowledge, values and action: Recent German empirical contributions. Environ. Educ. Res. 2006, 12, 65–84. [Google Scholar] [CrossRef]
- Robinson, J.M.; Jorgensen, A. Rekindling Old Friendships in New Landscapes: The Environment-Microbiome-Health Axis in the Realms of Landscape Research. People Nat. 2018. manuscript submitted. [Google Scholar]
- Kim, H.J.; Kim, H.; Kim, J.J.; Myeong, N.R.; Kim, T.; Park, T.; Kim, E.; Choi, J.Y.; Lee, J.; An, S.; et al. Fragile skin microbiomes in megacities are assembled by a predominantly niche-based process. Sci. Adv. 2018, 4, e1701581. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.A. Regulation of the immune system by biodiversity from the natural environment: An ecosystem service essential to health. Proc. Natl. Acad. Sci. USA 2013, 110, 18360–18367. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, ethnicity or subsistence-specific variations in human microbiome composition and diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef] [PubMed]
- Nasidze, I.; Li, J.; Schroeder, R.; Creasey, J.L.; Li, M.; Stoneking, M. High diversity of the saliva microbiome in Batwa Pygmies. PLoS ONE 2011, 6, e23352. [Google Scholar] [CrossRef] [PubMed]
- Schnorr, S.L.; Candela, M.; Rampelli, S.; Centanni, M.; Consolandi, C.; Basaglia, G.; Turroni, S.; Biagi, E.; Peano, C.; Severgnini, M.; et al. Gut microbiome of the Hadza hunter-gatherers. Nat. Commun. 2014, 5, 3654. [Google Scholar] [CrossRef] [PubMed]
- Fragiadakis, G.K.; Smits, S.A.; Sonnenburg, E.D.; Van Treuren, W.; Reid, G.; Knight, R.; Manjurano, A.; Changalucha, J.; Dominguez-Bello, M.G.; Leach, J.; et al. Links between environment, diet, and the hunter-gatherer microbiome. bioRxiv 2018, 319673. [Google Scholar] [CrossRef] [PubMed]
- Franco, L.S.; Shanahan, D.F.; Fuller, R.A. A review of the benefits of nature experiences: More than meets the eye. Int. J. Environ. Res. Public Health 2017, 14, 864. [Google Scholar] [CrossRef] [PubMed]
- Van den Bosch, M.; Bird, W. Oxford Textbook of Nature and Public Health; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Shackleton, C.M.; Hurley, P.T.; Dahlberg, A.C.; Emery, M.R.; Nagendra, H. Urban foraging: A ubiquitous human practice overlooked by urban planners, policy, and research. Sustainability 2017, 9, 1884. [Google Scholar] [CrossRef]
- Kowalski, J.M.; Conway, T.M. Branching out: The inclusion of urban food trees in Canadian urban forest management plans. Urban For. Urban Green. 2018, in press. [Google Scholar] [CrossRef]
- Grow Sheffield. ‘The Abundance Project’. 2018. Available online: http://growsheffield.com/abundance/ (accessed on 2 November 2018).
- Avant Gardening. Mobile Allotment. 2018. Available online: http://www.avantgardening.org/projects/mobile.allotment (accessed on 2 November 2018).
- Jang, H.S.; Gim, G.M.; Kim, K.J.; Jung, H.H. Psychological Healing Change of Workers Following After the Experiencing Indoor Garden Installation. J. Hum. Ecol. 2017, 20, 373–383. [Google Scholar] [CrossRef]
- Veen, E.J.; Bock, B.B.; Van den Berg, W.; Visser, A.J.; Wiskerke, J.S. Community gardening and social cohesion: Different designs, different motivations. Local Environ. 2016, 21, 1271–1287. [Google Scholar] [CrossRef]
- Lowry, C.A.; Hollis, J.H.; De Vries, A.; Pan, B.; Brunet, L.R.; Hunt, J.R.; Paton, J.F.; van Kampen, E.; Knight, D.M.; Evans, A.K.; et al. Identification of an immune-responsive mesolimbocortical serotonergic system: Potential role in regulation of emotional behavior. Neuroscience 2007, 146, 756–772. [Google Scholar] [CrossRef] [PubMed]
- Fonken, L.K.; Frank, M.G.; D’Angelo, H.M.; Heinze, J.D.; Watkins, L.R.; Lowry, C.A.; Maier, S.F. Mycobacterium vaccae immunization protects aged rats from surgery-elicited neuroinflammation and cognitive dysfunction. Neurobiol. Aging 2018, 71, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Chen, L.; Liu, L.; Li, H.; Liu, B.; Zheng, D.; Liu, T.; Dong, J.; Sun, L.; Zhu, Y.; et al. Proteogenomic analysis and discovery of immune antigens in Mycobacterium vaccae. Mol. Cell. Proteom. 2017, 16, 1578–1590. [Google Scholar] [CrossRef] [PubMed]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Van Renterghem, T.; Hornikx, M.; Forssen, J.; Botteldooren, D. The potential of building envelope greening to achieve quietness. Build. Environ. 2013, 61, 34–44. [Google Scholar] [CrossRef]
- Abhijith, K.V.; Kumar, P.; Gallagher, J.; McNabola, A.; Baldauf, R.; Pilla, F.; Broderick, B.; Di Sabatino, S.; Pulvirenti, B. Air pollution abatement performances of green infrastructure in open road and built-up street canyon environments—A review. Atmos. Environ. 2017, 162, 71–86. [Google Scholar] [CrossRef]
- Soyiri, I.N.; Alcock, I. Green spaces could reduce asthma admissions. Lancet Respir. Med. 2018, 6, e1. [Google Scholar] [CrossRef]
- Crul, M. Super-diversity vs. assimilation: How complex diversity in majority–minority cities challenges the assumptions of assimilation. J. Ethn. Migr. Stud. 2016, 42, 54–68. [Google Scholar] [CrossRef]
- Nédélec, Y.; Sanz, J.; Baharian, G.; Szpiech, Z.A.; Pacis, A.; Dumaine, A.; Grenier, J.C.; Freiman, A.; Sams, A.J.; Hebert, S.; et al. Genetic ancestry and natural selection drive population differences in immune responses to pathogens. Cell 2016, 167, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Kwok, L.Y.; Zhang, J.; Guo, Z.; Gesudu, Q.; Zheng, Y.; Qiao, J.; Huo, D.; Zhang, H. Characterization of fecal microbiota across seven Chinese ethnic groups by quantitative polymerase chain reaction. PLoS ONE 2014, 9, e93631. [Google Scholar] [CrossRef] [PubMed]
- Gomez, A.; Petrzelkova, K.J.; Burns, M.B.; Yeoman, C.J.; Amato, K.R.; Vlckova, K.; Modry, D.; Todd, A.; Robinson, C.A.J.; Remis, M.J.; et al. Gut microbiome of coexisting BaAka Pygmies and Bantu reflects gradients of traditional subsistence patterns. Cell Rep. 2016, 14, 2142–2153. [Google Scholar] [CrossRef] [PubMed]
- Deschasaux, M.; Bouter, K.E.; Prodan, A.; Levin, E.; Groen, A.K.; Herrema, H.; Tremaroli, V.; Bakker, G.J.; Attaye, I.; Pinto-Sietsma, S.J.; et al. Depicting the composition of gut microbiota in a population with varied ethnic origins but shared geography. Nat. Med. 2018, 24, 1526–1531. [Google Scholar] [CrossRef] [PubMed]
- Gaulke, C.A.; Sharpton, T.J. The influence of ethnicity and geography on human gut microbiome composition. Nat. Med. 2018, 24, 1459–1496. [Google Scholar] [CrossRef] [PubMed]
- Soga, M.; Gaston, K.J.; Yamaura, Y.; Kurisu, K.; Hanaki, K. Both direct and vicarious experiences of nature affect children’s willingness to conserve biodiversity. Int. J. Environ. Res. Public Health 2016, 13, 529. [Google Scholar] [CrossRef] [PubMed]
- Hand, K.L.; Freeman, C.; Seddon, P.J.; Recio, M.R.; Stein, A.; van Heezik, Y. The importance of urban gardens in supporting children’s biophilia. Proc. Natl. Acad. Sci. USA 2017, 114, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Köberl, M.; Rybakova, D.; Müller, H.; Grosch, R.; Smalla, K. Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.D.; Ng, S.L.; Schwanen, T.; Zacharias, J.; Zhou, M.; Kawachi, I.; Sun, G. Pokémon GO and Physical Activity in Asia: Multilevel Study. J. Med. Internet Res. 2018, 20, e217. [Google Scholar] [CrossRef] [PubMed]
- Mallon, C.A.; van Elsas, J.D.; Salles, J.F. Microbial invasions: The process, patterns.; mechanisms. Trends Microbiol. 2015, 23, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Urbina, H.; Breed, M.F.; Zhao, W.; Gurrala, K.L.; Andersson, S.G.; Ågren, J.; Baldauf, S.; Rosling, A. Specificity in Arabidopsis thaliana recruitment of root fungal communities from soil and rhizosphere. Fungal Biol. 2018, 122, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Latz, E.; Eisenhauer, N.; Rall, B.C.; Scheu, S.; Jousset, A. Unravelling linkages between plant community composition and the pathogen-suppressive potential of soils. Sci. Rep. 2016, 6, 23584. [Google Scholar] [CrossRef] [PubMed]
- Mueller, U.G.; Sachs, J.L. Engineering microbiomes to improve plant and animal health. Trends Microbiol. 2015, 23, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Wallenstein, M.D. Managing and manipulating the rhizosphere microbiome for plant health: A systems approach. Rhizosphere 2017, 3, 230–232. [Google Scholar] [CrossRef]
- Hussain, S.S.; Mehnaz, S.; Siddique, K.H. Harnessing the Plant Microbiome for Improved Abiotic Stress Tolerance. In Plant Microbiome: Stress Response; Springer: Singapore, 2018; pp. 21–43. [Google Scholar]
- Van Heezik, Y.; Brymer, E. Nature as a commodity: What’s good for human health might not be good for ecosystem health. Front. Psychol. 2018, 9, 1673. [Google Scholar] [CrossRef] [PubMed]
- Capaldi, C.A.; Dopko, R.L.; Zelenski, J.M. The relationship between nature connectedness and happiness: A meta-analysis. Front. Psychol. 2014, 5, 976. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.S.; Frantz, C.M.; Bruehlman-Senecal, E.; Dolliver, K. Why is nature beneficial? The role of connectedness to nature. Environ. Behav. 2009, 41, 607–643. [Google Scholar] [CrossRef]
- Lumber, R.; Richardson, M.; Sheffield, D. Beyond knowing nature: Contact, emotion, compassion, meaning, and beauty are pathways to nature connection. PLoS ONE 2017, 12, e0177186. [Google Scholar] [CrossRef] [PubMed]
- Bragg, R.; Atkins, G. A Review of Nature-Based Interventions for Mental Health Care (NECR204). Available online: http://publications.naturalengland.org.uk/publication/4513819616346112 (accessed on 20 September 2018).
- Van den Berg, A.E. From green space to green prescriptions: Challenges and opportunities for research and practice. Front. Psychol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lovell, R.; Depledge, M.; Maxwell, S. Health and the Natural Environment: A Review of Evidence, Policy, Practice and Opportunities for the Future. 2018. Available online: https://beyondgreenspace.net/2018/09/07/defra_health_review/ (accessed on 21 September 2018).
- Prescott, S.; Logan, A.C. The Secret Life of Your Microbiome: Why Nature and Biodiversity Are Essential to Health and Happiness; New Society Publishers: Gabriola Island, BC, Canada, 2017. [Google Scholar]
- Albrecht, G. Ecopsychology in the Symbiocene. Ecopsychology 2014, 6, 58–59. [Google Scholar] [CrossRef]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinson, J.M.; Mills, J.G.; Breed, M.F. Walking Ecosystems in Microbiome-Inspired Green Infrastructure: An Ecological Perspective on Enhancing Personal and Planetary Health. Challenges 2018, 9, 40. https://doi.org/10.3390/challe9020040
Robinson JM, Mills JG, Breed MF. Walking Ecosystems in Microbiome-Inspired Green Infrastructure: An Ecological Perspective on Enhancing Personal and Planetary Health. Challenges. 2018; 9(2):40. https://doi.org/10.3390/challe9020040
Chicago/Turabian StyleRobinson, Jake M., Jacob G. Mills, and Martin F. Breed. 2018. "Walking Ecosystems in Microbiome-Inspired Green Infrastructure: An Ecological Perspective on Enhancing Personal and Planetary Health" Challenges 9, no. 2: 40. https://doi.org/10.3390/challe9020040
APA StyleRobinson, J. M., Mills, J. G., & Breed, M. F. (2018). Walking Ecosystems in Microbiome-Inspired Green Infrastructure: An Ecological Perspective on Enhancing Personal and Planetary Health. Challenges, 9(2), 40. https://doi.org/10.3390/challe9020040